Effects of intercropping soybeans with different root architecture and sweet maize on crop growth and phosphorus uptake
-
摘要:目的
研究不同磷水平下不同根构型大豆与甜玉米间作对作物磷吸收与生长的影响,探究间作体系内根系形态、构型及根际土壤磷有效性等的关系。
方法田间试验于2022年8月在广东省广州市增城区华南农业大学教学试验基地进行。以2个不同根构型的大豆品种‘本地2号’(深根型)和‘粤春03-3’(浅根型)为材料,采用裂区设计,主区为施磷(+P:大豆40 kg·hm−2、甜玉米120 kg· hm−2)和不施磷(−P:大豆0 kg·hm−2、甜玉米0 kg·hm−2) 2种施磷水平,副区为甜玉米单作、‘本地2号’‖甜玉米、‘粤春03-3’‖甜玉米、‘本地2号’单作和‘粤春03-3’单作5种种植模式。测定甜玉米和大豆的产量、生物量、磷吸收量以及根系形态和构型的相关指标,计算间作系统的土地当量比和种间竞争力。
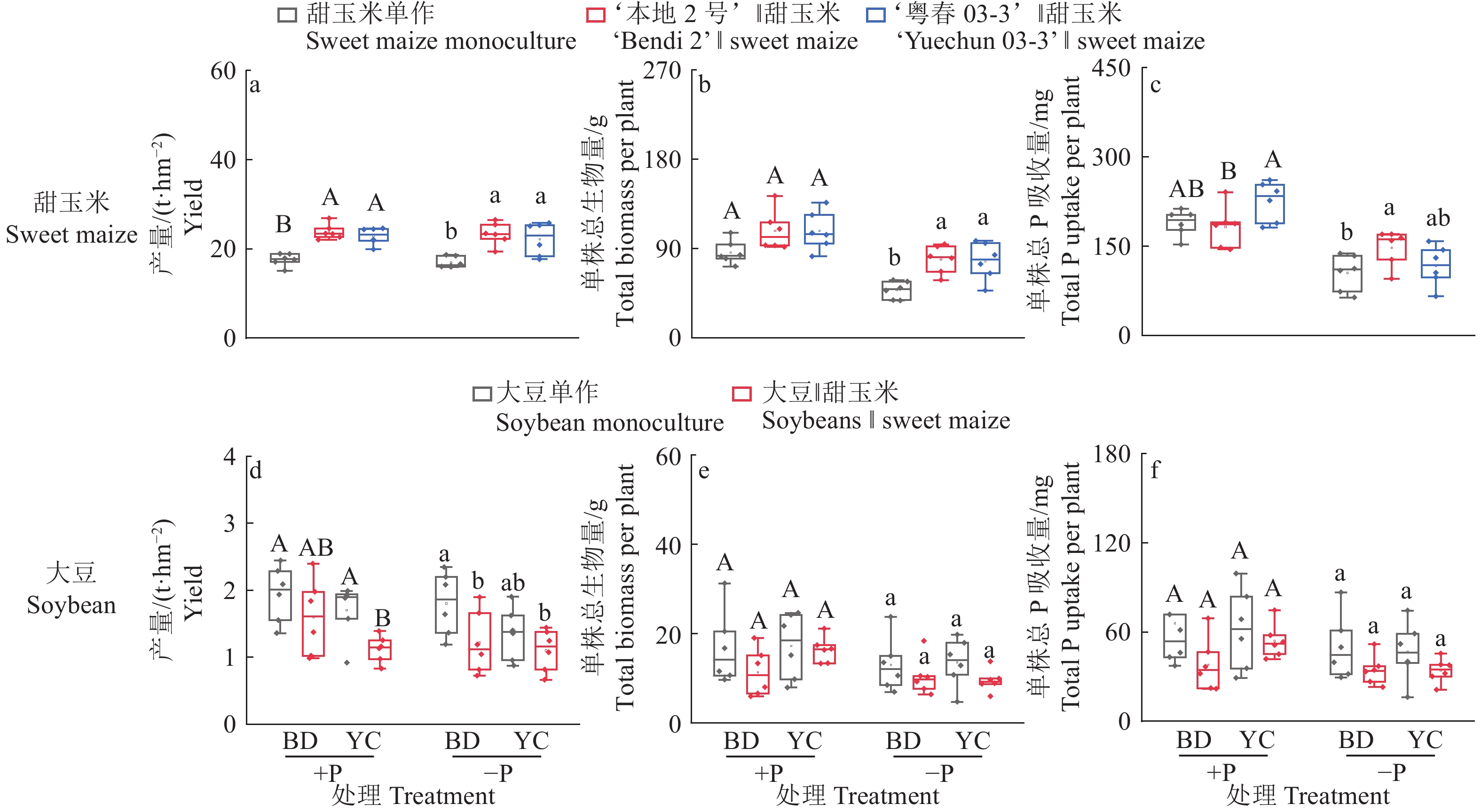
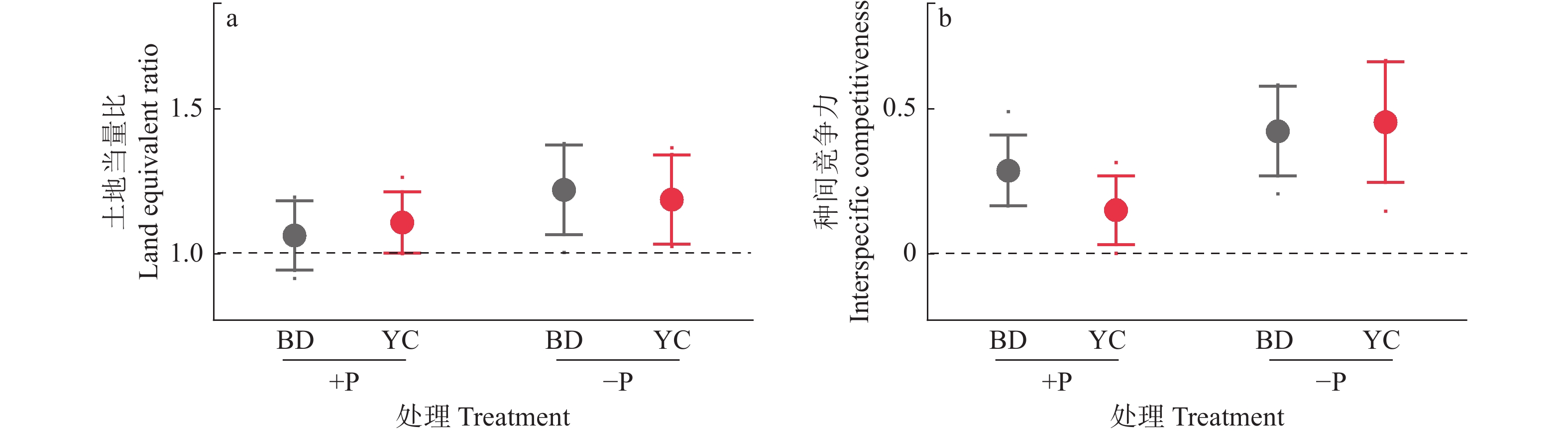
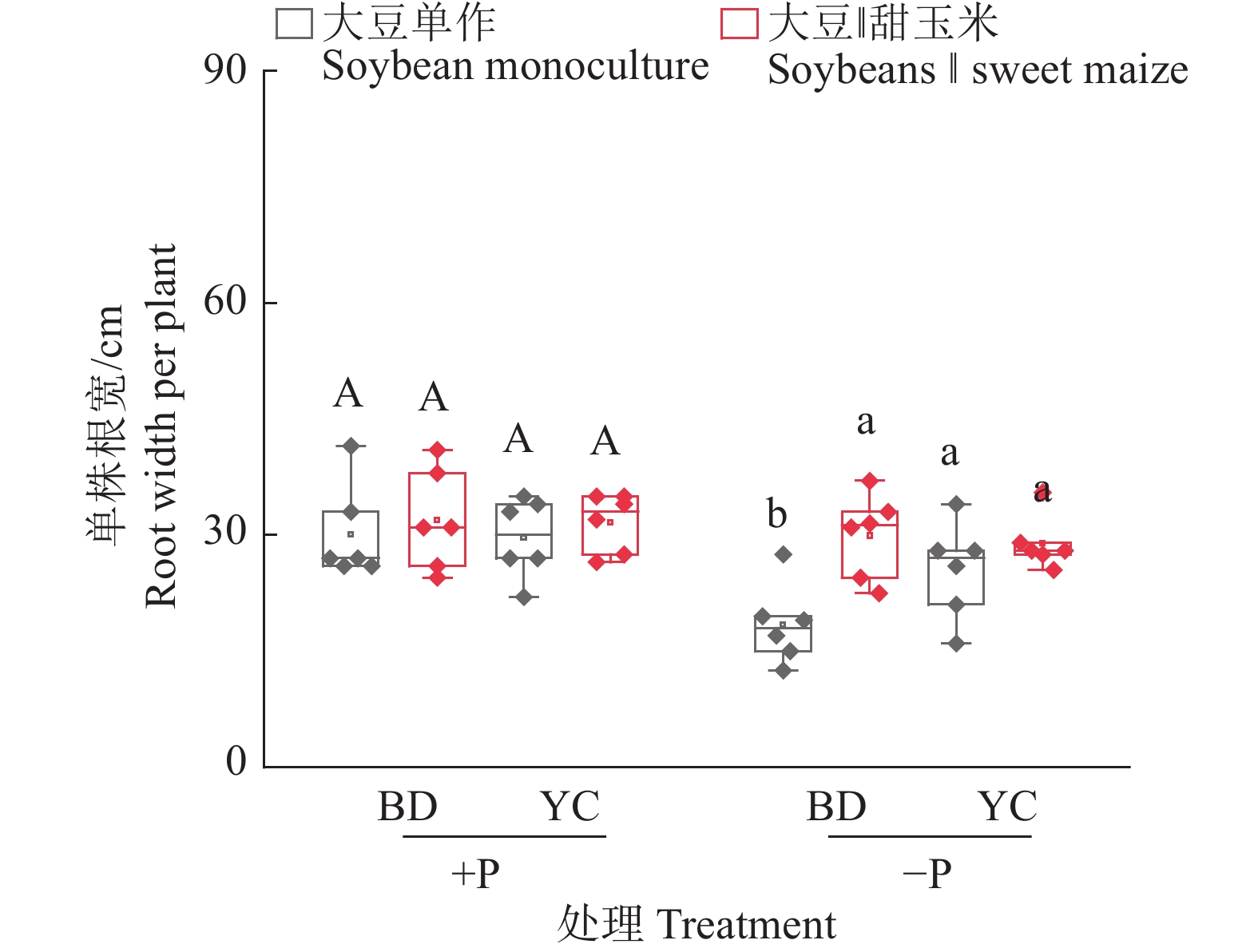
结果无论施磷还是不施磷条件下,间作大豆显著增加甜玉米产量,平均增产33.4%;不施磷条件下,间作大豆显著增加甜玉米生物量,平均增加62.7%。甜玉米和大豆间作具有间作优势,且受施磷水平影响;施磷条件下甜玉米与大豆间作的土地当量比为1.08,而不施磷条件下为1.21。种间竞争力分析表明,间作体系中,甜玉米竞争力显著强于大豆(种间竞争力>0),这种竞争优势在不施磷条件下更明显。此外,不施磷条件下间作深根型大豆‘本地2号’显著促进甜玉米磷吸收,平均增加40.6%。进一步分析发现,不施磷间作改变深根型大豆细根(直径≤0.5 mm)占比,同时诱导其根系拓宽,并显著提升甜玉米根际土壤有效磷含量。对间作体系内大豆和甜玉米性状的相关性分析发现,甜玉米的生物量与其总根长、大豆根宽和大豆根际土壤有效磷含量呈显著正相关;大豆的生物量与其磷吸收量、总根长、根宽以及根际土壤有效磷含量呈显著正相关,与其细根占比(直径≤0.5 mm)呈显著负相关。
结论不施磷条件下,间作深根型大豆可有效促进甜玉米生长与磷吸收,提高甜玉米产量。研究结果为充分挖掘甜玉米‖大豆间作系统磷素利用潜力、筛选适合间作的大豆品种、实现磷肥减施增效提供了重要理论依据。
Abstract:ObjectiveTo study the effects of intercropping soybeans with different root architecture and sweet maize on crop phosphorus (P) uptake and growth under different P levels, and explore the relationship among root morphology, root architecture, P availability in rhizospheric soil and so on within the intercropping system.
MethodField experiments were conducted in August 2022 at the Teaching and Experimental Base of South China Agricultural University in Zengcheng District, Guangzhou City, Guangdong Province. Two different soybean cultivars, namely ‘Bendi 2’ (deep-rooted genotype) and ‘Yuechun 03-3’ (shallow-rooted genotype) were selected as experimental materials. The field experiment employed a split-plot design, with the main plots comprising two P application levels: With P (+P: 40 kg·hm−2 for soybean, 120 kg·hm−2 for sweet maize) and without P (−P: 0 kg·hm−2 for both soybean and sweet maize). The subplots consisted of five different planting patterns as follows: Sweet maize monocropping, soybean ‘Bendi 2’ ‖ sweet maize, soybean ‘Yuechun 03-3’ ‖ sweet maize, soybean ‘Bendi 2’ monocropping and soybean ‘Yuechun 03-3’ monocropping. The yield, biomass, P uptake, root morphology and architecture of sweet maize and soybean were measured, and the land equivalent ratio (LER) and interspecific competitiveness of the intercropping system were calculated.
ResultIntercropping soybean significantly increased the yield of sweet maize by average of 33.4% under both P levels. Under −P condition, intercropping soybean significantly increased the biomass of sweet maize by average of 62.7%. Intercropping sweet maize and soybean exhibited advantages, influencing by P application levels. Under +P condition, the average LER for intercropped sweet maize and soybean was 1.08, while under −P condition, the average LER increased to 1.21. Interspecific competition analysis indicated that in the intercropping system, sweet maize showed significantly stronger competitiveness than soybean (interspecitic competitiveness > 0). This competitive advantage was more pronounced under −P condition. In addition, intercropping with deep-rooted soybean genotype (‘Bendi 2’) significantly promoted P uptake in sweet maize under −P condition by average of 40.6%. Further analysis revealed that for deep-rooted soybean ‘Bendi 2’, intercropping with sweet maize under −P condition widened its root width, altered the proportion of fine roots (diameter ≤ 0.5 mm), and significantly increased P availability in the rhizosphere of intercropping maize. The correlation analysis of traits of soybean and sweet maize in the intercropping system revealed a significant positive correlation between the biomass of sweet maize and its total root length, root width of soybean, and available P content in the soybean rhizosphere soil. The biomass of soybean showed a significant positive correlation with its P uptake, total root length, root width, and available P content in the rhizosphere soil. However, it exhibited a significant negative correlation with its proportion of fine roots (diameter ≤ 0.5 mm).
ConclusionIntercropping with deep rooting soybean under −P condition can effectively promote the growth and P absorption of sweet maize, and increase its yield. These findings provide important theoretical basis for fully exploring the P utilization potential in the sweet maize‖soybean intercropping system, screening suitable soybean varieties for crop intercropping, and achieving reduced P fertilization rate with increased P use efficiency.
-
Keywords:
- Phosphorus level /
- Intercropping /
- Soybean /
- Sweet maize /
- Root configuration /
- Phosphorus uptake
-
-
![]()
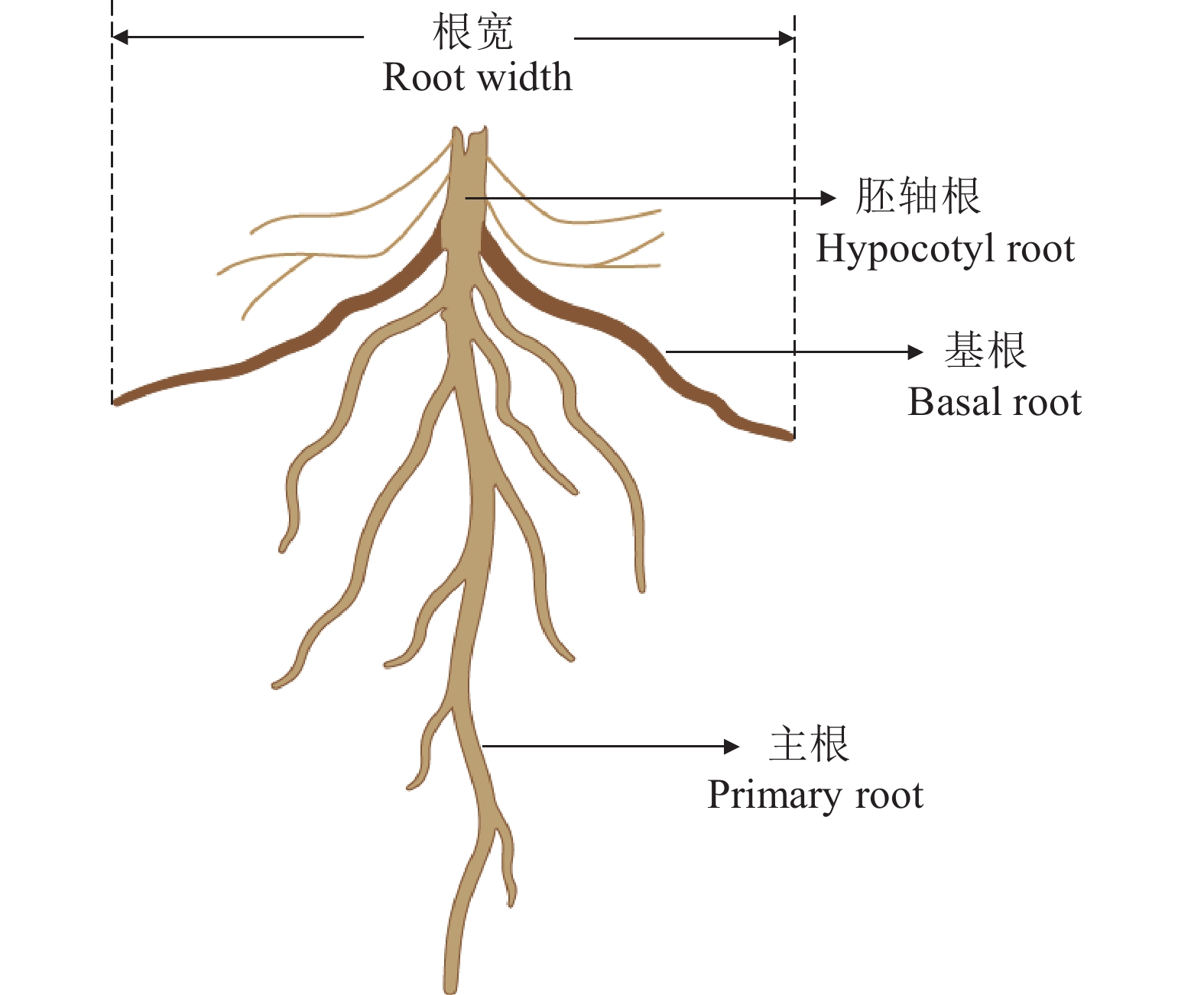
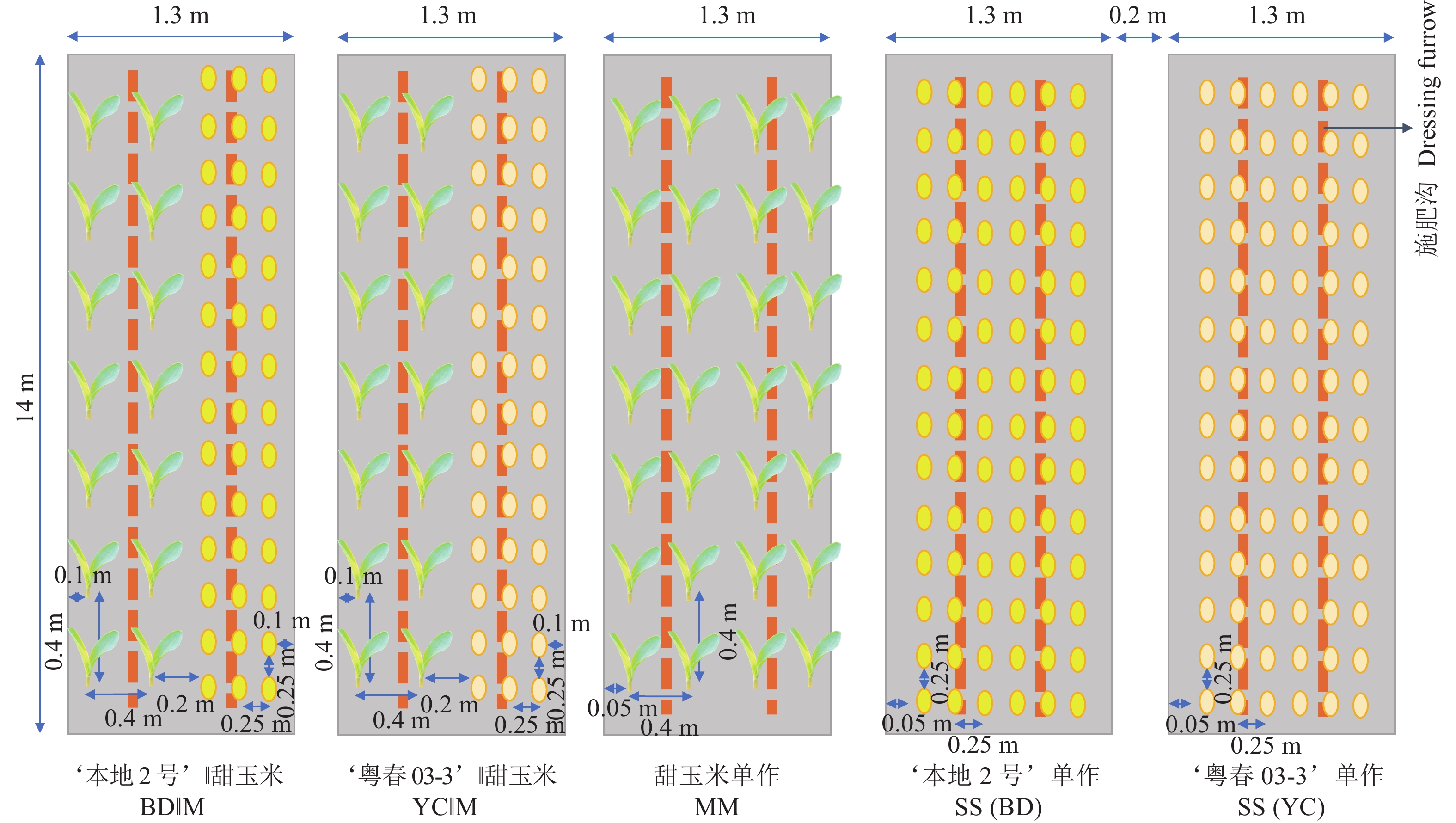
图 1 大豆、甜玉米间作和单作种植模式
Figure 1. Layouts of soybean, sweet maize intercropping system and monocultures
![]()
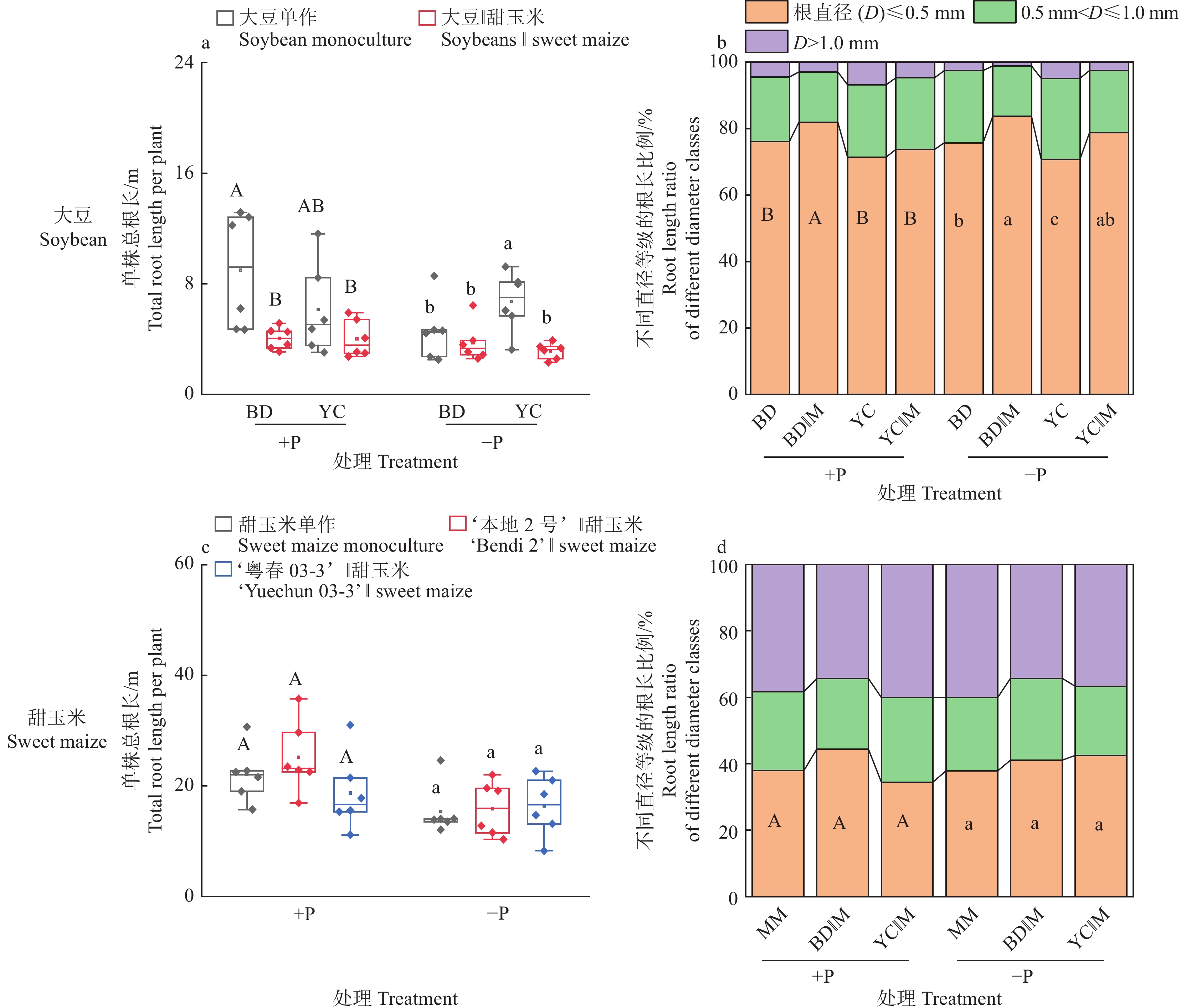
图 3 不同根构型大豆与甜玉米间作对作物产量、生物量及磷吸收量的影响
+P:施磷处理,−P:不施磷处理;BD:‘本地2号’,YC:‘粤春03-3’;箱体上不同大写和小写字母分别表示施磷和不施磷条件下不同种植模式之间差异显著(P<0.05,Duncan’s法)
Figure 3. Effects of intercropping of soybean with different root architecture and sweet maize on crop yield, biomass and phosphorus uptake
+P: Treatment with phosphate fertilizer, −P: Treatment without phosphate fertilizer; BD: ‘Bendi 2’, YC: ‘Yuechun 03-3’; Different capital and lowercase letters on the boxes indicate significant differences among different planting modes under +P and −P conditions respectively (P<0.05, Duncan’s method)
![]()
图 4 不同根构型大豆与甜玉米间作的间作优势(A)和种间竞争力(B)
+P:施磷处理,−P:不施磷处理;BD:‘本地2号’,YC:‘粤春03-3’
Figure 4. Intercropping advantages (A) and interspecific competitiveness (B) of soybean with different root architecture intercropping with sweet maize
+P: Treatment with phosphate fertilizer, −P: Treatment without phosphate fertilizer; BD: ‘Bendi 2’, YC: ‘Yuechun 03-3 ’
![]()
图 5 不同根构型大豆与甜玉米间作对大豆根宽的影响
+P:施磷处理,−P:不施磷处理;BD:‘本地2号’,YC:‘粤春03-3’;箱体上不同大写和小写字母分别表示施磷和不施磷条件下不同种植模式之间差异显著(P<0.05,Duncan’s法)
Figure 5. Effects of intercropping soybean with different root architecture and sweet maize on root width of soybeans
+P: Treatment with phosphate fertilizer, −P: Treatment without phosphate fertilizer; BD: ‘Bendi 2’, YC: ‘Yuechun 03-3’; Different capital and lowercase letters on the boxes indicate significant differences among different planting modes under +P and −P conditions respectively (P<0.05, Duncan’s method)
![]()
图 6 不同根构型大豆与甜玉米间作对植株根系形态的影响
+P:施磷处理,−P:不施磷处理;BD:‘本地2号’,YC:‘粤春03-3’;箱体上不同大写和小写字母分别表示施磷和不施磷条件下不同种植模式之间差异显著(P<0.05,Duncan’s法)
Figure 6. Effects of intercropping soybean with different root architecture and sweet maize on root morphology of plants
+P: Treatment with phosphate fertilizer, −P: Treatment without phosphate fertilizer; BD: ‘Bendi 2’, YC: ‘Yuechun 03-3’; Different capital and lowercase letters on the boxes indicate significant differences among different planting modes under +P and −P conditions respectively (P<0.05, Duncan’s method)
![]()
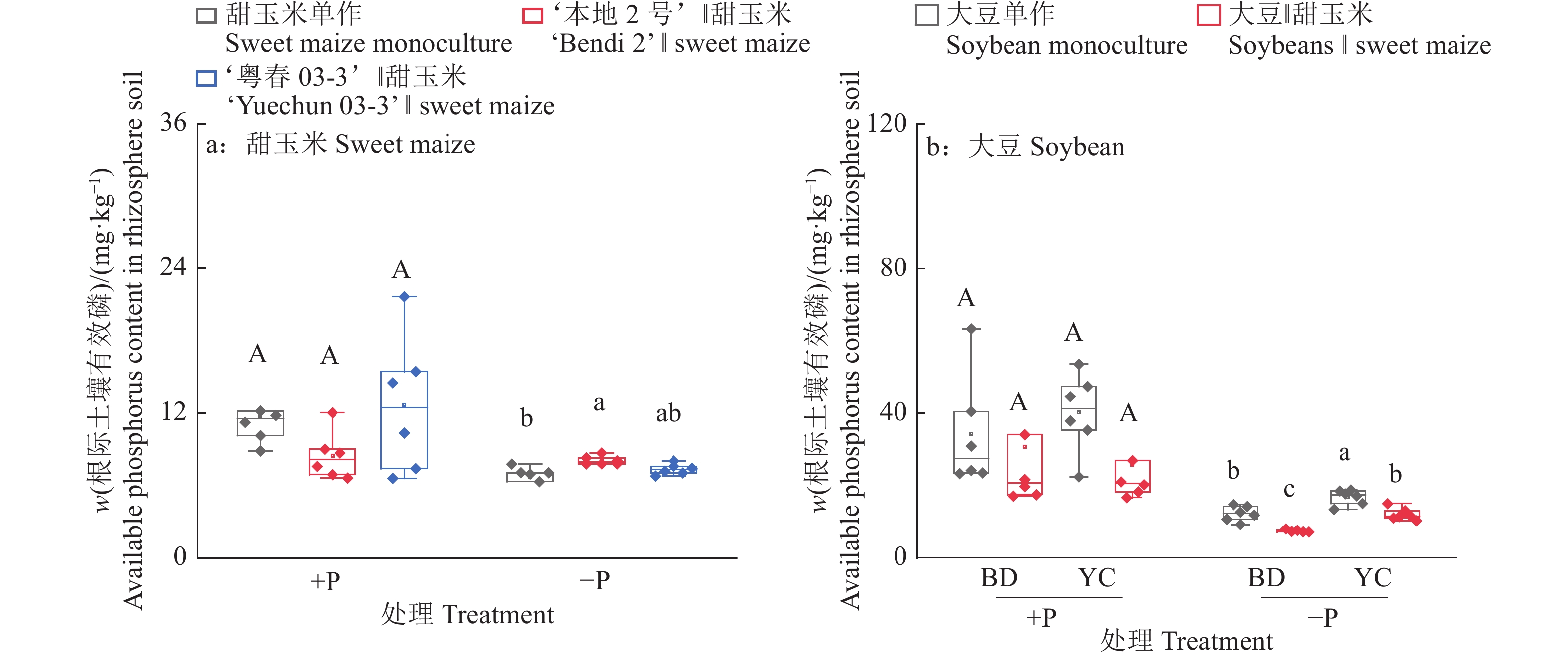
图 7 不同根构型大豆与甜玉米间作对植株根际土壤有效磷含量的影响
+P:施磷处理,−P:不施磷处理;BD:‘本地2号’,YC:‘粤春03-3’;箱体上不同大写和小写字母分别表示施磷和不施磷条件下不同种植模式之间差异显著(P<0.05,Duncan’s法)
Figure 7. Effects of intercropping soybean with different root architecture and sweet maize on available phosphorus content in rhizospheric soil of plants
+P: Treatment with phosphate fertilizer, −P: Treatment without phosphate fertilizer; BD: ‘Bendi 2’, YC: ‘Yuechun 03-3’; Different capital and lowercase letters on the boxes indicate significant differences among different planting modes under +P and −P conditions respectively (P<0.05, Duncan’s method)
![]()
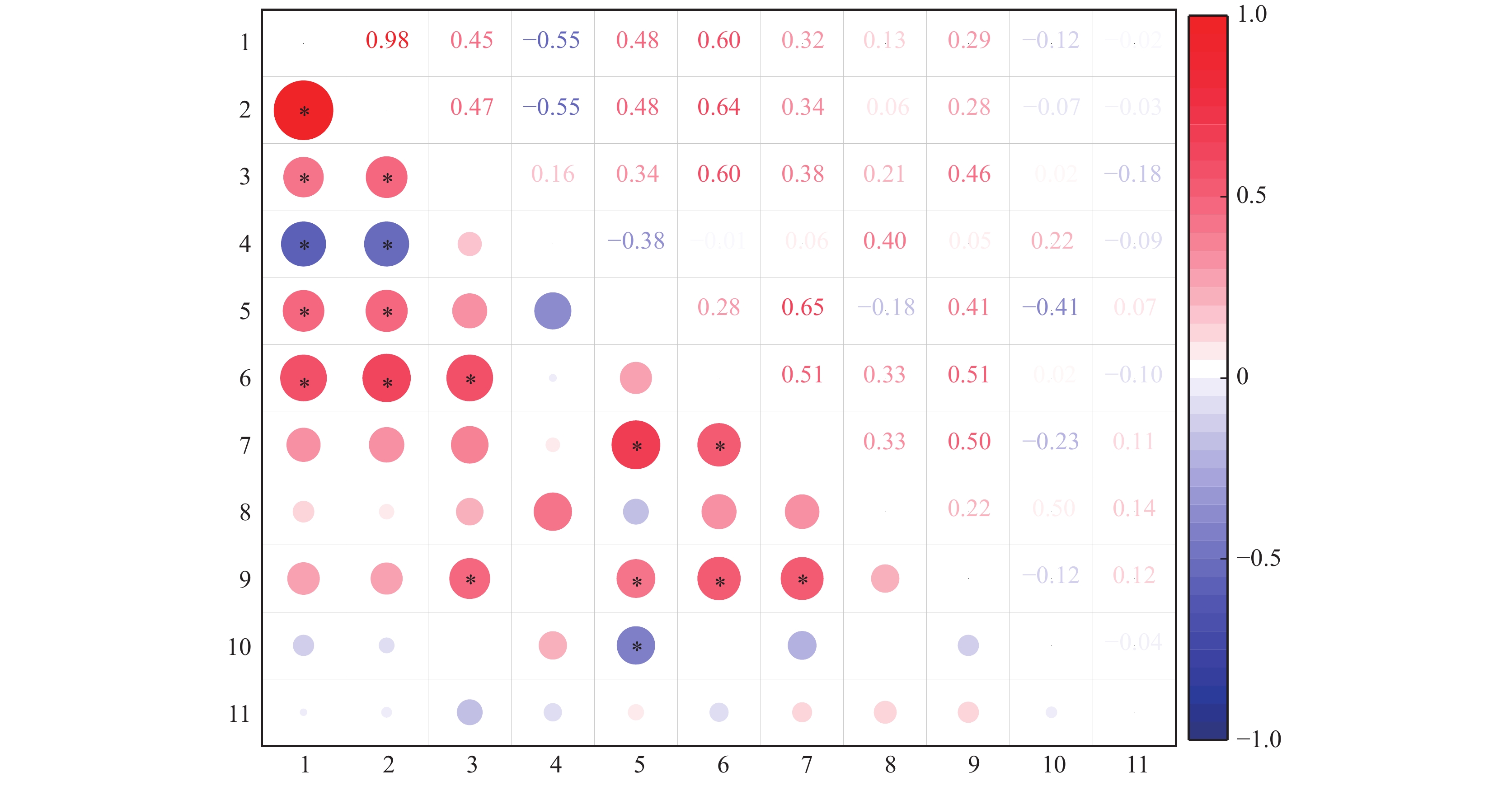
图 8 间作大豆和甜玉米表型参数间的相关性分析
1:大豆总生物量,2:大豆总磷吸收量,3:大豆总根长,4:大豆直径≤0.5 mm细根占比,5:大豆根际土壤有效磷含量,6:大豆根宽,7:甜玉米总生物量,8:甜玉米总磷吸收量,9:甜玉米总根长,10:甜玉米直径≤0.5 mm细根占比,11:甜玉米根际土壤有效磷含量;相关系数(R)在图中以不同颜色展示,右侧图例是不同R值的颜色区间,红色表示正相关,蓝色表示负相关;“*”表示在P<0.05水平显著相关(Spearman法)
Figure 8. Correlation analysis of phenotypic parameters in intercropped soybean and sweet maize
1: Total biomass of soybean, 2: Total phosphorus uptake of soybean, 3: Total root length of soybean, 4: Proportion of diameter ≤0.05 mm fine roots to total roots in soybean, 5: Available phosphorus content in soybean rhizospheric soil, 6: Root width of soybean, 7: Total biomass of sweet maize, 8: Total phosphorus uptake of sweet maize, 9: Total root length of sweet maize, 10: Proportion of diameter ≤0.05 mm fine roots to total roots in sweet maize, 11: Available phosphorus content in sweet maize rhizospheric soil; Colors in the heatmap indicate different correlation coefficients (R) and the color interval is shown in the right legend, red indicates positive correlation, blue indicates negative correlation; “*” indicates significant correlation at P<0.05 level (Spearman method)
-
[1] SHEN J, YUAN L, ZHANG J, et al. Phosphorus dynamics: From soil to plant[J]. Plant Physiology, 2011, 156(3): 997-1005. doi: 10.1104/pp.111.175232
[2] 刘瑾, 杨建军. 近三十年农田土壤磷分子形态的研究进展[J]. 土壤学报, 2021, 58(3): 558-567. [3] 赵学强, 潘贤章, 马海艺, 等. 中国酸性土壤利用的科学问题与策略[J]. 土壤学报, 2023, 60(5): 1248-1263. [4] REID J B, TAN Y, GRIFFIN W B, et al. Root system plasticity as a factor in crop breeding for improved nutrient uptake[C]//Agronomy New Zealand. 40th Agronomy Society Conference. Lincoln, 2010, 40: 51-64.
[5] KARLOVA R, BOER D, HAYES S, et al. Root plasticity under abiotic stress[J]. Plant Physiology, 2021, 187(3): 1057-1070. doi: 10.1093/plphys/kiab392
[6] HALING R E, BROWN L K, BENGOUGH A G, et al. Root hairs improve root penetration, root-soil contact, and phosphorus acquisition in soils of different strength[J]. Journal of Experimental Botany, 2013, 64(12): 3711-3721. doi: 10.1093/jxb/ert200
[7] SOUMYA P R, SHARMA S, MEENA M K, et al. Response of diverse bread wheat genotypes in terms of root architectural traits at seedling stage in response to low phosphorus stress[J]. Plant Physiology Reports, 2021, 26(1): 152-161. doi: 10.1007/s40502-020-00540-6
[8] HALING R E, YANG Z, SHADWELL N, et al. Root morphological traits that determine phosphorus-acquisition efficiency and critical external phosphorus requirement in pasture species[J]. Functional Plant Biology, 2016, 43(9): 815-826. doi: 10.1071/FP16037
[9] 刘鹏, 区伟贞, 王金祥, 等. 磷有效性与植物侧根的发生发育[J]. 植物生理学通讯, 2006(3): 395-400. [10] MIGUEL M A, POSTMA J A, LYNCH J P. Phene synergism between root hair length and basal root growth angle for phosphorus acquisition[J]. Plant Physiology, 2015, 167(4): 1430-1439. doi: 10.1104/pp.15.00145
[11] YI K, LI X, CHEN D, et al. Shallower root spatial distribution induced by phosphorus deficiency contributes to topsoil foraging and low phosphorus adaption in sugarcane (Saccharum officinarum L. )[J]. Frontiers in Plant Science, 2022, 12: 797635. doi: 10.3389/fpls.2021.797635
[12] XIA H, ZHAO J, SUN J, et al. Dynamics of root length and distribution and shoot biomass of maize as affected by intercropping with different companion crops and phosphorus application rates[J]. Field Crops Research, 2013, 150: 52-62. doi: 10.1016/j.fcr.2013.05.027
[13] LI L, LIU Y X, LI X F. Intercropping to maximize root-root interactions in agricultural plants: Agronomic aspects[M]//The root systems in sustainable agricultural intensification. 2021: 309-328. doi: 10.1002/9781119525417.ch12.
[14] 覃潇敏, 潘浩男, 肖靖秀, 等. 不同磷水平下玉米−大豆间作系统根系形态变化[J]. 应用生态学报, 2021, 32(9): 3223-3230. [15] AN R, YU R, XING Y, et al. Enhanced phosphorus-fertilizer-use efficiency and sustainable phosphorus management with intercropping[J]. Agronomy for Sustainable Development, 2023, 43(5): 57. doi: 10.1007/s13593-023-00916-6
[16] 雍太文, 刘小明, 宋春, 等. 种植方式对玉米−大豆套作体系中作物产量、养分吸收和种间竞争的影响[J]. 中国生态农业学报, 2015, 23(6): 659-667. [17] 唐劲驰, ISMAEL A M, 佘丽娜, 等. 大豆根构型在玉米/大豆间作系统中的营养作用[J]. 中国农业科学, 2005(6): 1196-1203. [18] 赵静, 付家兵, 廖红, 等. 大豆磷效率应用核心种质的根构型性状评价[J]. 科学通报, 2004, 49(13): 1249-1257. [19] 朱同林, 方素琴, 李志垣, 等. 基于图像重建的根系三维构型定量分析及其在大豆磷吸收研究中的应用[J]. 科学通报, 2006, 51(16): 1885-1893. [20] 郑林生, 刘志强, 陈康, 等. 基因型影响磷镁互作下大豆生长及根瘤和菌根性状[J]. 植物营养与肥料学报, 2022, 28(9): 1685-1698. [21] BURRIDGE J, JOCHUA C N, BUCKSCH A, et al. Legume shovelomics: High-throughput phenotyping of common bean (Phaseolus vulgaris L.) and cowpea (Vigna unguiculata subsp, unguiculata) root architecture in the field[J]. Field Crops Research, 2016, 192: 21-32.
[22] WALK T C, JARAMILLO R, LYNCH J P. Architectural tradeoffs between adventitious and basal roots for phosphorus acquisition[J]. Plant and Soil, 2006, 279(1/2): 347-366.
[23] AO J, FU J, TIAN J, et al. Genetic variability for root morph-architecture traits and root growth dynamics as related to phosphorus efficiency in soybean[J]. Functional Plant Biology, 2010, 37(4): 304-312. doi: 10.1071/FP09215
[24] 阮文亮, 彭松, 祝晓慧, 等. 减量施氮与间作大豆对甜玉米土壤团聚体及有机碳含量的影响[J]. 四川农业大学学报, 2023, 41(5): 811-819. [25] 白伟, 孙占祥, 郑家明, 等. 东北风沙半干旱区仁用杏作物间作对作物产量和土地生产力的影响[J]. 生态学杂志, 2017, 36(9): 2521-2528. [26] 董宛麟, 张立祯, 于洋, 等. 向日葵和马铃薯间作模式的生产力及水分利用[J]. 农业工程学报, 2012, 28(18): 127-133. [27] YANG W, LI Z, WANG J, et al. Crop yield, nitrogen acquisition and sugarcane quality as affected by interspecific competition and nitrogen application[J]. Field Crops Research, 2013, 146: 44-50. doi: 10.1016/j.fcr.2013.03.008
[28] BARRY K E, MOMMER L, VAN RUIJVEN J, et al. The future of complementarity: Disentangling causes from consequences[J]. Trends in Ecology & Evolution, 2019, 34(2): 167-180.
[29] LI C, HOFFLAND E, KUYPER T W, et al. Syndromes of production in intercropping impact yield gains[J]. Nature Plants, 2020, 6(6): 653-660 doi: 10.1038/s41477-020-0680-9
[30] YU X, XIAO S, YAN T, et al. Interspecific competition as affected by nitrogen application in sweet corn-soybean intercropping system[J]. Agronomy, 2023, 13(9): 2268. doi: 10.3390/agronomy13092268
[31] 刘天学, 张绍芬, 赵霞, 等. 我国玉米主要间作技术研究进展[J]. 河南农业科学, 2008(5): 14-17. [32] BETENCOURT E, DUPUTEL M, COLOMB B, et al. Intercropping promotes the ability of durum wheat and chickpea to increase rhizosphere phosphorus availability in a low P soil[J]. Soil Biology and Biochemistry, 2012, 46: 181-190. doi: 10.1016/j.soilbio.2011.11.015
[33] TANG X, PLACELLA S A, DAYDÉ F, et al. Phosphorus availability and microbial community in the rhizosphere of intercropped cereal and legume along a P-fertilizer gradient[J]. Plant and Soil, 2016, 407(1/2): 119-134.
[34] BEDOUSSAC L, JUSTES E. The efficiency of a durum wheat-winter pea intercrop to improve yield and wheat grain protein concentration depends on N availability during early growth[J]. Plant and Soil, 2010, 330(1/2): 19-35.
[35] ADAMS A E, BESOZZI E M, SHAHROKHI G, et al. A case for associational resistance: Apparent support for the stress gradient hypothesis varies with study system[J]. Ecology Letters, 2022, 25(1): 202-217. doi: 10.1111/ele.13917
[36] LI L, LI S, SUN J, et al. Diversity enhances agricultural productivity via rhizosphere phosphorus facilitation on phosphorus-deficient soils[J]. Proceedings of the National Academy of Sciences of the United States of America, 2007, 104(27): 11192-11196. doi: 10.1073/pnas.0704591104
[37] 安迪, 杨令, 王冠达, 等. 磷在土壤中的固定机制和磷肥的高效利用[J]. 化工进展, 2013, 32(8): 1967-1973. [38] HODGE A, BERTA G, DOUSSAN C, et al. Plant root growth, architecture and function[J]. Plant and Soil, 2009, 321(1/2): 153-187.
[39] 任媛媛, 李朝怡, 闫敏飞, 等. 黄土塬区玉米||大豆间作对根系形态的影响[J]. 草业科学, 2021, 38(12): 2449-2459. [40] 李隆. 间套作强化农田生态系统服务功能的研究进展与应用展望[J]. 中国生态农业学报, 2016, 24(4): 403-415. [41] LUO W, NI M, WANG Y, et al. Limited evidence of vertical fine-root segregation in a subtropical forest[J]. New Phytologist, 2021, 231(6): 2308-2318. doi: 10.1111/nph.17546
[42] ZHANG D, LI H, SHEN J. Strategies for root’s foraging and acquiring soil nutrient in high efficiency under intensive cropping systems[J]. Journal of Plant Nutrition and Fertilizers, 2017, 23(6): 1547-1555.
 下载:
下载: