
Trapping effect of inserting bamboo poles in the paddy fields on spawning of Pomacea canaliculata
-
摘要:目的
研究田间不同插竿方式对福寿螺Pomacea canaliculata产卵的影响。
方法通过插竿位置、插竿间距、插竿高度及竹竿形状4个因素研究不同的插竿方式对福寿螺产卵的诱集效率。
结果稻田插竹竿能诱集福寿螺在其上产卵,且在竹竿上收集到的福寿螺卵块数量呈现返青期>拔节期>抽穗期的变化趋势。在效果最明显的返青期,距离田埂1 m的竹竿对螺卵诱集效率达到50%以上,显著高于其他位置竹竿及对照水稻茎秆上的卵块数(P<0.05);距离田埂2 m的竹竿次之,距离田埂3 m的竹竿和对照最少。间距100 cm的竹竿诱集螺卵效率为41%,间距30 cm的为33%,显著高于间距70 cm及未插竹竿区域内水稻茎秆上的卵块数(P<0.05)。顶端距离水面高度30 cm的竹竿与距离水面高度50 cm的竹竿诱集产卵效率分别为43%和40%,显著高于高度10 cm及未插竹竿区域内水稻茎秆上的卵块数(P<0.05)。圆形竹条与扁竹竿在此时期收集得到的卵块数无显著差异。水稻移栽后,在靠近田埂1 m处和顶端距离水面30~50 cm的高度扦插1圈间距为100 cm的竹竿,在福寿螺为害盛期,螺卵发生密集的小范围区域内减小间距为30 cm沿田埂内周扦插1圈竹竿,均能达到较好的螺卵防除效果。
结论实际生产中在田间合理地扦插竹竿可有效减少福寿螺卵块,降低福寿螺繁殖率,减少螺害。本研究可为物理控螺提供一条新的有效途径。
Abstract:ObjectiveTo study different methods of inserting bamboo poles in paddy field on spawning ofPomacea canaliculata.
MethodEffects of four factors including position, distance, height and shape of inserting poles on trapping efficiency of P. canaliculata spawning were investigated.
ResultThe bamboo poles inserted in paddy field could lure P. canaliculata to spawn on them. The number of egg masses tended to decrease successively from reviving stage, jointing stage to heading stage. At reviving stage with the best trapping effect, more than 50% egg masses were found on the bamboo poles which were 1 m from the paddy ridge, and the number was significantly higher compared with bamboo poles at other positions and rice straws(control)(P<0.05). The bamboo poles at 3 m distance from the paddy ridge and rice straws had the least numbers of egg masses, followed by the bamboo poles at 2 m distance from the paddy ridge. The trapping efficiencies for bamboo pole layouts with 100 cm distance and 30 cm distance between poles were 41% and 33% respectively, which were significantly higher than those for bamboo pole layout with 70 cm distance between poles and the control (P<0.05). The trapping efficiencies for bamboo poles with the tops being 30 and 50 cm high above water surface were 43% and 40% respectively, which were significantly higher than those for bamboo poles with the tops being 10 cm high above water surface and the control (P<0.05). There was no significant difference in the numbers of egg masses between circular-shaped and flat-shaped bamboo poles. After transplanting the rice, high trapping efficiency ofP. canaliculata spawning could be obtained by inserting bamboo poles at 1 m distance from paddy ridge, with 100 cm distance between poles and the tops being 30–50 cm high above water surface. Specially, at peak attack period ofP. canaliculata, bamboo poles inserted at 30 cm distance between poles could realize efficient trapping of egg masses.
ConclusionAppropriately inserting bamboo poles in paddy field can help reduce the egg mass of P. canaliculata, hence reduce the reproductive rate of P. canaliculata and decrease the harm of P. canaliculata in practice. This research provides a new physical control approach of P. canaliculata.
-
福寿螺Pomacea canaliculata又名大瓶螺、苹果螺,原产于南美洲,20世纪80年代作为食用经济动物被迅速推广到东亚和东南亚国家[1-2]。该螺食性广、繁殖迅速、抗逆能力强,大量扩散到稻田后啃食水稻茎秆,成为危害水稻的恶性水生动物[3]。此外,福寿螺还是人畜共患寄生虫广州管圆线虫Angiostrongylus cantonensis的重要中间宿主之一,浙江温州和福建长乐等地广州管圆线虫病的暴发均由食用福寿螺引起[4]。2000年,福寿螺被世界自然保护联盟外来入侵物种专家委员会列入世界100种恶性外来入侵物种的黑名单[5]。目前国内外对福寿螺的防控研究主要集中在化学、生物及农业防治等方面[6-8],但容易产生生态问题:福寿螺对化学杀螺剂易产生抗药性;杀螺剂对其他非目标生物及环境的不利影响;引入的天敌可能造成新的生态入侵灾害;机械农耕对土表的不良影响等[9-11]。我国大部分地区福寿螺在3月中旬开始产卵[12],卵块呈绯红色,易被发现,为了避免天敌的捕食及水中酸性物质对胚囊发育的影响[13]等,雌螺在夜间爬出水面,将卵产于距水面10~50 cm的支撑物上,卵块表面有一层胶膜覆盖,保证其胚胎正常发育[14];卵块产出7~15 d后,幼螺破壳孵出掉落水中,在水中继续完成生长发育过程[15-16]。有研究认为环境是决定卵块分布型的主要因子[13, 17-18]。深水处或低洼积水处的水稻植株上卵块密度较高,在稻田中呈现出中间稀疏、四周稠密的特点[19]。田埂周围杂草较多,一方面给福寿螺提供了丰富的食物,另一方面为其躲避不良环境提供了场所,因此田埂边卵量通常要大于田中卵量[20]。
在对有害昆虫的防治研究中发现,特定的植物对一些害虫产卵具有诱集作用[21]。一些植物浸提液和化学引诱剂含有能吸引害虫发生摄食或交配等活动的物质[22]。此外,对于趋光性害虫,利用灯光、色带等方式,能引诱其在诱集物上大量产卵[23];在林场中适当放置诱木,能有效诱集天牛Cerambycidae在其上产卵,定期将诱木上的卵块去除,能降低天牛的繁殖率从而减少其对林木的危害[24]。有研究发现,福寿螺产卵的高峰期,在稻田中插一些竹片、木条等,也能引诱福寿螺在其上集中产卵,可每2~3 d摘除1次卵块进行销毁[25]。本研究采用不同位置、间距、高度以及不同竹竿形状的插竿方式,研究竹竿吸引福寿螺产卵的分布情况,探索利用竹竿诱集福寿螺产卵,并集中捕获卵块以减少福寿螺孵化繁殖扩散的环境友好型技术。
1. 材料与方法
1.1 材料
2016年3—7月在华南农业大学增城教学科研基地(E113°29′~114°00′,N23°50′~23°37′)稻田中进行试验。种植水稻品种为‘黄华占’,单季种植。在试验区农田及周边的水渠与池塘采集福寿螺。供试竹竿为长60 cm、宽3 cm的竹片和长60 cm、直径1 cm的圆竹条,表面不作其他处理。
1.2 小区前期处理
本研究共有4个试验,同步在不同的小区进行。每个试验共3个重复小区,小区面积为8 m×8 m,各小区间有约35 cm高的水泥田埂。2016年3月1日整田后,在小区出入水口设置拦截网,灌水后投放密达杀螺剂,3月3日再次投放杀螺剂(以灭杀和清除各小区福寿螺),3月5—20日排、灌水同时进行(以排除杀螺剂),21日停止排水,灌水至水深5~8 cm,移栽20日龄水稻,22—26日排、灌水同时进行(以排净杀螺剂),26日停止排水,灌水至水深5~8 cm。3月27日各小区随机投放性成熟的福寿螺雌螺和雄螺各30只,并进行插竿。
1.3 方法
1.3.1 不同插竹竿位置对螺卵的诱集效率
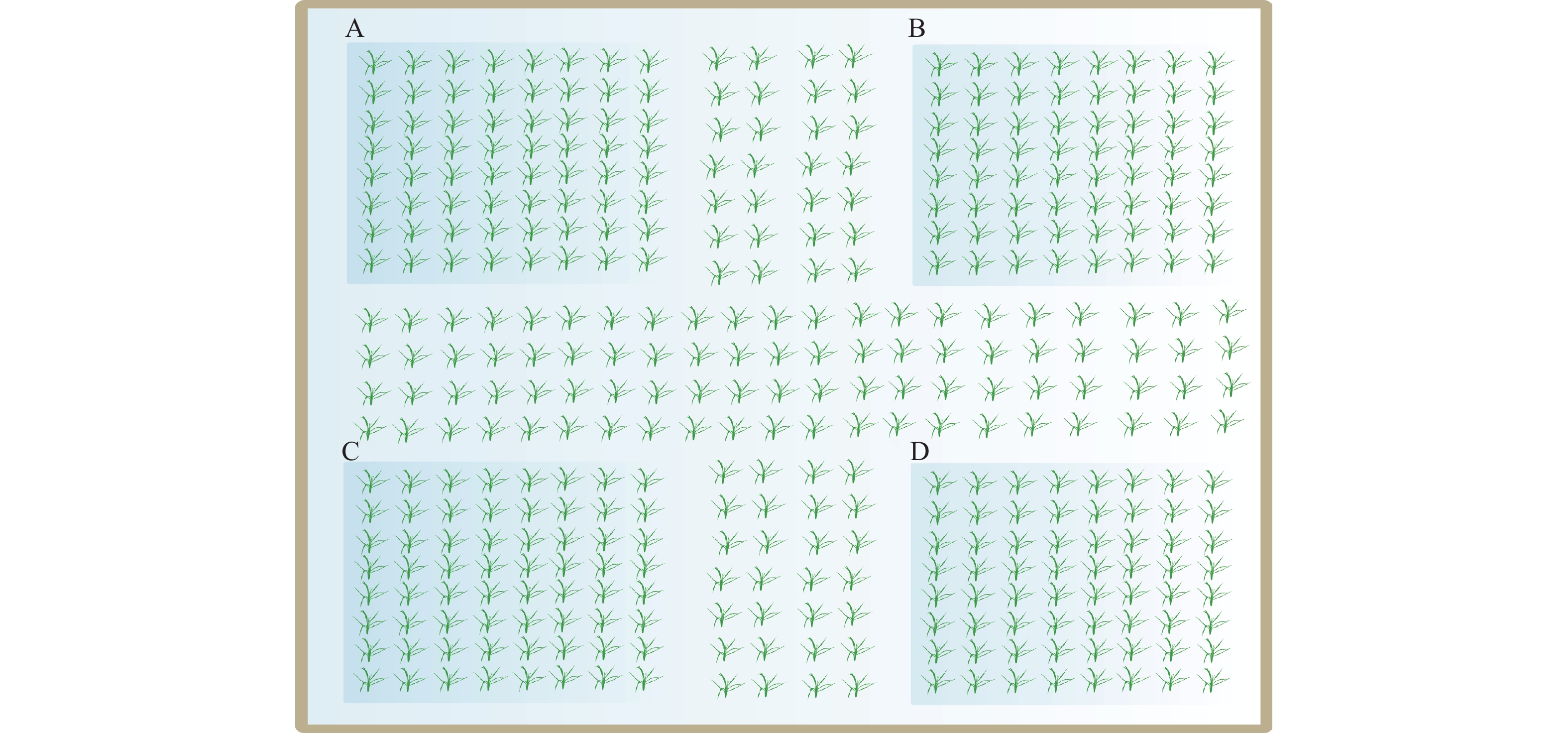
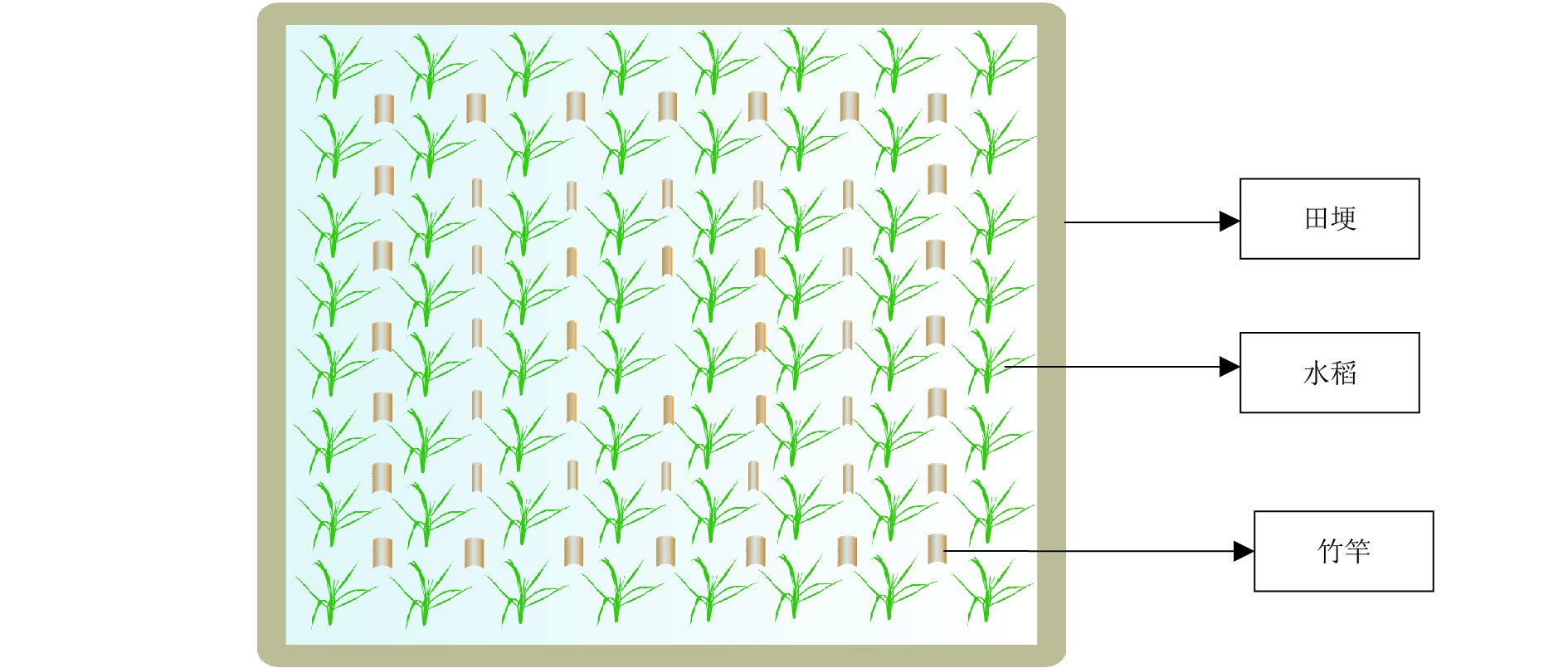
本试验各小区分别在距离田埂1、2、3 m处,插竹竿24根、16根、8根,形成外圈、中圈、内圈3个处理;水稻茎秆为对照(CK),如图1所示。各处理中的插竹竿的顶端距离水面40 cm,间距1 m。分别在4月7日(返青期)、5月1日(拔节期)及5月30日(抽穗期)各进行1次调查,测量水稻株高及茎秆高度,收集每根竹竿上以及水稻茎秆上的卵块(不拔出竹竿,只清除卵块)并计数。
![]() 图 1 稻田中不同位置插竹竿试验小区设计示意图Figure 1. An illustrated test plot with bamboo poles inserted at different positions in paddy field
图 1 稻田中不同位置插竹竿试验小区设计示意图Figure 1. An illustrated test plot with bamboo poles inserted at different positions in paddy field1.3.2 不同插竹竿间距对螺卵的诱集效率
本试验各小区内,不同处理组间有2 m宽不插竹竿只种植水稻的区域,不影响福寿螺自由活动,该区域呈“十”字,将小区划分成“田”字状,分隔出A、B、C、D 4个区域,如图2所示,每个区域面积为3 m×3 m。在区域A间隔30 cm均匀插竹竿32根;区域B间隔70 cm均匀插竹竿16根;区域C间隔100 cm均匀插竹竿8根,为3个不同竹竿间距处理。D区域内水稻茎秆为对照(CK)。各处理中的插竹竿的顶端距离水面40 cm。调查方式、时期与观测指标同“1.3.1”。
1.3.3 不同插竹竿高度对螺卵的诱集效率
前期区域处理同“1.3.2”,在A、B、C区域中各插8根竹竿,竹竿顶端分别距离水面10、30和50 cm;D区域中,水稻茎秆为对照(CK)。各处理中的插竹竿间距为1 m。调查方式、时期与观测指标同“1.3.1”。
1.3.4 不同形状竹竿对螺卵的诱集效率
前期区域处理同“1.3.2”,在A、B区域中,分别插宽3 cm扁竹片、直径1 cm圆竹条各8根为2个不同形状处理。区域C中水稻茎秆为对照(CK);区域D不作使用。各处理中的竹竿顶端距离水面40 cm,间距1 m。调查方式、时期与观测指标同“1.3.1”。
1.4 数据分析
采用SPSS 20.0、Excel 2007软件对试验数据进行整理和统计学分析,对不同位置、间距、离水面高度、不同形状的竹竿处理之间,不同时期之间的卵块数差异采用单因素方差分析,多重比较采用Duncan’s法。绘图使用软件Origin 8.0。
2. 结果与分析
2.1 不同插竹竿位置对螺卵的诱集效率
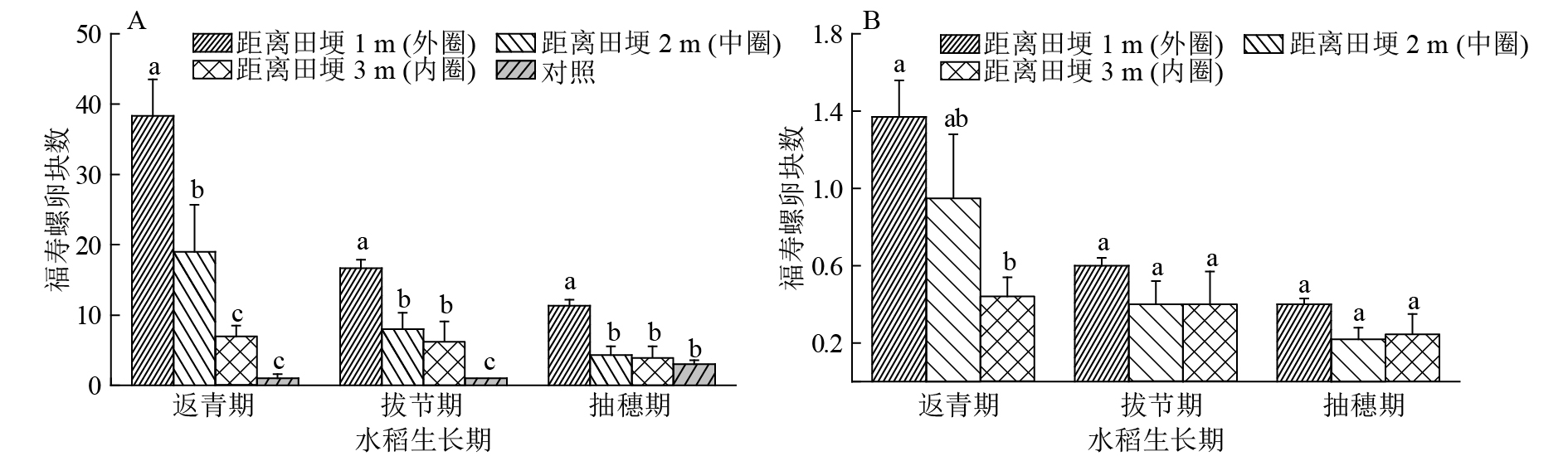
由图3A可见,在水稻返青期,外圈竹竿上收集福寿螺卵块总数为38.33块,中圈为19.00块,内圈为7.00块,对照处理为1.00块,外圈收集得到的卵块数与中圈、内圈、对照处理之间的差异显著(P<0.05)。到拔节期,3个处理收集得到的福寿螺卵块总数有所下降,其中外圈为16.67块,中圈为8.00块,内圈为6.33块,对照为1.00块,外圈与中圈、内圈、对照处理之间的差异显著(P<0.05)。抽穗期时,3个处理的竹竿收集得到的福寿螺卵块数进一步下降,其中外圈收集11.33块,中圈与内圈均为4.00块,对照有所增加,有3.00块,外圈与中圈、内圈、对照差异显著(P<0.05)。以上结果表明,在稻田中不同位置插竹竿均能诱集福寿螺产卵。调查的3个水稻生长时期,外圈竹竿收集福寿螺卵块数显著高于其他圈层及对照的卵块数量(P<0.05),占小区内卵块数比例超过50%;3个处理竹竿上的福寿螺卵块总数呈返青期>拔节期>抽穗期的变化趋势;水稻茎秆上的福寿螺卵块数随水稻生长也有所增加。
![]() 图 3 在水稻不同生育期不同插竹竿位置(A)及其单位面积(1 m2)内(B)所收集的福寿螺卵块情况各图中相同水稻生长期不同柱子上凡是有一个相同小写字母者,表示处理间差异不显著(P>0.05,Duncan’s法)Figure 3. The numbers of egg masses on bamboo poles inserted at different positions (A) and collected per square meter at different positions (B) during rice growth stages
图 3 在水稻不同生育期不同插竹竿位置(A)及其单位面积(1 m2)内(B)所收集的福寿螺卵块情况各图中相同水稻生长期不同柱子上凡是有一个相同小写字母者,表示处理间差异不显著(P>0.05,Duncan’s法)Figure 3. The numbers of egg masses on bamboo poles inserted at different positions (A) and collected per square meter at different positions (B) during rice growth stages试验中,不同处理所占区域面积不同,其中外圈层为28 m2,中圈层为20 m2,内圈层为16 m2。本研究对各处理单位面积(1 m2)内卵块数量差异作了比较,如图3B所示,在水稻返青期,外圈层区域单位面积内竹竿所收集的福寿螺卵块数量为1.37块,中圈层区域单位面积内竹竿所收集的卵块数量为0.95块,内圈层区域单位面积内竹竿收集的卵块0.44块,外圈层与内圈层单位面积收集的卵块数量差异显著(P<0.05),中圈层区域单位面积内所收集的卵块数量与外圈、内圈差异均不显著。在拔节期及抽穗期,卵块数量减少,圈层之间差异不显著。
2.2 不同插竹竿间距对螺卵的诱集效率
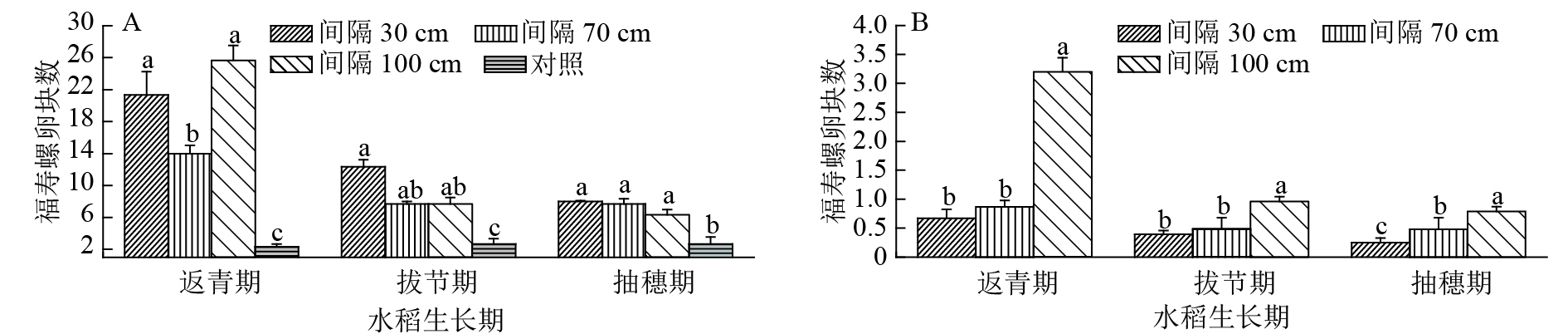
从图4A可知,在水稻返青期时,间距30 cm的竹竿上收集福寿螺卵块总数为21.33块,间距70 cm的卵块总数为14.00块,间距100 cm的为25.67块,间距30 cm与间距100 cm 两种处理显著高于间距70 cm、对照区域的卵块总数(P<0.05),但这2种处理间的卵块总数差异不显著。到拔节期,3个处理收集的福寿螺卵块总数有所下降,间距30、70和100 cm间的差异不显著,与对照差异显著(P<0.05)。抽穗期时,3种间距插竹竿收集到的卵块总数与对照收集到的卵块总数之间差异不显著。以上结果表明,稻田中不同间距的竹竿均能诱集福寿螺在其上产卵。3个时期福寿螺卵块总数总体呈返青期>拔节期>抽穗期的变化趋势,随着水稻的不断生长,竹竿上的卵块总数减少,而对照区域收集的福寿螺卵块总数增加。间距30 cm的竹竿诱集螺卵的效率所占比例最大,但随着水稻的生长,占比下降。间距70 cm所占比例随着水稻的生长进程有所提高,间距100 cm的竹竿上收集的卵块数量占比随水稻的生长有所下降,对照收集的卵块数量占比上升。
![]() 图 4 在水稻不同生育期不同扦插间距的所有竹竿(A)和单竿(B)所收集的福寿螺卵块情况各图中相同水稻生长期柱子上凡是有一个相同小写字母者,表示处理间差异不显著(P>0.05,Duncan’s法)Figure 4. The numbers of egg masses on all bamboo poles (A) and on each bamboo pole (B) with different inserting distance between poles at different rice growth stages
图 4 在水稻不同生育期不同扦插间距的所有竹竿(A)和单竿(B)所收集的福寿螺卵块情况各图中相同水稻生长期柱子上凡是有一个相同小写字母者,表示处理间差异不显著(P>0.05,Duncan’s法)Figure 4. The numbers of egg masses on all bamboo poles (A) and on each bamboo pole (B) with different inserting distance between poles at different rice growth stages试验中,由于各处理的面积相同而插竿间距不同,因此不同处理的竹竿数量有所不同。对单杆上卵块数量差异进行分析,结果如图4B所示,在不同时期不同插竹竿间距的情况下,每种处理的单竿收集福寿螺卵块数有所不同。其中,在返青期时,间距为100 cm的单竿上所收集的福寿螺卵块数为3.21块,间距70 cm处理的单竿所收集福寿螺卵块数为0.88块,间距30 cm处理0.67块。间距为100 cm处理显著高于其他两个处理的单竿卵块数量(P<0.05)。在拔节期和抽穗期,单竿上的卵块数量有所下降,但间距100 cm的卵块数量显著高于其他处理(P<0.05)。
2.3 不同插竹竿高度对螺卵的诱集效率
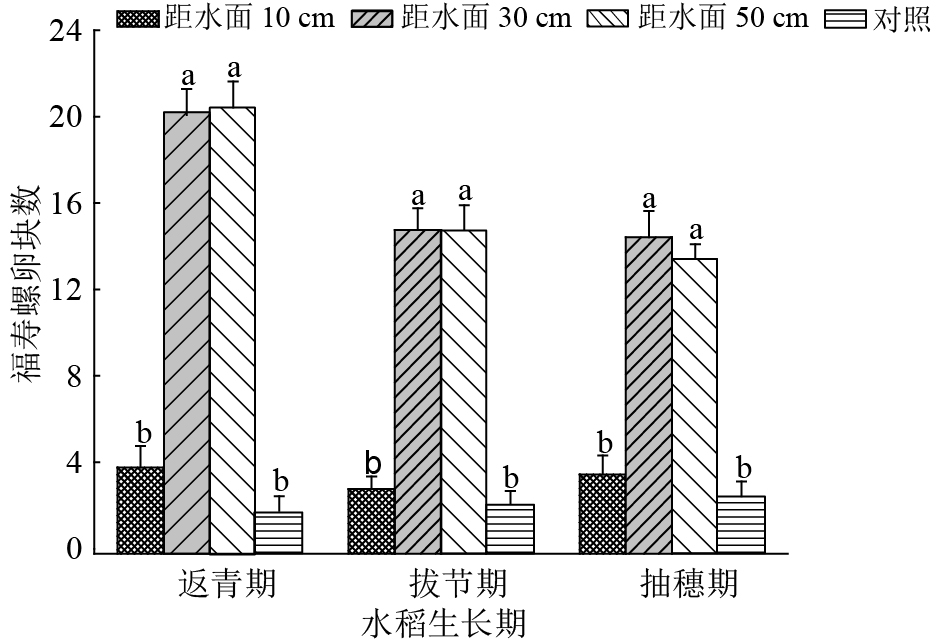
从图5可以看出,不同时期下竹竿顶端距离水面高度不同,竹竿所吸引福寿螺产卵的效率不同。其中返青期时(水稻植株高15~30 cm,水稻茎秆高5~20 cm),距离水面10 cm处理卵块数为4.00块,30 cm处理卵块数为20.33块,距离水面50 cm处理卵块数为20.67块,对照处理卵块数为2.00块。距离水面10 cm处理与距离水面30、50 cm处理的卵块总数均差异显著(P<0.05),与对照组差异不显著。距离水面30、50 cm 处理与对照差异显著(P<0.05)。拔节期时(水稻植株高45~60 cm,水稻茎秆高25~40 cm),距离水面10 cm处理的竹竿上卵块数为3.00块,距离水面30 cm处理竹竿上的卵块数为15.00块,距离水面50 cm处理的卵块数为15.00块,对照区域为2.33块。距离水面10 cm竹竿诱集卵块数量显著低于距离水面30、50 cm竹竿(P<0.05),与对照组差异不显著。距离水面30与50 cm处理间差异不显著;对照与30、50 cm 处理的差异显著(P<0.05)。抽穗期时(水稻植株高75~90 cm,水稻茎秆高45~60 cm),福寿螺卵块总数与拔节期持平,其中距离水面10 cm处理卵块数为3.67块,距离水面30 cm处理卵块数为14.67块,距离水面50 cm处理卵块数为13.67块,对照处理卵块数为2.67块。距离水面10 cm处理与距离水面30、50 cm处理差异显著(P<0.05),与对照组差异不显著。距离水面30、50 cm 处理之间差异不显著。距离水面30、50 cm处理与对照差异显著(P<0.05)。距离水面30、50 cm处理的竹竿对螺卵诱集效率持平,均保持在40%左右,而且随着水稻生长的变化不大。距离水面10 cm高的竹竿与对照占比较低,为5%~10%。
![]() 图 5 水稻各生育期距水面不同高度的竹竿所收集的福寿螺卵块情况图中相同水稻生长期柱子上凡是有一个相同小写字母者,表示处理间差异不显著(P>0.05,Duncan’s法)Figure 5. The numbers of egg masses on bamboo poles inserted with different heights above water surface at different rice growth stages
图 5 水稻各生育期距水面不同高度的竹竿所收集的福寿螺卵块情况图中相同水稻生长期柱子上凡是有一个相同小写字母者,表示处理间差异不显著(P>0.05,Duncan’s法)Figure 5. The numbers of egg masses on bamboo poles inserted with different heights above water surface at different rice growth stages2.4 不同形状竹竿对螺卵的诱集效率
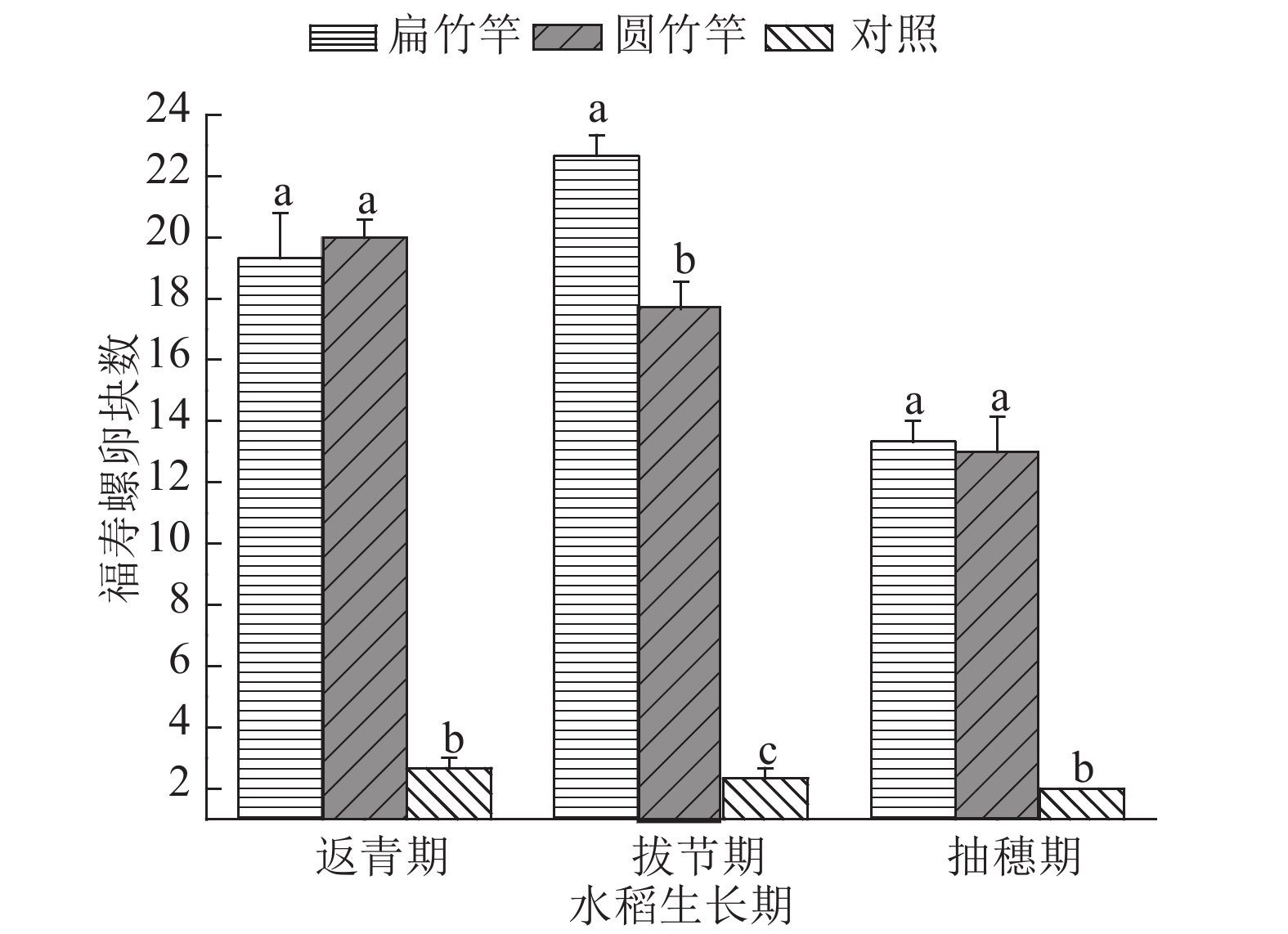
由图6可见,在返青期,扁竹竿上收集到的卵块总数为19.33块,圆竹竿处理所收集的卵块总数为20.00块,对照收集的卵块总数为2.67块,扁竹竿与圆竹竿处理之间差异不显著,但两种处理均与对照差异显著(P<0.05)。在拔节期,扁竹竿上的卵块总数有所增加,为22.67块,圆竹竿有所减少,为17.67块,对照为2.33块,扁竹竿与圆竹竿处理、对照之间差异显著(P<0.05),圆竹竿与对照差异显著(P<0.05)。抽穗期,各种处理所收集卵块总数均减少。其中扁竹竿处理为13.33块,圆竹竿处理为13.00块,对照处理为2.00块,2种竹竿处理之间差异不显著,但均与对照差异显著(P<0.05)。圆竹竿与扁竹竿处理对福寿螺卵的诱集效率持平,为40%~50%,远多于对照的诱集效率(5%~10%),且随着水稻生长变化不大。
![]() 图 6 水稻各生育期不同形状的竹竿所收集到的福寿螺卵块情况图中相同水稻生长期柱子上凡是有一个相同小写字母者,表示处理间差异不显著(P>0.05,Duncan’s法)Figure 6. The numbers of egg masses on bamboo poles with various shapes at different rice growth stages
图 6 水稻各生育期不同形状的竹竿所收集到的福寿螺卵块情况图中相同水稻生长期柱子上凡是有一个相同小写字母者,表示处理间差异不显著(P>0.05,Duncan’s法)Figure 6. The numbers of egg masses on bamboo poles with various shapes at different rice growth stages3. 讨论与结论
3.1 稻田插竹竿诱集福寿螺产卵的总体效果
福寿螺具有沿壁产卵的特性,福寿螺聚集的区域能在田埂边、水渠壁及池塘中露出水面的植物茎秆处发现鲜红色福寿螺卵块,福寿螺喜欢在干燥处产卵。以往的调查中也发现,福寿螺在稻田的分布呈边缘向中心减少的趋势[25],这与福寿螺产卵特性有关。本试验通过调查发现,无论什么形状的竹竿,无论扦插的竹竿处于稻田的何种位置、何种间距、何种高度,在竹竿上均能采集到福寿螺卵块。说明在稻田中插竹竿为福寿螺提供了产卵的环境和载体。
同时,本研究还发现,从返青期到水稻抽穗期、成熟期,在竹竿上收集的福寿螺产卵数量逐渐降低,即随着水稻的生长,竹竿上收集到的福寿螺产卵数量逐渐减少;但在对照区,随着水稻的生长,卵块数量有所增加,这与前人所研究的结论一致,即福寿螺产卵情况在不同水稻生长期有所不同[26]。其主要原因是,在插竹竿区,通过前期竹竿上卵块的移出,稻田中福寿螺繁殖的数量减少,种群规模变小,而在对照区,由于竹竿上无卵块的移出,其福寿螺卵块数并没明显减少。对水稻不同生育期的株高测量结果表明,水稻在返青期株高为15~30 cm,可供产卵的茎秆高度为5~20 cm,此时期水稻植株矮,而福寿螺喜欢在高的附着物上产卵,此外,水稻稀疏,福寿螺容易找到竹竿,因此福寿螺选择附着在竹竿上产卵。到了拔节期、成熟期,水稻植株高度增为60~90 cm,茎秆高度为40~70 cm,茎秆变粗,分蘖增多导致植株茂盛,福寿螺不容易找到竹竿,因此福寿螺附着在茎秆产卵的数量增加。
上述的结果表明,扦插竹竿上收集的福寿螺卵块数量大体随生育期的进程而呈下降趋势,其中返青期竹竿上收集的卵块数量最多,是拔节期的2倍多,与其他时期相比差异显著。因此,在水稻移栽后即进行插竿,可最大限度地诱集福寿螺在其上产卵,可集中收集捣毁卵块或再利用,从而实现通过物理手段有效防除福寿螺的目的。
3.2 不同插竹竿方式对福寿螺产卵的影响
本研究证实福寿螺喜欢将卵块产在竹竿上,但竹竿在稻田中扦插的位置不同,收集到的福寿螺卵块数量也有较大差别。外圈(靠近田埂)的竹竿所占区域面积较大,所收集的总卵块数较多,在进一步分析中发现其单位面积所收集的卵块也较多,在整个小区收集的总卵块数量中所占比重达到50%以上,这说明外圈不只是范围大竿数多而收集的卵块较多,在位置上也对吸引福寿螺产卵有优势。这与前人得出的福寿螺田埂到田中心数量由多到少的分布规律与沿壁产卵习性结论是一致的。水稻田中福寿螺随水流移动,稻田灌溉水入、出口处一般聚集较多的螺量,在田间调查时,螺数量从稻田埂边开始向稻田中部减少,卵数量从田埂上的杂草茎秆等附着物向稻田中部的水稻茎秆减少[25]。因此,在实际运用中,可将竹竿插在靠近田埂的位置,能达到诱集福寿螺产卵的最佳效果。
已有研究表明,成年福寿螺种群间喜欢聚集,个体间相互吸引,但其产卵受到干扰时会中断,因而不喜欢集中产卵[6],从本研究中的竹竿间距试验结果来看,在水稻返青期后,扦插的竹竿越多、越密,能收集到福寿螺卵块越多,但单竿上卵块数量越少;竹竿数量越少、距离越宽,福寿螺卵块总数越低,但单竿上数量越高。这说明了福寿螺在竹竿上产卵总量受竹竿数量的影响大,而单竿卵块数量受竹竿的间距影响更大。这些结果表明,在实际应用中,若要收集更多卵块,达到更佳防治效果,且田块面积不大时,可减小竹竿间距至30 cm,加大竹竿扦插密度;若要节省劳动力,且田块范围较大,则可以适当加大竹竿间距至100 cm,进而减少竹竿的数量,但仍需保证一定数量的竹竿以收集更多的卵块。
前人研究结果表明,福寿螺产卵高度与水面距岸壁的高度有关联[26],在自然条件下,由于福寿螺卵在水中不能正常发育,为了更好地繁衍后代,福寿螺总是将卵产在距水面一定高度的岸壁或者其他物体上[27],在露出水面10~80 cm的物体上均能发现福寿螺的卵块,其中露出水面30~50 cm高的物体上的卵块数量最多[28]。在本研究中,我们设计了离稻田水面10、30和50 cm的竹竿这3个处理,结果表明,距离水面30和50 cm的竹竿上收集到的福寿螺卵块数量最多,距离水面10 cm的处理收集的卵块数最少,与对照一样,几乎没有发现卵块,这与前人研究得到的福寿螺产卵习性一致[28]。此外,水稻株高在返青期为15~30 cm,茎秆高度为5~20 cm,与10 cm处理高度相近,卵块数量也与之相近;拔节期时水稻株高为45~60 cm,水稻茎秆高度为25~40 cm,与30和50 cm处理高度相近,但此时水稻茎秆上卵块数量显著低于竹竿上卵块数量;到了抽穗期时,水稻株高为75~90 cm,水稻茎秆高度为45~60 cm,高于竹竿各处理的高度,但水稻茎秆上的卵块数量仍显著低于竹竿上卵块数量,这说明无论水稻茎秆高度如何,福寿螺均选择附着在距离水面30~50 cm竹竿上产卵,这可能是由于水稻茎秆上的分蘖导致其表面凹凸不平,且成熟期的叶片表面有细小的毛刺,影响雌螺的爬行及卵块的附着,因而福寿螺会选择附着在表面更平整的竹竿上产卵。因此,在实际运用中,插入距离水面30~50 cm的竹竿防治效果最佳。另外,本研究还发现,在扦插圆竹条或扁竹竿上所收集到的卵块数量在返青期和抽穗期无显著差异,但在拔节期达到显著差异,说明竹竿的不同形状(扁圆程度)对诱集福寿螺产卵存在一定影响,这可能与扁竹竿更利于福寿螺附着和固定其卵块有关。
3.3 结论
本试验研究结果综合表明,在稻田中扦插竹竿能够有效吸引福寿螺产卵。水稻移栽后,在靠近田埂1 m处,距离水面30~50 cm,且竹竿间距100 cm时,沿田埂内周扦插一圈扁竹竿(若在螺卵发生密集的小范围区域,可减小间距为30 cm,高密度扦插竹竿),能更大限度地吸引福寿螺产卵,即对福寿螺卵诱集效率更高。由于福寿螺繁殖能力极强,单只雌螺1次能产卵50~500粒,并且有80%的孵化成功率,孵化期仅需10~15 d,繁殖力惊人[29-30],因此,根据本试验结果,在福寿螺繁殖盛期,通过适当的插竹竿方式,可很好地收集福寿螺卵块,并统一销毁处理,从而大大降低福寿螺繁殖的速率,以达到减少螺害的目的。采用这样的物理方式防控福寿螺,成本低、简单易行、见效快,属于一种环境友好型技术,值得推广应用。
-
![]()
图 1 稻田中不同位置插竹竿试验小区设计示意图
Figure 1. An illustrated test plot with bamboo poles inserted at different positions in paddy field
![]()
图 3 在水稻不同生育期不同插竹竿位置(A)及其单位面积(1 m2)内(B)所收集的福寿螺卵块情况
各图中相同水稻生长期不同柱子上凡是有一个相同小写字母者,表示处理间差异不显著(P>0.05,Duncan’s法)
Figure 3. The numbers of egg masses on bamboo poles inserted at different positions (A) and collected per square meter at different positions (B) during rice growth stages
![]()
图 4 在水稻不同生育期不同扦插间距的所有竹竿(A)和单竿(B)所收集的福寿螺卵块情况
各图中相同水稻生长期柱子上凡是有一个相同小写字母者,表示处理间差异不显著(P>0.05,Duncan’s法)
Figure 4. The numbers of egg masses on all bamboo poles (A) and on each bamboo pole (B) with different inserting distance between poles at different rice growth stages
![]()
图 5 水稻各生育期距水面不同高度的竹竿所收集的福寿螺卵块情况
图中相同水稻生长期柱子上凡是有一个相同小写字母者,表示处理间差异不显著(P>0.05,Duncan’s法)
Figure 5. The numbers of egg masses on bamboo poles inserted with different heights above water surface at different rice growth stages
-
[1] HALWART M. The golden apple snail Pomacea canaliculata in Asian rice farming systems: Present impact and future threat[J]. Int J Pest Manage, 1994, 40(2): 199-206.
[2] YOSHIE H, YUSA Y. Effects of predation on the exotic freshwater snail Pomacea canaliculata (Caenogastropoda: Ampullariidae) by the indigenous turtle Chinemys reevesii(Testudines: Geoemydidae)[J]. Appl Entomol Zool, 2008, 43(4): 475-482.
[3] YUSA Y, WADA T. Impact of the introduction of apple snails and their control in Japan[J]. Naga: The IC LARM Quarterly, 1999, 22(3): 9-13.
[4] 潘长旺, 凌洪博, 梁韶晖, 等. 温州地区广州管圆线虫病的临床及流行病学研究[J]. 中华医学杂志, 2000, 80(3): 204-205. [5] LOWE S, BROWNE M, BOUDJELAS S, et al. 100 of the world’s worst invasive alien species: A selection from the global invasive species database[M]. Auckland, New Zealand: Invasive Species Specialist Group. 2000.
[6] 叶建人, 李云明. 福寿螺在水稻田的消长规律及其田间分布动态[J]. 中国农学通报, 2009, 25(3): 185-188. [7] 俞小平, 和田节, 顾中方, 等. 稻田福寿螺的发生和治理[J]. 浙江农业学报, 2001, 13(5): 247-252. [8] 王志高, 谭继才, 刘军, 等. 福寿螺综合防治研究进展[J]. 中国农学通报, 2009, 25(12): 201-205. [9] CHENG E, KAO C. Control of golden apple snail, Pomacea canaliculata (Lamarck), in Taiwan[J]. Phil Rice, 2006: 155-167.
[10] 陶红群, 贾滨洋, 张峰瑜, 等. 成都市境内外来物种福寿螺的危害现状调查及防治措施分析[J]. 四川环境, 2005, 24(3): 108-110. [11] ITO K. Environmental factors influencing over wintering success of the golden apple snail, Pomacea canaliculata (Gastropoda: Ampullariidae), in the northernmost population of Japan[J]. Appl Entomol Zool, 2002, 37(4): 655-661.
[12] 周外, 刀学琼, 邓才明, 等. 福寿螺的繁殖与生长研究[J]. 安徽农业科学, 2015, 43(11): 134-135. [13] 段清星, 李湘, 王本君, 等. 福寿螺密度变化的环境影响因素研究[J]. 四川动物, 2016, 35(4): 503-510. [14] 董朝莉. 福寿螺的生物生态学特性及在广西的分布与危害现状研究[D]. 桂林: 广西师范大学, 2006. [15] WANG Z, TAN J, TAN L, et al. Control the egg hatchling process of Pomacea canaliculata (Lamarck) by water spraying and submersion[J]. Acta Ecologica Sinica, 2012, 32(4): 184-188.
[16] FRASSA M V, CEOLIN M, DREON M S, et al. Structure and stability of the neurotoxin PV2 from the eggs of the apple snail Pomacea canaliculata[J]. Biochim Biophys Acta, 2010, 1804(7): 1492-1499.
[17] SEUFFERT M E, MARTÍN P R. Distribution of the apple snail Pomacea canaliculata in Pampean streams (Argentina) at different spatial scales[J]. Limnologica, 2013, 43(2): 91-99.
[18] 朱丽霞, 黄瑶瑶, 张泽宏, 等. pH, 食物和光周期对福寿螺生长发育和繁殖的影响[J]. 生态学报, 2015, 35(8): 2644-2651. [19] SEUFFERT M. A lentic dweller in lotic habitats: The behavior of the invasive South American apple snail Pomacea canaliculata in flowing water[J]. Aquat Ecol, 2012, 46(1): 129-142.
[20] HARA A, HAMASAKI K, YOSHIDA K, et al. Canal type affects invasiveness of the apple snail Pomacea canaliculata through its effects on animal species richness and waterweed invasion[J]. Biol Invasions, 2015, 17(1): 63-71.
[21] 鲁艳辉, 高广春, 郑许松, 等. 诱集植物香根草对二化螟幼虫致死的作用机制[J]. 中国农业科学, 2017, 50(3): 486-495. [22] 李凯利, 高景鹏, 曹杰, 等. 3种植物浸液对白纹伊蚊产卵的影响研究[J]. 中国媒介生物学及控制杂志, 2017, 28(1): 4-8. [23] 王文龙, 任利利, 张连生, 等. 不同波长LED灯对油松毛虫的诱捕效果与评价[J]. 应用昆虫学报, 2017, 54(6): 955-960. [24] 陈水呇. 松诱木设置对松墨天牛诱集效果的影响[J]. 青海农林科技, 2016, 2: 30-32. [25] 毛琦, 朱豪红, 党绍东, 等. 稻田福寿螺空间分布型和抽样技术的研究[J]. 广西植保, 2011, 24(1): 11-15. [26] 贤振华, 覃荣乐, 冷付春, 等. 稻田福寿螺为害程度上升原因分析及防控策略[J]. 广西农业科学, 2009, 40(8): 1007-1009. [27] ESTEBENET A L, MARTIN P R. Pomacea canaliculata (Gastropoda: Ampullaridae): Life- history traits and their plasticity[J]. Biocell, 2002, 26(1): 83-89.
[28] 杨爱娟, 谭济才, 袁哲明, 等. 福寿螺产卵高度与克氏原螯虾捕食卵块的研究[J]. 中国农学通报, 2012, 28(8): 90-93. [29] JOSHI R C, VREYSEN M J B, ROBINSON A S, et al. Problems with the management of the golden apple snail Pomacea canaliculata: An important exotic pest of rice in Asia[M]//RAVINDRA C J. Area-wide control of insect pests: From research to field implementation. Berlin: Springer Netherlands, 2007: 257-264.
[30] 刘军, 何跃进, 谭济才, 等. 自然条件下福寿螺繁殖特性[J]. 应用生态学报, 2012, 23(2): 559-565.
 下载:
下载:
计量
- 文章访问数: 1803
- HTML全文浏览量: 3
- PDF下载量: 1656
