
Diversity of Ralstonia solanacearum strains from tomato in the south of Jiangxi Province
-
摘要:目的
分离鉴定赣南地区番茄青枯病菌,明确菌系分化,为当地番茄抗青枯病育种和病害防治奠定基础。
方法从江西省赣南地区采集番茄青枯病病株,经选择性平板分离、纯化和分子鉴定,获得不同地理来源的青枯菌Ralstonia solanacearum菌株。通过生理生化测定和接种番茄试验,鉴定青枯菌的生化变种和致病类型。PCR扩增内切葡聚糖酶基因egl序列,明确青枯菌的演化型和序列变种。双层平板培养法测定其对8个不同噬菌体的敏感性。
结果获得了来自赣南地区9个市(县)的番茄青枯菌菌株44个,其中,41个菌株为生化变种Ⅲ,3个菌株为生化变种Ⅳ;致病力测定结果聚为I、II和III类,其致病力分别为强、中和弱,其中,强致病力菌株占65.9%。所有菌株属于亚洲分支演化型(Ⅰ),并进一步划分为Sequevar13、14、15、17、18、34、44和48等8个序列变种。大部分菌株对供试的8个噬菌体敏感。
结论赣南地区番茄青枯菌以生化变种III和强致病力菌株为主,对噬菌体较敏感,存在8个序列变种,具有明显的菌系分化现象和遗传多样性。
Abstract:ObjectiveIsolating and identifying Ralstonia solanacearum strains from tomato plants in the Southern of Jiangxi Province, and clarifying the bacterial differentiation can lay the foundation for local tomato bacterial wilt resistance breeding and disease control.
MethodThe diseased tomato plants were collected from the south of Jiangxi Province, R. solanacearum strains with different geographical origins were isolated by selective plate, purificated and identificated by PCR. The test of physiology and biochemistry and inoculation on tomato plants were conducted for the determination of biovar and virulence difference. The endoglucanase gene (egl) fragments were amplified by PCR to determine the phylotype and sequevar of R. solanacearum.
ResultA total of 44 R. solanacearum strains were obtained from nine cities (counties) in the south of Jiangxi Province, among which 41 strains were identified as biovar III and three strains were identified as biovar IV. According to the results of virulence difference, 44 strains were clustered into three groups, namely group I (high virulence), group II (moderate virulence) and group III (weak virulence), of which group I (high virulence) strains accounted for 65.9%. All strains were belonged to the phylotype I and further divided into eight sequevars, namely Sequevar 13, 14, 15, 17, 18, 34, 44 and 48 respeclively. Most R. solanacearum strains were sensitive to the eight tested bacteriophages.
ConclusionThe strains of R. solanacearum from tomato in the south of Jiangxi Province are mainly biovar III and high virulence, sensitive to bacteriophages, have eight sequevars, and have obvious differentiation and genetic diversity.
-
Keywords:
- Ralstonia solanacearum /
- Tomato bacterial wilt /
- Biovar /
- Virulence /
- Phylotype /
- Sequevar /
- Bacteriophage
-
茄科雷尔氏菌Ralstonia solanacearum俗称青枯菌,是危害严重的世界性植物病原细菌[1],广泛分布于热带、亚热带、温带地区,寄主范围极广[2]。青枯菌侵染番茄引起的番茄青枯病,在我国南方番茄产区普遍发生且危害严重[3-4]。
青枯菌具有高度变异性及适应性,菌系分化明显,不同菌株间基因组变异性较强。明确不同来源青枯菌的致病力分化,探索青枯菌的种内遗传多样性,具有重要意义。关于青枯菌的种下分类,Hayward等[5]、华静月等[6]根据3种双糖和3种己醇的利用能力不同,划分出5个生化变种或生化型。Prior等[7]根据地理起源的密切相关性,将青枯菌分为4个演化型,即亚洲分支演化型(Ⅰ)、美洲分支演化型(Ⅱ)、非洲分支演化型(Ⅲ)和印度尼西亚分支演化型(Ⅳ),而在演化型内根据内切葡聚糖酶(Endoglucanase)基因egl序列的差异,又细分为多个序列变种(Sequevar)。目前,国际上已经鉴定出51个序列变种,来自中国的青枯菌属于演化型Ⅰ和Ⅱ,以及1、12、13、14、15、16、17、18、34、44和48等11个序列变种[8]。尽管我国有些省份对番茄青枯病菌有研究报道[9],但来自江西省的番茄青枯菌菌系研究较少。
近年来,在江西省赣南地区种植番茄、辣椒和茄子等茄科蔬菜面积扩大,发展态势迅猛,然而,由于青枯病的严重危害,给当地蔬菜生产造成了巨大的经济损失。目前,番茄青枯病的防治尚无有效的防治药剂,虽然有研究表明选择合适的噬菌体或噬菌体组合对作物青枯病具有较好的防治作用[10-12],但其应用仍受到许多限制。选育抗病或耐病品种是青枯病防治最经济有效的方法,但由于青枯菌菌系分化明显,且与寄主、环境之间构成复杂的互作关系,使得生产上推广的抗病品种往往随着种植年限的延长、青枯菌菌系致病力的变化,导致品种的抗性丧失[13-15],因此,抗病育种工作任务艰巨。分离赣南地区的番茄青枯菌菌株,建立该地区青枯菌资源库,明确青枯菌的生化变种、致病力、序列变种以及对噬菌体的敏感性,将有助于解析青枯病的发生流行机理、指导番茄抗青枯病育种和制定相关的病害防治措施。
1. 材料与方法
1.1 供试培养基及噬菌体
固体培养基:牛肉膏3 g·L−1,酵母膏3 g·L−1,蛋白胨3 g·L−1,硫酸镁0.25 g·L−1,磷酸氢二钾2 g·L−1,磷酸二氢钾0.5 g·L−1,蔗糖15 g·L−1,琼脂粉18 g·L−1。
半固体培养基:牛肉膏3 g·L−1,酵母膏3 g·L−1,蛋白胨3 g·L−1,硫酸镁0.25 g·L−1,磷酸氢二钾2 g·L−1,磷酸二氢钾0.5 g·L−1,蔗糖15 g·L−1,琼脂粉8 g·L−1。
LB液体培养基:酵母提取物5 g·L−1,氯化钠10 g·L−1,胰蛋白胨10 g·L−1。
生化变种鉴定基础培养基:蛋白胨1.0 g·L−1,磷酸二氢胺 1.0 g·L−1,氯化钾 0.2 g·L−1,七水硫酸镁0.2 g·L−1,溴百里酚蓝指示剂3.0 ml·L−1,pH调至7.0。
2,3,5−三苯基氯化四氮唑(TTC)培养基:水解干酪素1 g·L−1,蛋白胨10 g·L−1,甘油5 mL·L−1,琼脂粉32 g·L−1,使用前,每1 L培养基加5 mL 质量浓度为10 g·L−1的TTC溶液。
牛肉膏蛋白胨(NA)液体培养基:牛肉浸膏3 g·L−1,葡萄糖10 g·L−1,蛋白胨5 g·L−1,酵母粉0.5 g·L−1,pH调至7.0。
供试噬菌体:编号分别为P1556-1、P1556-2、P7-1、P574、P1521、P1555-L、P1555-1和P1555-M,分离自江西、广东等地作物青枯病土壤,由华南农业大学植物细菌研究室提供并保存。
1.2 青枯菌的分离纯化与保存
2019—2020年,在番茄青枯病发病高峰期间,前往江西省赣南地区各市、县区采集番茄青枯病标本,用TTC选择性培养基分离青枯菌,30 ℃条件下培养24~48 h,挑取青枯菌典型单菌落,在TTC平板上划线纯化,继代3次,获得纯化菌株。用青枯菌特异性引物759(5′-GTCGCCGTCAACTCACTTTCC-3′)和760(5′-GTCGCCGTCAGCAATGCGGAATCG-3′)[7]进行PCR鉴定,将扩增得到280 bp特异目的片段的青枯菌菌株,−80 ℃甘油保存。同时,用细菌基因组DNA提取试剂盒,提取各菌株DNA,−20 ℃保存备用。
1.3 青枯菌生化变种测定
参照Hayward等[5]、华静月等[6]方法。分别将乳糖、甘露醇、山梨醇和甜醇加入基础培养基中,质量浓度均为10 g·L−1,分装试管,每管4~5 mL,110 ℃条件下灭菌20 min。纤维二糖和麦芽糖经过滤灭菌后,分别加入灭菌的基础培养基中,质量浓度为10 g·L−1。每个菌株每种化合物接种3支试管,以不接种青枯菌为对照,28 ℃条件下培养21 d。
1.4 青枯菌致病性测定及数据处理
参照何自福等[16]的方法,选择对青枯病表现不同抗性的5个番茄品种,分别为红圣佳二号(抗病)、金艳(中抗)、多宝(中抗)、粉霸(感病)和精棚T红(高感),作为致病性分化的鉴别品种。将健康番茄种子播于穴盘消毒基质中,待4~5片叶苗龄时用于接种。采用浸根接种方法,将番茄根部浸于1×108 CFU·mL−1青枯菌悬液中,15 min后移栽到盆钵中,每盆种2株,以浸无菌水的植株为空白对照,每个菌株接种每个番茄品种20株,定期记录发病情况,接种后第45天,各处理病情稳定,统计发病率。采用离差平均和的系统聚类方法(即Ward聚类)对数据进行聚类分析。
1.5 egl基因扩增和系统发育树构建
内源葡聚糖酶基因(egl基因)扩增引物:Endo-F(5′-ATGCATGCCGCTGGTCGCCGC-3′)和Endo-R(5′-GCGTTGCCCGGCACGAACACC-3′)。PCR扩增反应程序:96 ℃预变性9 min;95 ℃变性1 min,70 ℃退火1 min,72 ℃延伸2 min,30个循环;72 ℃延伸10 min,扩增产物交由睿博兴科生物技术公司进行测序,将序列提交到GenBank数据库,并与相关序列进行比对(参考序列信息见表1)。用Clustalx和MEGA软件进行系统发育分析,采用Jukes and Cantor模型邻接法(Neighbor-joining,NJ)构建系统发育树,1000次重复Bootstrap统计学检验后构建发育树。
表 1 参考序列信息Table 1. Referenced sequence information参考菌株
Reference strain寄主
Host来源
Origin演化型
Phylotype序列变种
Sequevaregl登录号
Accession numberJT523 马铃薯 Solanum tuberosum 留尼汪岛 Reunion Ⅰ 13 AF295252 PSS8 番茄 S. lycopersicum 中国 China Ⅰ 14 FJ561066 PSS358 番茄 S. lycopersicum 中国 China Ⅰ 15 EU407298 UW151 姜 Zingiber officinale 澳大利亚 Australia Ⅰ 16 AF295254 P11 花生 Arachis hypogaea 中国 China Ⅰ 17 FJ561068 GMI1000 番茄 S. lycopersicum 法国 France Ⅰ 18 AF295251 JT519 天竺葵 Pelargonium hortorum 留尼汪岛 Reunion Ⅰ 31 GU295032 PSS219 番茄 S. lycopersicum 中国 China Ⅰ 34 FJ561167 O3 橄榄树 Olea europaea 中国 China Ⅰ 44 FJ561069 TB28 烟草 Nicotiana tabacum 中国 China Ⅰ 44 FJ561127 Tb43 烟草 N. tabacum 中国 China Ⅰ 44 FJ561129 BdlI 木槿 Hibiscus syriacus 中国 China Ⅰ 44 FJ561098 CIIP365 马铃薯 S. tuberosum 菲律宾 The Philippines Ⅰ 45 GQ907151 MADI7 辣椒 Capsicum annuum 马达加斯加 Madagascar Ⅰ 46 GU295040 GMI8254 番茄 S. lycopersicum 印度尼西亚 Indonesia Ⅰ 47 GU295014 M2 桑树 Morus alba 中国 China Ⅰ 48 FJ561067 CMR87 番茄 S. lycopersicum 喀麦隆 Cameroon Ⅱ 35 EF439727 CMR12 番茄 S. lycopersicum 喀麦隆 Cameroon Ⅱ 52 EF439725 CMR39 番茄 S. lycopersicum 喀麦隆 Cameroon Ⅱ 41 EF439726 CFBP2972 马铃薯 S. tuberosum 马提尼克 Martinique Ⅱ 35 EF371809 UW551 天竺葵 P. hortorum 肯尼亚 Kenya Ⅱ 1 DQ657596 ICMIP7963 马铃薯 S. tuberosum 肯尼亚 Kenya Ⅱ 7 AF295263 续表 1 Continued table 1 参考菌株
Reference strain寄主
Host来源
Origin演化型
Phylotype序列变种
Sequevaregl登录号
Accession numberUW162 香蕉 Musa nana 秘鲁 Peru Ⅱ 4 AF295256 MOLK2 香蕉 M. nana 菲律宾 The Philippines Ⅱ 3 EF371841 CMR66 木龙葵 S. scabrum 喀麦隆 Cameroon Ⅲ 49 EF439729 JT525 天竺葵 P. hortorum 留尼汪岛 Reunion Ⅲ 19 AF295272 CFBP3059 茄子 S. melongena 布基纳法索 Burkina Faso Ⅲ 23 AF295270 NCPPB332 马铃薯 S. tuberosum 津巴布韦 Zimbabwe Ⅲ 22 DQ657649 MAFF301558 马铃薯 S. tuberosum 日本 Japan Ⅳ 8 AY465002 Psi 番茄 S. lycopersicum 印度尼西亚 Indonesia Ⅳ 10 EF371804 ACH732 番茄 S. lycopersicum 澳大利亚 Australia Ⅳ 11 GQ907150 1.6 噬菌体敏感性测定
噬菌体培养:在15 mL的LB液体培养基中,将青枯菌与噬菌体按照体积比1︰1混匀,在30 ℃、180 r·min−1条件下培养过夜。
双层平板制备:取青枯菌菌液100 μL加入15 mL、50 ℃的半固体培养基,迅速混匀倒入固体培养基平板上,制成双层平板,点接3 μL噬菌体液,30 ℃条件下培养24 h,观察噬菌斑产生情况。根据青枯菌被侵染的噬菌体数量(n/个)判别青枯菌对噬菌体的敏感性,标准:n≤2,敏感性弱;3≤n≤5,敏感性中等;6≤n≤7,敏感性强;n≥8,敏感性特强。
2. 结果与分析
2.1 青枯菌的分离纯化
2019—2020年,在江西省赣南地区各市、县番茄种植地均有青枯病发生,采集自于都、上犹、石城、瑞金等9个县(市)的番茄青枯病病株中,共分离和鉴定出44个青枯菌株,其中于都县9个,上犹县7个,石城县4个,瑞金市2个,大余县3个,安远县3个,会昌县6个,兴国县2个,全南县8个。
2.2 青枯菌的生化变种鉴定
根据对6种碳水化合物的利用情况(表2),44个番茄青枯菌可划分为生化变种Ⅲ和Ⅳ,其中,来自大余县的3个青枯菌菌株为生化变种Ⅳ,其余的41个青枯菌均为生化变种Ⅲ,表明赣南地区番茄青枯菌以生化变种Ⅲ为主。
表 2 青枯菌生化变种鉴定1)Table 2. Biovar identification of Ralstonia solanacearum来源
Origin菌株编号
No. of strain菌株数/个
Strain quantity麦芽糖
Maltose纤维二糖
Cellobiose乳糖
Lactose甘露醇
Mannitol山梨醇
Sorbitol甜醇
Dulcitol生化变种
Biovar于都县
Yudu CountyTm1901~Tm1908、
Tm1920~Tm19249 + + + + + + Ⅲ 上犹县
Shangyou CountyTm1913~Tm1919 7 + + + + + + Ⅲ 石城县
Shicheng CountyTm1925~Tm1929 4 + + + + + + Ⅲ 瑞金市
Ruijin CityTm1930、Tm1931 2 + + + + + + Ⅲ 大余县
Dayu CountyTm1932~Tm1934 3 − − − + + + Ⅳ 安远县
Anyuan CountyTm1935~Tm1937 3 + + + + + + Ⅲ 会昌县
Huichang CountyTm1938~Tm1943 6 + + + + + + Ⅲ 兴国县
Xingguo CountyTm2046、Tm2047 2 + + + + + + Ⅲ 全南县
Quannan CountyTm1944、Tm1945、
Tm2048~Tm20588 + + + + + + Ⅲ 1)“+”表示被利用,“−”表示不被利用
1)“+”indicates to be used, “−” indicates not to be used2.3 青枯菌的致病力测定
青枯菌接种红圣佳二号(抗病)、金艳(中抗)、多宝(中抗)、粉霸(感病)、精棚T红(高感)5个抗性程度不同的番茄品种,结果表明,44个菌株在不同的番茄品种上的致病力存在明显差异(表3)。
表 3 44个青枯菌接种5个番茄品种的发病率及聚类分组Table 3. Incidence of 44 Ralstonia solanacearum strains inoculated to five tomato cultivars and their clustering results来源
Origin菌株编号
No. of strain发病率/% Incidence rate 聚类分组
Cluster红圣佳2号
Hongshengjia 2金艳
Jinyan多宝
Duobao粉霸
Fenba精棚T红
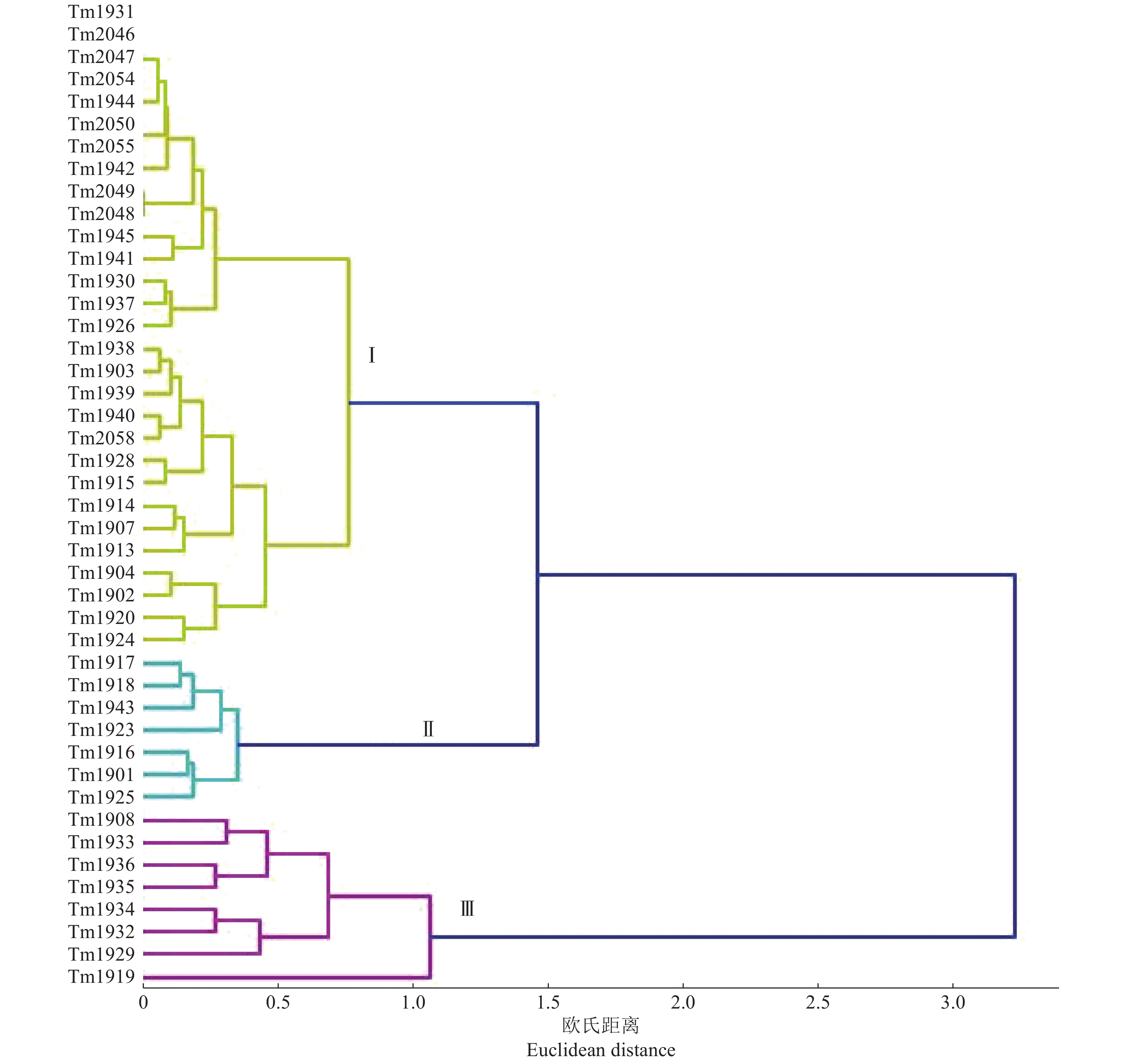
Jingpeng T red于都县 Yudu County Tm1901 55 80 95 100 95 Ⅱ Tm1902 90 85 75 100 100 Ⅰ Tm1903 85 80 95 100 100 Ⅰ Tm1904 90 95 70 100 100 Ⅰ Tm1907 85 75 80 100 100 Ⅰ Tm1908 37 65 60 72 90 Ⅲ 上犹县 Shangyou County Tm1913 90 70 90 100 90 Ⅰ Tm1914 95 75 85 95 100 Ⅰ Tm1915 85 75 95 90 100 Ⅰ Tm1916 60 75 90 85 95 Ⅱ Tm1917 40 70 85 90 100 Ⅱ Tm1918 50 70 95 90 100 Ⅱ Tm1919 0 5 15 40 65 Ⅲ 于都县 Yudu County Tm1920 80 85 80 90 90 Ⅰ Tm1923 50 80 95 68 95 Ⅱ Tm1924 80 95 85 85 100 Ⅰ 石城县 Shicheng County Tm1925 65 85 85 95 85 Ⅱ Tm1926 95 100 90 100 100 Ⅰ Tm1928 80 75 100 95 100 Ⅰ Tm1929 35 21 40 50 95 Ⅲ 瑞金市 Ruijin City Tm1930 90 95 95 100 100 Ⅰ Tm1931 100 100 100 100 100 Ⅰ 大余县 Dayu County Tm1932 20 35 65 45 80 Ⅲ Tm1933 35 35 55 80 90 Ⅲ Tm1934 20 40 75 60 100 Ⅲ 安远县 Anyuan County Tm1935 40 52 85 95 100 Ⅲ Tm1936 30 35 90 80 89 Ⅲ Tm1937 85 95 90 100 95 Ⅰ 会昌县 Huichang County Tm1938 90 85 95 100 100 Ⅰ Tm1939 80 85 95 100 95 Ⅰ Tm1940 85 90 100 95 100 Ⅰ Tm1941 100 84 100 100 100 Ⅰ Tm1942 95 100 100 100 95 Ⅰ Tm1943 40 85 95 90 100 Ⅱ 全南县 Quannan County Tm1944 100 100 95 100 100 Ⅰ Tm1945 100 90 100 90 100 Ⅰ 兴国县 Xingguo County Tm2046 100 100 100 100 100 Ⅰ Tm2047 100 100 100 100 100 Ⅰ 全南县 Quannan County Tm2048 100 95 100 100 89 Ⅰ Tm2049 100 95 100 100 90 Ⅰ Tm2050 100 95 100 100 100 Ⅰ Tm2054 100 100 100 100 100 Ⅰ Tm2055 100 95 100 100 100 Ⅰ Tm2058 90 90 100 100 100 Ⅰ 采用系统聚类中的离差平均和的聚类方法(即Ward聚类),对44个菌株接种5个番茄品种的青枯病发病率结果进行聚类分析,明确青枯菌之间的致病力差异。采用离差平均和方法,样本间的距离采用欧氏距离,该距离数值的大小反映样本之间的相似度,数值越小,2个样本之间的相似度越高,数值越大,则相似度越低。本研究以欧氏距离1.2为参考标准,将44个青枯菌聚为3个组(图1)。第Ⅰ组共有29个菌株,分别来自除大余县外的8个县(市);第Ⅱ组的7个菌株分离自于都、上犹、石城、会昌4个县,该组致病力为中等强度;第Ⅲ组的8个菌株分离自于都、上犹、石城、大余、安远5个县,致病力弱,其中来自上犹县的Tm1919致病力最弱。上述结果表明,赣南地区番茄青枯菌致病力分化现象明显,在于都、上犹和石城县均存在致病力强、中、弱3种类型的菌株,但总体来看,赣南地区番茄青枯菌以致病较强的菌株占优势。
![]() 图 1 基于44个青枯菌接种5个番茄品种发病率的聚类分析结果Figure 1. Clustering results based on the incidence of 44 Ralstonia solanacearum strains inoculated to five tomato cultivars
图 1 基于44个青枯菌接种5个番茄品种发病率的聚类分析结果Figure 1. Clustering results based on the incidence of 44 Ralstonia solanacearum strains inoculated to five tomato cultivars2.4 青枯菌序列变种及系统发育分析
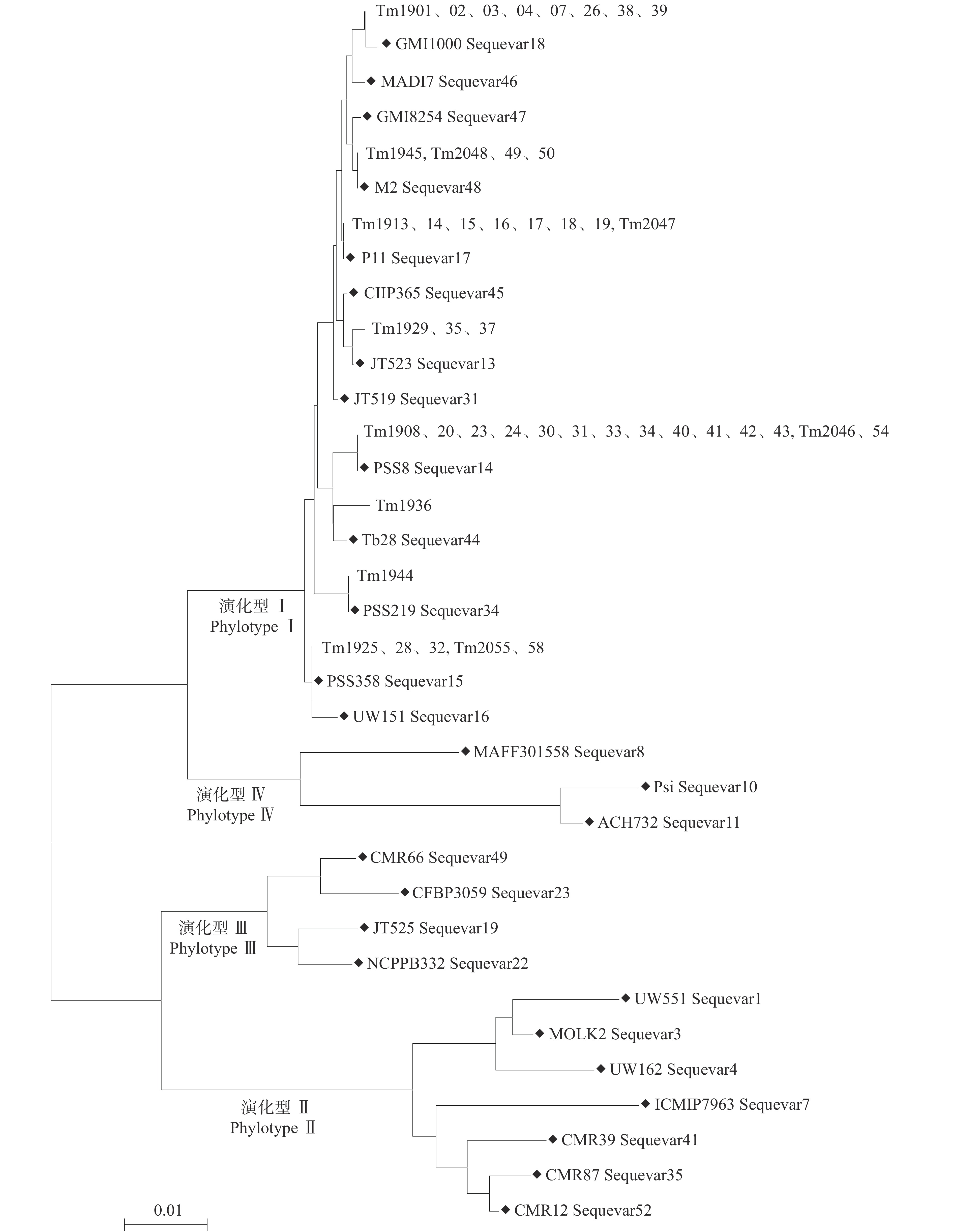
44个青枯菌的egl基因片段序列与标准参考菌株进行比对,构建系统发育树(图2)。结果表明,江西省赣南地区的番茄青枯菌属于亚洲分支、演化型I(PhylotypeⅠ)的8个序列变种(Sequevar)。其中Tm1929、Tm1935和Tm1937与马铃薯青枯菌JT523(留尼汪岛)属于Sequevar 13;Tm1908、Tm1920、Tm1923、Tm1924、Tm1930、Tm1931、Tm1933、Tm1934、Tm1940~Tm1943、Tm2046、Tm2054共14个菌株与来自中国的番茄青枯菌PSS8属于Sequevar 14;Tm1925、Tm1928、Tm1932、Tm2055和Tm2058与来自中国的番茄青枯菌PSS358属于Sequevar 15;Tm1913~Tm1919、Tm2047共8个菌株与来自中国的花生青枯菌P11属于Sequevar 17;Tm1901~Tm1904、Tm1907、Tm1926、Tm1938、Tm1939共8个菌株与来自法国的番茄青枯菌GMI1000均为Sequevar 18;Tm1944与来自中国台湾的番茄青枯菌PSS219为Sequevar 34;Tm1936与来自中国的烟草青枯菌Tb28为Sequevar 44;Tm1945、Tm2048~Tm2050共4个菌株与来自中国的桑青枯菌M2同为Sequevar 48。
![]() 图 2 基于青枯菌egl基因序列的系统发育分析将序列变种相同的菌株置于同一水平分支上,“◆”代表参考菌株Figure 2. Phylogenetic analysis based on sequence of egl gene of Ralstonia solanacearumStrains with the same sequevar are settled at the same branch, “◆” indicates reference strain
图 2 基于青枯菌egl基因序列的系统发育分析将序列变种相同的菌株置于同一水平分支上,“◆”代表参考菌株Figure 2. Phylogenetic analysis based on sequence of egl gene of Ralstonia solanacearumStrains with the same sequevar are settled at the same branch, “◆” indicates reference strain2.5 青枯菌序列变种(Sequevar)的地理分布
在所分离的44个番茄青枯菌中,Sequevar 14共有14个菌株,分布于于都、瑞金、大余、会昌、兴国、全南6个县(市);Sequevar 18有8个菌株,分布于会昌、石城、于都3个县;Sequevar 17有8个菌株,其中7个分布于上犹县,1个来自兴国县;Sequevar 15有5个菌株,来自于石城、大余、全南3个县;Sequevar 48的4个菌株,全部来自全南县;Sequevar 13有3个菌株,分布于石城和安远县;Sequevar 34和Sequevar 44各有1个菌株,分别来自全南和安远县。上述结果表明,赣南地区番茄青枯菌序列变种存在丰富的多样性。
2.6 青枯菌对噬菌体的敏感性测定
双层平板法检测8个噬菌体裂解青枯菌的试验结果(表4)表明,44个番茄青枯菌中,只有菌株Tm2047对噬菌体的敏感性弱,有12个菌株对噬菌体的敏感性表现为中等,有4个菌株对噬菌体的敏感性较强,另有27个菌株都能被8个噬菌体裂解,其敏感性表现为特强。上述结果表明,赣南地区番茄青枯菌对噬菌体普遍表现敏感。
表 4 44个青枯菌对8个噬菌体的敏感性测定Table 4. Sensitivity determination of 44 Ralstonia solanacearum to eight bacteriophages来源
Origin菌株编号
No. of strain数量/个
Quantity噬菌体1) Bacteriophage 敏感性2)
SensitivityP1555-L P1555-1 P1555-M P1556-1 P1556-2 P7-1 P574 P1521 于都县
Yudu CountyTm1901~Tm1907 5 – – – + + + – + M Tm1908 1 + + + + + + + + V 上犹县
Shangyou CountyTm1913、Tm1914 2 + + + + – + + + S Tm1915~Tm1919 4 + + + + + + + + V Tm1916 1 + + + + – + + + S 于都县
Yudu CountyTm1920~Tm1924 3 + + + + + + + + V 石城县
Shicheng CountyTm1925~Tm1929 3 + + + + + + + + V Tm1928 1 + + + + – + + + S 瑞金市
Ruijin CityTm1930、Tm1931 2 + + + + + + + + V 大余县
Dayu CountyTm1932~Tm1934 3 + + + + + + + + V 安远县
Anyuan CountyTm1935~Tm1937 3 + + + + + + + + V 会昌县
Huichang CountyTm1938、Tm1939 2 – – – + + + – + M Tm1940~Tm1943 4 + + + + + + + + V 全南县
Quannan CountyTm1944 1 – – – + + + + + M Tm1945 1 – – – – – + + + M 兴国县
Xingguo CountyTm2046 1 + + + + + + + + V Tm2047 1 – – – + – + – – W 全南县
Quannan CountyTm2048、Tm2049 2 – – – + + + + + M Tm2050 1 – – – – + + + + M Tm2054~Tm2058 3 + + + + + + + + V 1)“+”表示产生噬菌斑,“–”表示不产生噬菌斑;2)M:中等,S:强,V:特强,W:弱
1) “+” indicates having plaques,“–” indicates no having plaques;2) M: Moderate, S: Strong, V:Very strong, W: Weak3. 讨论与结论
不同地理来源的青枯菌在与寄主长期协同进化的过程中,演化出明显的生理分化型或菌系多样性。国外报道,番茄青枯菌属于1号生理小种,分为演化型Ⅰ、Ⅱ、Ⅲ和Ⅳ[17-18],生化变种Ⅰ、II、III和IV[19-20],存在Sequevar 1、4、5、6、7、8、9、10、11、13、14、15、18、20、29、31、35、41、38、39、46、52等22个序列变种[21-23]。在我国,已报道的番茄青枯菌属于1号生理小种、演化型Ⅰ,分为生化变种II、III和IV,存在Sequevar13、14、15、16、17、18、34、44、48和54等10个序列变种[24-25]。曾宪铭等[26]曾测定广东省13种农作物上的129个青枯菌,其生化变种鉴定为Ⅰ、Ⅱ、Ⅲ、Ⅳ及其亚型,其中番茄青枯菌为生化变种Ⅲ和Ⅳ,郑向华等[27]也获得类似的研究结果,将番茄青枯菌鉴定为生化变种Ⅲ。2009年,Xu等[28]基于演化型分类框架,对来自我国13个省、17种不同寄主植物上的286株青枯菌进行多样性分析,明确了我国青枯病菌具有丰富的遗传多样性,其中番茄青枯菌属于演化型Ⅰ,并有Sequevar 13、14、15、16、17、18、44等7个序列变种。2011年,Xue等[24]分析了我国15个省、14种寄主植物上的319株青枯菌,基于PCR指纹图谱及演化型框架,将其中的番茄青枯菌鉴定为演化型Ⅰ,存在Sequevar 13、14、15、17、18、34、44、48等8个序列变种。
本研究对江西省赣南地区的番茄青枯菌进行了分离和菌系分析,结果显示,在分离鉴定出的44个青枯菌中,有41个菌株为生化变种Ⅲ(占93.2%),另有3个菌株属于生化变种IV,不存在生化变种I和II,与国内报道的番茄青枯菌生化变种结果基本相似。尽管我国的番茄青枯菌均属于1号生理小种,但菌株之间致病力存在差异。本研究通过接种试验,根据在5个不同青枯病抗性的番茄品种上的发病情况,将来自赣南地区的番茄青枯菌分为Ⅰ、Ⅱ、Ⅲ共3个致病类型,而且致病力较强的菌株占65.91%,15.91%的菌株致病力中等,18.18%的菌株致病力较弱,可见,赣南地区番茄青枯菌的致病力存在明显差异,而且致病力较强的菌株是本地区的优势菌群,这将为赣南地区的番茄抗青枯病育种工作提供指导。演化型和egl基因序列分析结果表明,44个番茄青枯菌株均属于演化型I,符合亚洲起源的演化型Ⅰ分类框架[7],进一步研究显示,赣南地区番茄青枯菌存在8个序列变种,分别为Sequevar 13、14、15、17、18、34、44和48,未发现Sequevar 16,其中 Sequevar 14(占31.8%)在本地区的番茄主要种植地区均有分布,表明赣南地区番茄青枯菌存在丰富的遗传多样性。
噬菌体是侵染细菌的病毒,具有高度的特异性及快速裂解宿主(寄主)的能力,可用于植物细菌病害的防治[29-31]。有研究表明,通过使用噬菌体组合的方法,能够显著降低青枯菌数量,对番茄青枯病具有较好的防治效果[32]。而测定青枯菌对噬菌体的敏感性,可为利用噬菌体防治植物青枯病提供参考。本研究结果表明,在44个青枯菌中,有61.36%的青枯菌对供试的8个噬菌体敏感,即都能被8个噬菌体裂解,表明赣南地区番茄青枯菌对噬菌体普遍表现敏感。然而,仍有12个菌株对8个噬菌体的综合敏感性表现为中等,而Tm2047菌株对噬菌体不敏感。由此可见,生产上应根据本地区青枯菌菌系的多样性及对噬菌体的敏感性,有选择性地进行噬菌体组合,以减轻番茄青枯病的发生危害。
-
![]()
图 1 基于44个青枯菌接种5个番茄品种发病率的聚类分析结果
Figure 1. Clustering results based on the incidence of 44 Ralstonia solanacearum strains inoculated to five tomato cultivars
![]()
图 2 基于青枯菌egl基因序列的系统发育分析
将序列变种相同的菌株置于同一水平分支上,“◆”代表参考菌株
Figure 2. Phylogenetic analysis based on sequence of egl gene of Ralstonia solanacearum
Strains with the same sequevar are settled at the same branch, “◆” indicates reference strain
表 1 参考序列信息
Table 1 Referenced sequence information
参考菌株
Reference strain寄主
Host来源
Origin演化型
Phylotype序列变种
Sequevaregl登录号
Accession numberJT523 马铃薯 Solanum tuberosum 留尼汪岛 Reunion Ⅰ 13 AF295252 PSS8 番茄 S. lycopersicum 中国 China Ⅰ 14 FJ561066 PSS358 番茄 S. lycopersicum 中国 China Ⅰ 15 EU407298 UW151 姜 Zingiber officinale 澳大利亚 Australia Ⅰ 16 AF295254 P11 花生 Arachis hypogaea 中国 China Ⅰ 17 FJ561068 GMI1000 番茄 S. lycopersicum 法国 France Ⅰ 18 AF295251 JT519 天竺葵 Pelargonium hortorum 留尼汪岛 Reunion Ⅰ 31 GU295032 PSS219 番茄 S. lycopersicum 中国 China Ⅰ 34 FJ561167 O3 橄榄树 Olea europaea 中国 China Ⅰ 44 FJ561069 TB28 烟草 Nicotiana tabacum 中国 China Ⅰ 44 FJ561127 Tb43 烟草 N. tabacum 中国 China Ⅰ 44 FJ561129 BdlI 木槿 Hibiscus syriacus 中国 China Ⅰ 44 FJ561098 CIIP365 马铃薯 S. tuberosum 菲律宾 The Philippines Ⅰ 45 GQ907151 MADI7 辣椒 Capsicum annuum 马达加斯加 Madagascar Ⅰ 46 GU295040 GMI8254 番茄 S. lycopersicum 印度尼西亚 Indonesia Ⅰ 47 GU295014 M2 桑树 Morus alba 中国 China Ⅰ 48 FJ561067 CMR87 番茄 S. lycopersicum 喀麦隆 Cameroon Ⅱ 35 EF439727 CMR12 番茄 S. lycopersicum 喀麦隆 Cameroon Ⅱ 52 EF439725 CMR39 番茄 S. lycopersicum 喀麦隆 Cameroon Ⅱ 41 EF439726 CFBP2972 马铃薯 S. tuberosum 马提尼克 Martinique Ⅱ 35 EF371809 UW551 天竺葵 P. hortorum 肯尼亚 Kenya Ⅱ 1 DQ657596 ICMIP7963 马铃薯 S. tuberosum 肯尼亚 Kenya Ⅱ 7 AF295263  下载: 导出CSV
下载: 导出CSV
续表 1 Continued table 1 参考菌株
Reference strain寄主
Host来源
Origin演化型
Phylotype序列变种
Sequevaregl登录号
Accession numberUW162 香蕉 Musa nana 秘鲁 Peru Ⅱ 4 AF295256 MOLK2 香蕉 M. nana 菲律宾 The Philippines Ⅱ 3 EF371841 CMR66 木龙葵 S. scabrum 喀麦隆 Cameroon Ⅲ 49 EF439729 JT525 天竺葵 P. hortorum 留尼汪岛 Reunion Ⅲ 19 AF295272 CFBP3059 茄子 S. melongena 布基纳法索 Burkina Faso Ⅲ 23 AF295270 NCPPB332 马铃薯 S. tuberosum 津巴布韦 Zimbabwe Ⅲ 22 DQ657649 MAFF301558 马铃薯 S. tuberosum 日本 Japan Ⅳ 8 AY465002 Psi 番茄 S. lycopersicum 印度尼西亚 Indonesia Ⅳ 10 EF371804 ACH732 番茄 S. lycopersicum 澳大利亚 Australia Ⅳ 11 GQ907150
下载: 导出CSV
表 2 青枯菌生化变种鉴定1)
Table 2 Biovar identification of Ralstonia solanacearum
来源
Origin菌株编号
No. of strain菌株数/个
Strain quantity麦芽糖
Maltose纤维二糖
Cellobiose乳糖
Lactose甘露醇
Mannitol山梨醇
Sorbitol甜醇
Dulcitol生化变种
Biovar于都县
Yudu CountyTm1901~Tm1908、
Tm1920~Tm19249 + + + + + + Ⅲ 上犹县
Shangyou CountyTm1913~Tm1919 7 + + + + + + Ⅲ 石城县
Shicheng CountyTm1925~Tm1929 4 + + + + + + Ⅲ 瑞金市
Ruijin CityTm1930、Tm1931 2 + + + + + + Ⅲ 大余县
Dayu CountyTm1932~Tm1934 3 − − − + + + Ⅳ 安远县
Anyuan CountyTm1935~Tm1937 3 + + + + + + Ⅲ 会昌县
Huichang CountyTm1938~Tm1943 6 + + + + + + Ⅲ 兴国县
Xingguo CountyTm2046、Tm2047 2 + + + + + + Ⅲ 全南县
Quannan CountyTm1944、Tm1945、
Tm2048~Tm20588 + + + + + + Ⅲ 1)“+”表示被利用,“−”表示不被利用
1)“+”indicates to be used, “−” indicates not to be used
下载: 导出CSV
表 3 44个青枯菌接种5个番茄品种的发病率及聚类分组
Table 3 Incidence of 44 Ralstonia solanacearum strains inoculated to five tomato cultivars and their clustering results
来源
Origin菌株编号
No. of strain发病率/% Incidence rate 聚类分组
Cluster红圣佳2号
Hongshengjia 2金艳
Jinyan多宝
Duobao粉霸
Fenba精棚T红
Jingpeng T red于都县 Yudu County Tm1901 55 80 95 100 95 Ⅱ Tm1902 90 85 75 100 100 Ⅰ Tm1903 85 80 95 100 100 Ⅰ Tm1904 90 95 70 100 100 Ⅰ Tm1907 85 75 80 100 100 Ⅰ Tm1908 37 65 60 72 90 Ⅲ 上犹县 Shangyou County Tm1913 90 70 90 100 90 Ⅰ Tm1914 95 75 85 95 100 Ⅰ Tm1915 85 75 95 90 100 Ⅰ Tm1916 60 75 90 85 95 Ⅱ Tm1917 40 70 85 90 100 Ⅱ Tm1918 50 70 95 90 100 Ⅱ Tm1919 0 5 15 40 65 Ⅲ 于都县 Yudu County Tm1920 80 85 80 90 90 Ⅰ Tm1923 50 80 95 68 95 Ⅱ Tm1924 80 95 85 85 100 Ⅰ 石城县 Shicheng County Tm1925 65 85 85 95 85 Ⅱ Tm1926 95 100 90 100 100 Ⅰ Tm1928 80 75 100 95 100 Ⅰ Tm1929 35 21 40 50 95 Ⅲ 瑞金市 Ruijin City Tm1930 90 95 95 100 100 Ⅰ Tm1931 100 100 100 100 100 Ⅰ 大余县 Dayu County Tm1932 20 35 65 45 80 Ⅲ Tm1933 35 35 55 80 90 Ⅲ Tm1934 20 40 75 60 100 Ⅲ 安远县 Anyuan County Tm1935 40 52 85 95 100 Ⅲ Tm1936 30 35 90 80 89 Ⅲ Tm1937 85 95 90 100 95 Ⅰ 会昌县 Huichang County Tm1938 90 85 95 100 100 Ⅰ Tm1939 80 85 95 100 95 Ⅰ Tm1940 85 90 100 95 100 Ⅰ Tm1941 100 84 100 100 100 Ⅰ Tm1942 95 100 100 100 95 Ⅰ Tm1943 40 85 95 90 100 Ⅱ 全南县 Quannan County Tm1944 100 100 95 100 100 Ⅰ Tm1945 100 90 100 90 100 Ⅰ 兴国县 Xingguo County Tm2046 100 100 100 100 100 Ⅰ Tm2047 100 100 100 100 100 Ⅰ 全南县 Quannan County Tm2048 100 95 100 100 89 Ⅰ Tm2049 100 95 100 100 90 Ⅰ Tm2050 100 95 100 100 100 Ⅰ Tm2054 100 100 100 100 100 Ⅰ Tm2055 100 95 100 100 100 Ⅰ Tm2058 90 90 100 100 100 Ⅰ
下载: 导出CSV
表 4 44个青枯菌对8个噬菌体的敏感性测定
Table 4 Sensitivity determination of 44 Ralstonia solanacearum to eight bacteriophages
来源
Origin菌株编号
No. of strain数量/个
Quantity噬菌体1) Bacteriophage 敏感性2)
SensitivityP1555-L P1555-1 P1555-M P1556-1 P1556-2 P7-1 P574 P1521 于都县
Yudu CountyTm1901~Tm1907 5 – – – + + + – + M Tm1908 1 + + + + + + + + V 上犹县
Shangyou CountyTm1913、Tm1914 2 + + + + – + + + S Tm1915~Tm1919 4 + + + + + + + + V Tm1916 1 + + + + – + + + S 于都县
Yudu CountyTm1920~Tm1924 3 + + + + + + + + V 石城县
Shicheng CountyTm1925~Tm1929 3 + + + + + + + + V Tm1928 1 + + + + – + + + S 瑞金市
Ruijin CityTm1930、Tm1931 2 + + + + + + + + V 大余县
Dayu CountyTm1932~Tm1934 3 + + + + + + + + V 安远县
Anyuan CountyTm1935~Tm1937 3 + + + + + + + + V 会昌县
Huichang CountyTm1938、Tm1939 2 – – – + + + – + M Tm1940~Tm1943 4 + + + + + + + + V 全南县
Quannan CountyTm1944 1 – – – + + + + + M Tm1945 1 – – – – – + + + M 兴国县
Xingguo CountyTm2046 1 + + + + + + + + V Tm2047 1 – – – + – + – – W 全南县
Quannan CountyTm2048、Tm2049 2 – – – + + + + + M Tm2050 1 – – – – + + + + M Tm2054~Tm2058 3 + + + + + + + + V 1)“+”表示产生噬菌斑,“–”表示不产生噬菌斑;2)M:中等,S:强,V:特强,W:弱
1) “+” indicates having plaques,“–” indicates no having plaques;2) M: Moderate, S: Strong, V:Very strong, W: Weak
下载: 导出CSV
-
[1] 冯洁. 植物病原细菌分类最新进展[J]. 中国农业科学, 2017, 50(12): 2305-2314. doi: 10.3864/j.issn.0578-1752.2017.12.011 [2] HAYWARD A C. Biology and epidemiology of bacterial wilt caused by Pseudomonas solanacearum[J]. Annual Review of Phytopathology, 1991, 29(1): 65-87. doi: 10.1146/annurev.py.29.090191.000433
[3] 乔俊卿, 陈志谊, 刘邮洲, 等. 茄科作物青枯病研究进展[J]. 植物病理学报, 2013, 43(1): 1-10. doi: 10.3969/j.issn.0412-0914.2013.01.001 [4] 王杰, 龙世芳, 王正文, 等. 番茄青枯病防治研究进展[J]. 中国蔬菜, 2020, 40(1): 22-30. [5] HAYWARD A. Characteristics of Pseudomonas solanacearum[J]. Journal of Applied Bacteriology, 1964, 27(2): 265-277. doi: 10.1111/j.1365-2672.1964.tb04912.x
[6] 华静月, 张长龄, 何礼远. 我国植物青枯菌的生化型和其他生理差异[J]. 植物保护学报, 1984, 11(1): 43-50. [7] PRIOR P, FEGAN M. Recent development in the phylogeny and classification of Ralstonia solanacearum[J]. Acta Horticulturae, 2005, 695(14): 127-136.
[8] 徐进, 冯洁. 植物青枯菌遗传多样性及致病基因组学研究进展[J]. 中国农业科学, 2013, 46(14): 2902-2909. doi: 10.3864/j.issn.0578-1752.2013.14.006 [9] JIANG G, WEI Z, XU J, et al. Bacterial wilt in China: History, current status, and future perspectives[J/OL]. Frontiers in Plant Science, 2017, 8: 1549. [2021-01-18]. https://doi.org/10.3389/fpls.2017.01549.
[10] ADDY H S, ASKORA A, KAWASAKI T, et al. Loss of virulence of the phytopathogen Ralstonia solanacearum through infection by φRSM filamentous phages[J]. Phytopathology, 2012, 102(5): 469-477. doi: 10.1094/PHYTO-11-11-0319-R
[11] ADDY H S, ASKORA A, KAWASAKI T, et al. The filamentous phage фRSS1 enhances virulence of phytopathogenic Ralstonia solanacearum on tomato[J]. Phytopathology, 2012, 102(3): 244-251. doi: 10.1094/PHYTO-10-11-0277
[12] LIU N, LEWIS C, ZHENG W, et al. Phage cocktail therapy: Multiple ways to suppress pathogenicity[J]. Trends in Plant Science, 2020, 25(4): 315-317. doi: 10.1016/j.tplants.2020.01.013
[13] 佘小漫, 何自福. 作物青枯病研究进展[J]. 广东农业科学, 2020, 47(12): 82-89. [14] 汪国平, 林明宝, 吴定华. 番茄青枯病抗性遗传研究进展[J]. 园艺学报, 2004, 31(3): 403-407. doi: 10.3321/j.issn:0513-353X.2004.03.033 [15] 陈胜华. 番茄青枯病生物防治策略研究[J]. 农业与技术, 2018, 38(6): 24. [16] 何自福, 虞皓, 罗方芳. 广东茄科青枯菌致病力分化及其DNA多态性分析[J]. 植物病理学报, 2003, 33(5): 415-420. doi: 10.3321/j.issn:0412-0914.2003.05.007 [17] WICKER E, GRASSART L, CORANSON-BEAUDU R, et al. Ralstonia solanacearum strains from Martinique (French West Indies) exhibiting a new pathogenic potential[J]. Applied & Environmental Microbiology, 2007, 73(21): 6790-6801.
[18] PEREZ A S, MEJIA L, FEGAN M, et al. Diversity and distribution of Ralstonia solanacearum strains in Guatemala and rare occurrence of tomato fruit infection[J]. Plant Pathology, 2010, 57(2): 320-331.
[19] NORMAN D J, ZAPATA M, GABRIEL D W, et al. Genetic diversity and host range variation of Ralstonia solanacearum strains entering North America[J]. Phytopathology, 2009, 99(9): 1070-1077. doi: 10.1094/PHYTO-99-9-1070
[20] POUSSIER S, VANDEWALLE P, LUISETTI J. Genetic diversity of African and worldwide strains of Ralstonia solanacearum as determined by PCR-restriction fragment length polymorphism analysis of the hrp gene region[J]. Applied and Environmental Microbiology, 1999, 65(5): 2184-2194. doi: 10.1128/AEM.65.5.2184-2194.1999
[21] CHESNEAU T, MAIGNIEN G, BOYER C, et al. Sequevar diversity and virulence of Ralstonia solanacearum phylotype Ⅰ on Mayotte Island (Indian Ocean)[J]. Frontiers in Plant Science, 2017, 8: 2209. Doi: 10.3389/fpls.2017.02209.
[22] JI P, ALLEN C, SANCHEZ-PEREZ A, et al. New diversity of Ralstonia solanacearum strains associated with vegetable and ornamental crops in Florida[J]. Plant Disease, 2007, 91(2): 195-203. doi: 10.1094/PDIS-91-2-0195
[23] MAHBOU SOMO TOUKAN G, CELLIER G, WICKER E, et al. Broad diversity of Ralstonia solanacearum strains in Cameroon[J]. Plant Disease, 2009, 93(11): 1123-1130. doi: 10.1094/PDIS-93-11-1123
[24] XUE Q Y, YIN Y N, YANG W, et al. Genetic diversity of Ralstonia solanacearum strains from China assessed by PCR-based fingerprints to unravel host plant-and site-dependent distribution patterns[J]. FEMS Microbiology Ecology, 2011, 75(3): 507-519. doi: 10.1111/j.1574-6941.2010.01026.x
[25] SHE X M, HE Z F, LI H P. Genetic structure and phylogenetic relationships of Ralstonia solanacearum strains from diverse origins in Guangdong Povince, China[J]. Journal of Phytopathology, 2017, 166(3): 177-186.
[26] 曾宪铭, 董春. 广东农作物青枯病菌的生化型[J]. 华南农业大学学报, 1995, 16(1): 50-53. [27] 郑向华, 杨帆, 邓海滨, 等. 我国植物青枯菌的生物型及RAPD分析[C]. //中国植物病理学会. 中国植物病理学会2008年学术年会论文集. 广州: 中国农业科学技术出版社, 2008: 329-337. [28] XU J, PAN Z C, PRIOR P, et al. Genetic diversity of Ralstonia solanacearum strains from China[J]. European Journal of Plant Pathology, 2009, 125(4): 641-653. doi: 10.1007/s10658-009-9512-5
[29] 马超, 丛聪, 王丽丽, 等. 噬菌体控制植物细菌性疾病的研究进展[J]. 中国抗生素杂志, 2017, 42(9): 749-754. doi: 10.3969/j.issn.1001-8689.2017.09.006 [30] DI LALLO G, EVANGELISTI M, MANCUSO F, et al. Isolation and partial characterization of bacteriophages infecting Pseudomonas syringae pv. actinidiae, causal agent of kiwifruit bacterial canker[J]. Journal of Basic Microbiology, 2015, 54(11): 1210-1221.
[31] WEI C H, LIU J L, MAINA A, et al. Developing a bacteriophage cocktail for biocontrol of potato bacterial wilt[J]. Virologica Sinica, 2017, 32(6): 476-484. doi: 10.1007/s12250-017-3987-6
[32] WANG X F, WEI Z, YANG K M, et al. Phage combination therapies for bacterial wilt disease in tomato[J]. Nature Biotechnology, 2019, 37(12): 1513-1520. doi: 10.1038/s41587-019-0328-3
-
期刊类型引用(6)
1. 张廷槐,张丹丹,许佰峰,唐唯,刘晶. 马铃薯青枯病抗性鉴定方法的比较及抗病种质筛选. 云南师范大学学报(自然科学版). 2025(02): 37-43 .  百度学术
百度学术
2. 施春兰,吴国星,赵长飞,杨燕通,万丽娜,田永明,刘正玲,谢永辉,詹莜国,顾小飞. 球孢白僵菌和爪哇虫草菌对烟草青枯病的防治效果. 微生物学通报. 2024(08): 2974-2985 . 百度学术
3. 陈娜,邵勤. 番茄NAC转录因子SlNAP2的克隆、表达及功能分析. 核农学报. 2023(02): 251-261 . 百度学术
4. 张晓南,王建豪,王佳丽,朱鸿维,牛亚倩,熊婷婷,张英迪,徐福. 番石榴叶挥发油的抗菌性及优化提取研究. 热带作物学报. 2023(01): 174-185 . 百度学术
5. 陈娜,詹文文,刘兴雨,石磊鑫,李若楠,谢镕,却志群. 番茄WRKY转录因子SlWRKY75的克隆、表达及功能分析. 华北农学报. 2023(06): 1-10 . 百度学术
6. 戴小秋,陈婷,吴雨霜,杨冬,万小荣,郑奕雄,杨永,谭晓丹. 广东省花生青枯病病原的分离和遗传多样性鉴定. 仲恺农业工程学院学报. 2022(03): 20-27+33 . 百度学术
其他类型引用(3)
计量
- 文章访问数: 464
- HTML全文浏览量: 9
- PDF下载量: 962
- 被引次数: 9
