
Effects of Fusarium oxysporum on the biological characteristics and expression transcriptome of Caenorhabditis elegans
-
摘要:目的
探讨尖孢镰刀菌Fusarium oxvsporum对秀丽隐杆线虫Caenorhabditis elegans生物学特征的影响,明确线虫响应尖孢镰刀菌侵染的关键通路。
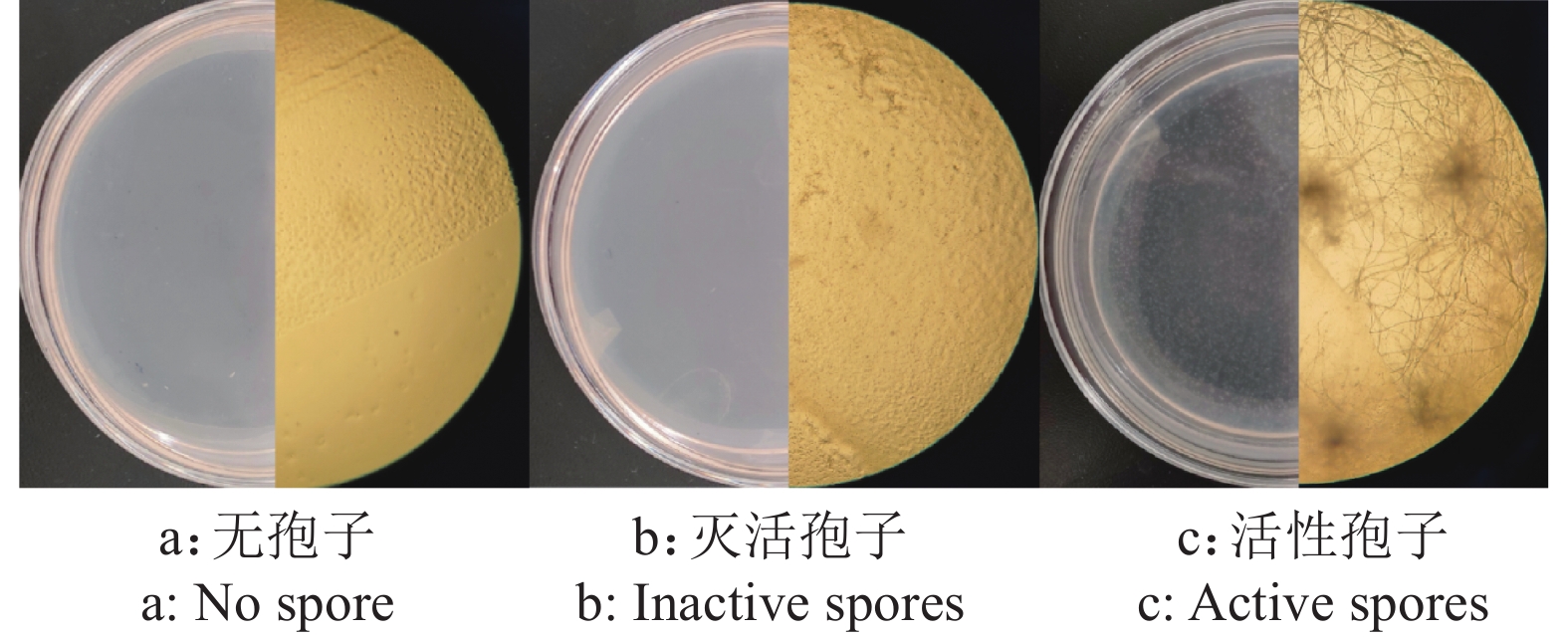
方法将尖孢镰刀菌活性孢子和灭活孢子分别与秀丽隐杆线虫共培养,测定C. elegans寿命、体长、运动能力、繁殖力以及咽泵运动速率等基本生物学特征;进行全基因组测序,通过GO和KEGG分析明确线虫响应尖孢镰刀菌侵染的关键通路和生物学功能。
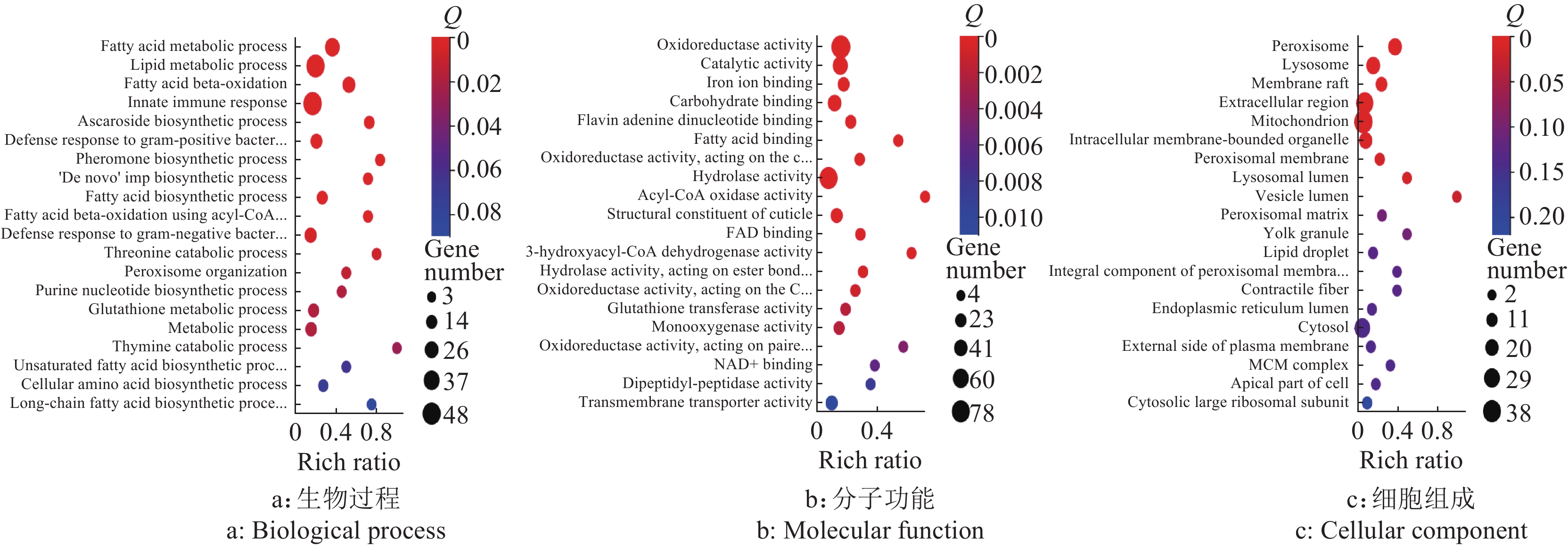
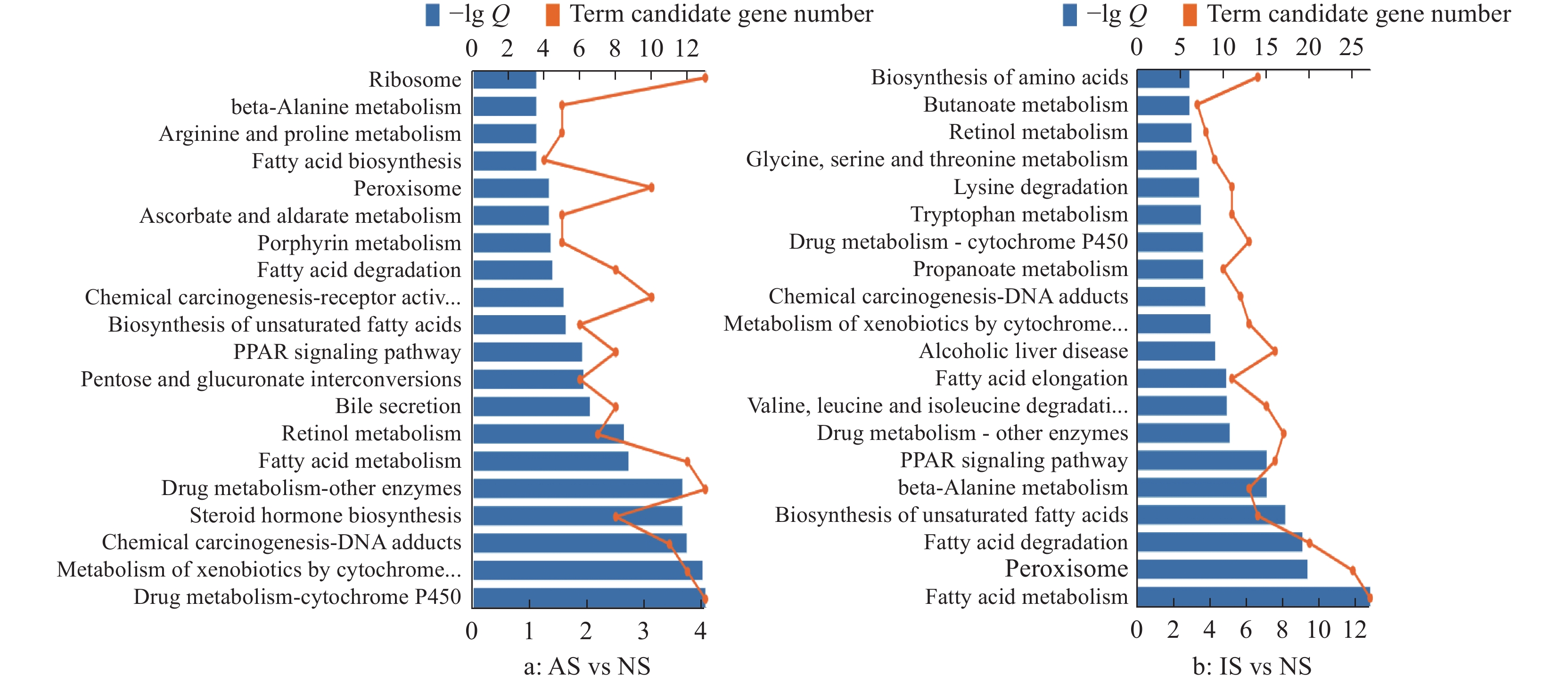
结果尖孢镰刀菌活性分生孢子与线虫共培养可导致线虫寿命下降,虫体显著缩短,线虫繁殖力显著降低;但共培养并未对线虫的咽泵运动速率及运动能力产生明显影响。GO分析发现,差异表达基因富集在先天免疫反应和氧化还原酶活性等生物学功能方面;KEGG分析发现,差异表达基因富集在药物代谢——细胞色素P450、代谢外源性物质的细胞色素P450和脂肪酸代谢等通路。
结论尖孢镰刀菌对秀丽隐杆线虫的基本生命活动有不利影响,这为深入探讨线虫响应尖孢镰刀菌的天然免疫机制及与植物免疫的相似性,从而寻找植物相关抗病基因奠定了基础。
Abstract:ObjectiveTo investigate how Fusarium oxysporum affects the fundamental biological traits of nematodes (Caenorhabditis elegans), and identify the critical mechanism of nematode responding to F. oxysporum infection.
MethodThe active and inactive spores were cocultured with C. elegans to determine the biological characteristics of C. elegans, such as lifespan, body length, motility/reproductive ability and pharyngeal pumping rate. The whole genome sequencing was performed to find the key pathways and biological functions of nematode responding to F. oxysporum infection by GO and KEGG analysis.
ResultThe coculture with active spores shortened the longevity of nematodes, significantly decreased the body size and reduced the reproductive ability of nematodes. However, the coculture did not obviously affect the pharyngeal pumping rate and motility ability of nematodes. GO analysis showed that the differentially expressed genes were enriched in biological functions such as innate immune response and oxidoreductase activity. KEGG analysis showed that the differentially expressed genes were enriched in cytochrome P450 metabolizing drug, cytochrome P450 metabolizing xenobiotics, fatty acid metabolism and other pathways.
ConclusionF. oxysporum has adverse effects on the basic life activities of C. elegans, which provides insight into the natural immune mechanism of nematodes in response to F. oxysporum and the similarity with plant immunity, so as to find plant-related disease resistance genes.
-
-
![]()
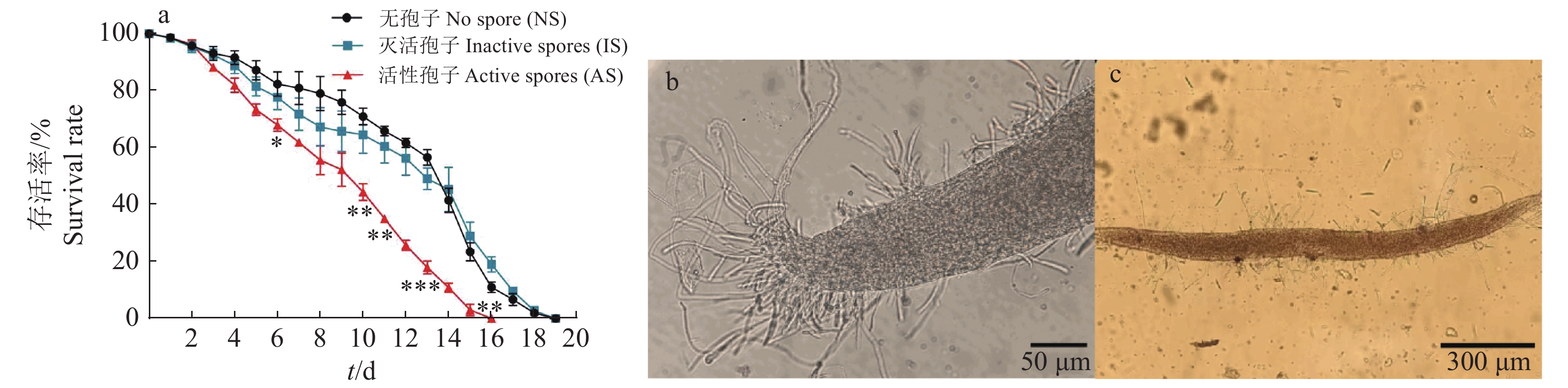
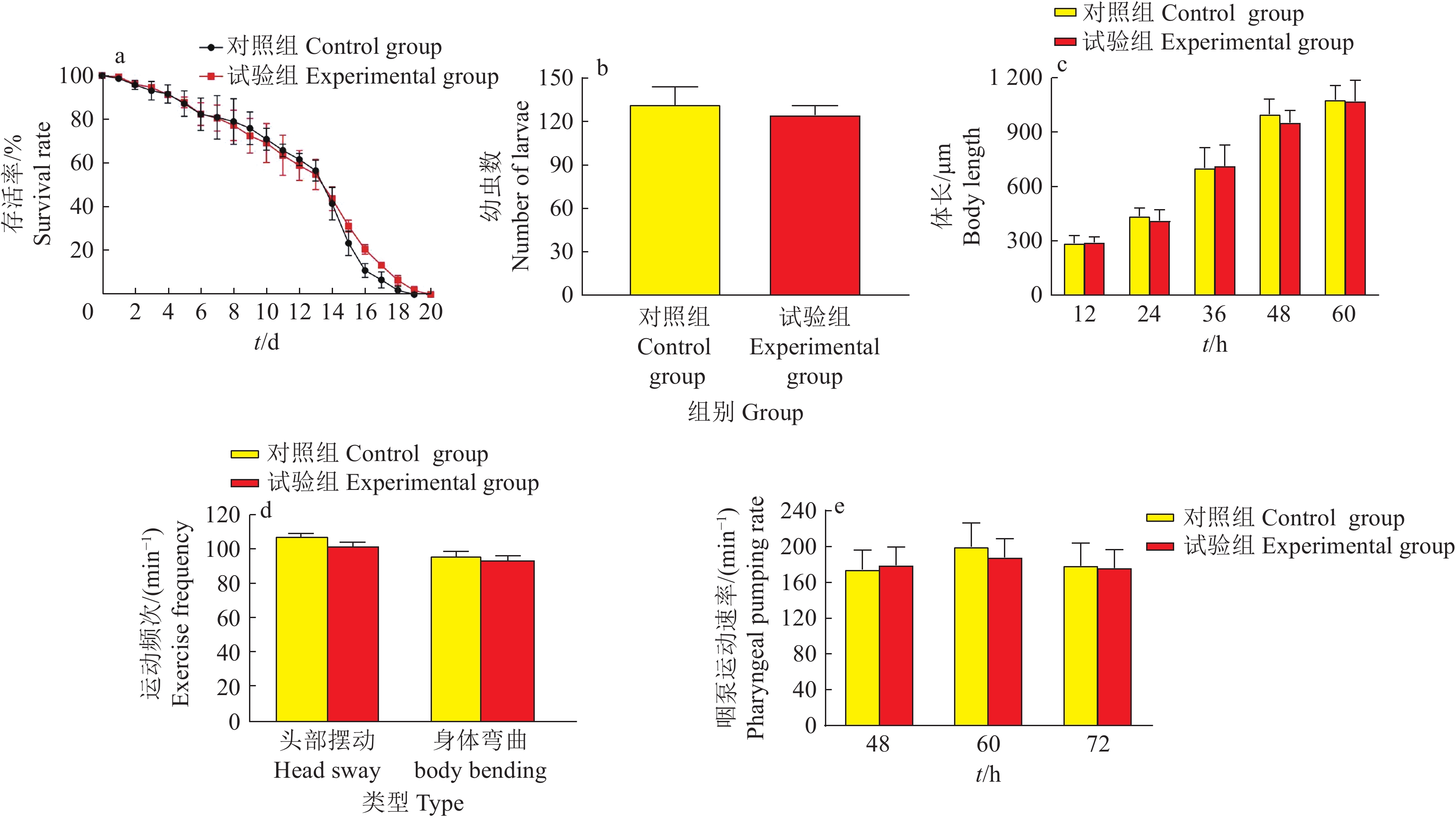
图 2 尖孢镰刀菌对线虫存活率的影响
“*”“**”“***”分别表示相同时间处理与对照(NS)在0.05、0.01和0.001水平差异显著(t检验)
Figure 2. Effect of Fusarium oxysporum on Caenorhabditis elegans survival rate
“*” “**” and “***” indicate that there are significant differences between treatment groups and the control group (NS) at 0.05, 0.01 and 0.001 levels at the same time, respectively (t test)
![]()
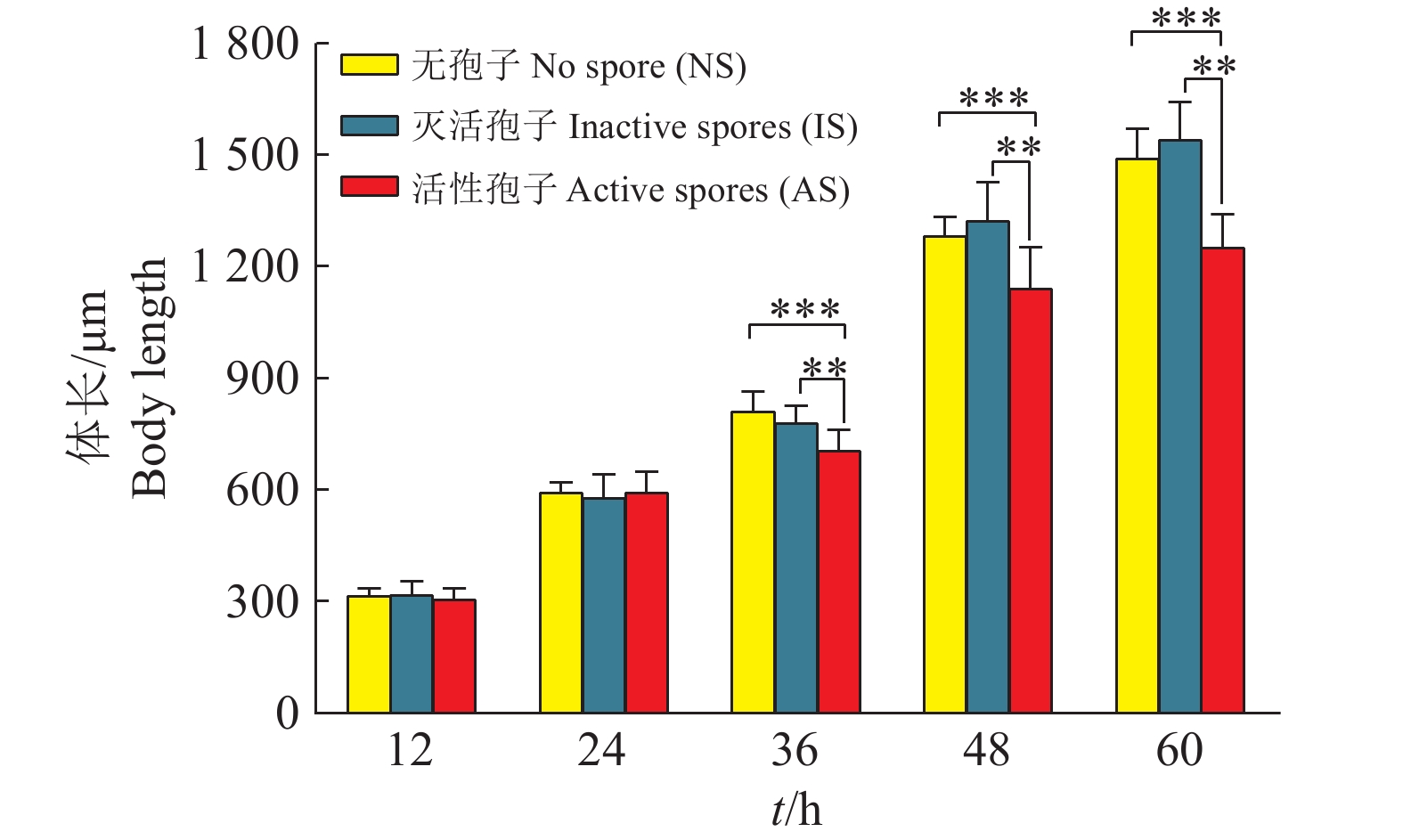
图 3 尖孢镰刀菌对线虫体长的影响
“**”和“***”分别表示相同时间内不同处理组间在0.01和0.001水平差异显著(t检验)
Figure 3. Effect of Fusarium oxysporum on nematode body length
“**” and “***” indicate that there are significant differences among different treatment groups at 0.01 and 0.001 levels at the same time, respectively (t test)
![]()
图 4 线虫分别在无孢子(NS)、灭活孢子(IS)及活性孢子(AS)组共培养不同时间的体长
Figure 4. Body length of nematodes in NS, IS and AS groups at different time of cocultrue
![]()
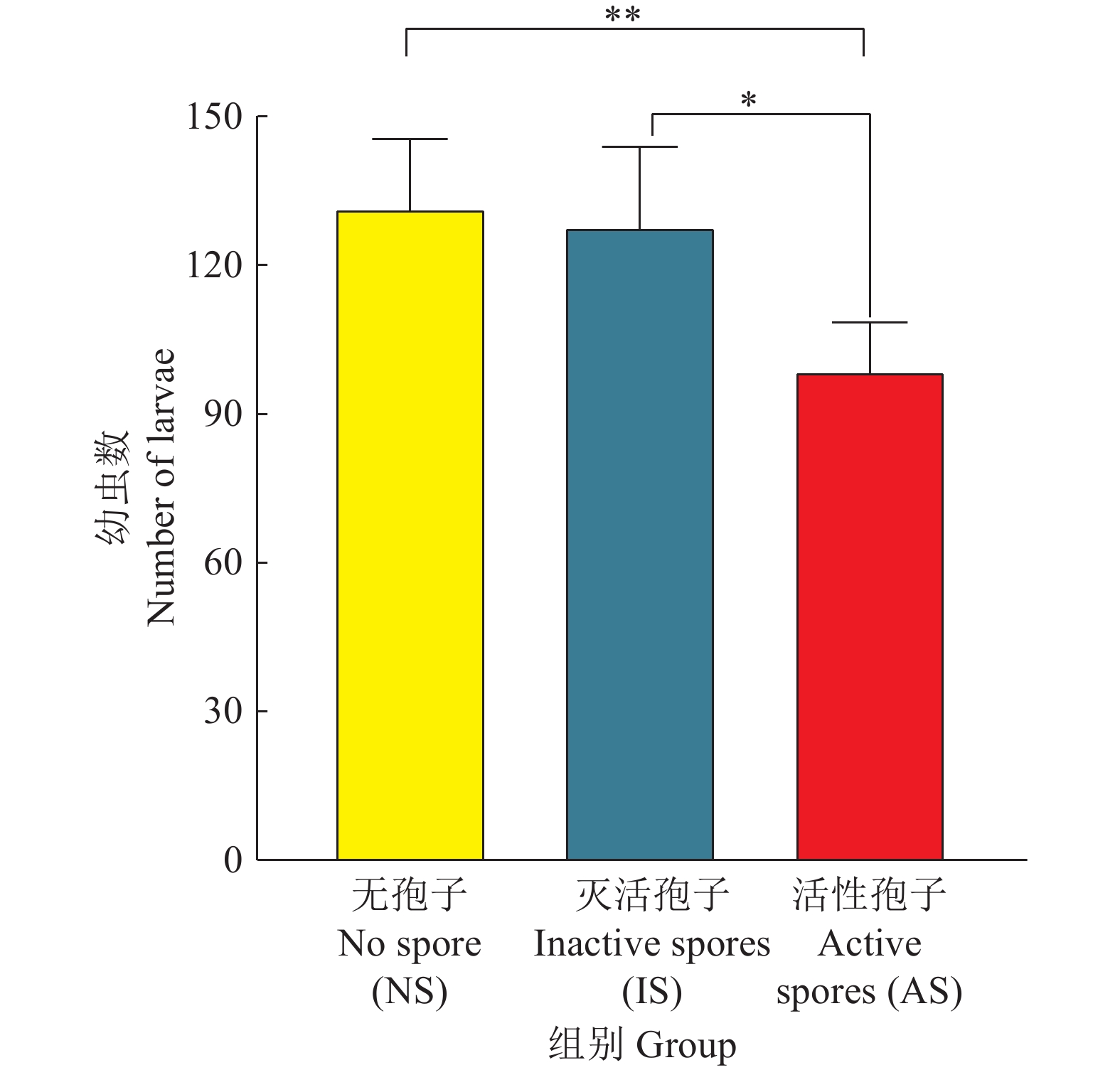
图 5 尖孢镰刀菌对线虫繁殖的影响
“*”和“**”分别表示相同时间内不同处理组间在0.05和0.01水平差异显著(t检验)
Figure 5. Effects of Fusarium oxysporum on fecundity in Caenorhabditis elegans
“*” and “**” indicate that there are significant differences among different treatment groups at 0.05 and 0.01 levels at the same time, respectively (t test)
![]()
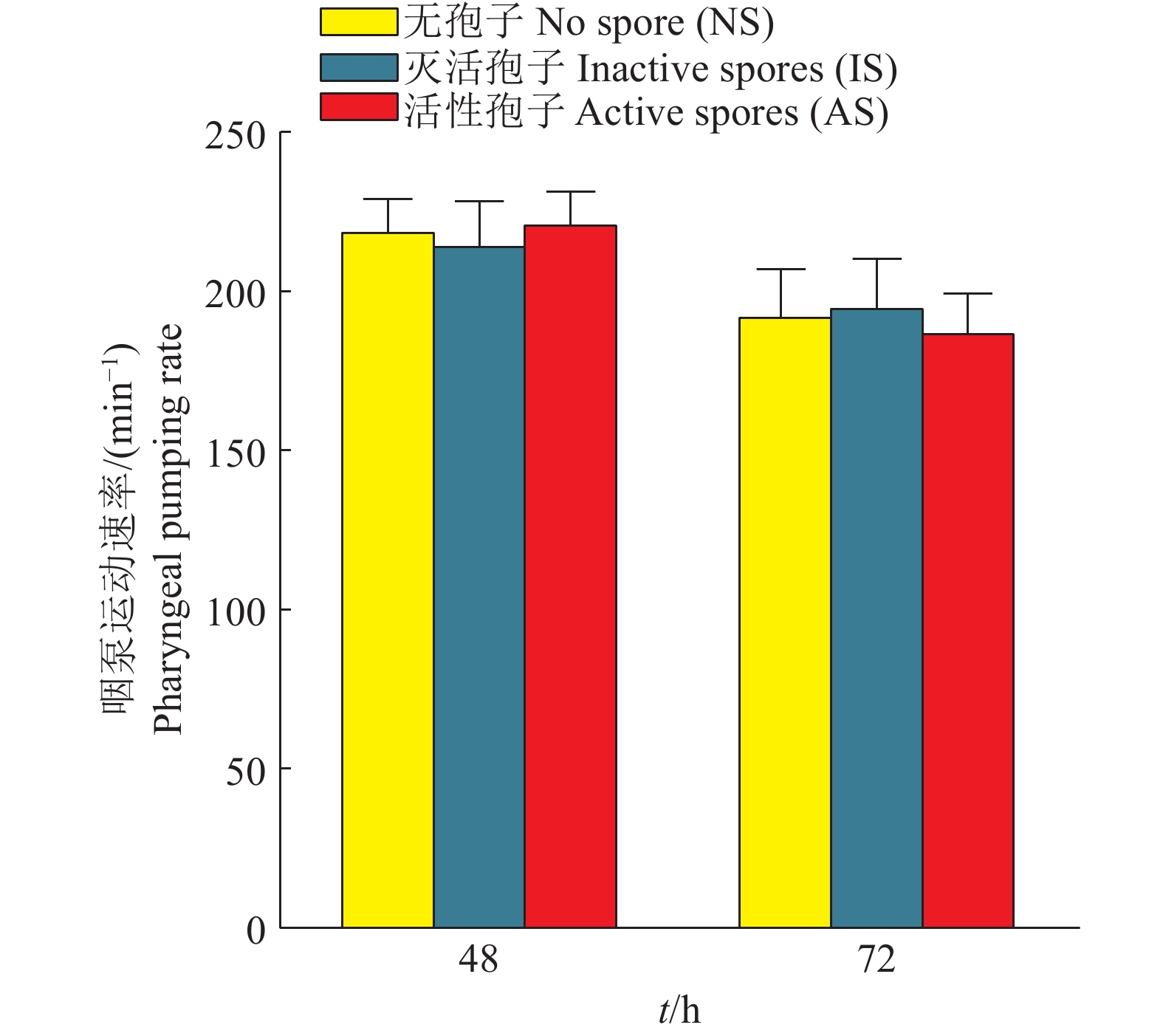
图 6 尖孢镰刀菌对线虫咽泵运动速率的影响
Figure 6. Effect of Fusarium oxysporum on pharyngeal pumping rate of nematodes
![]()
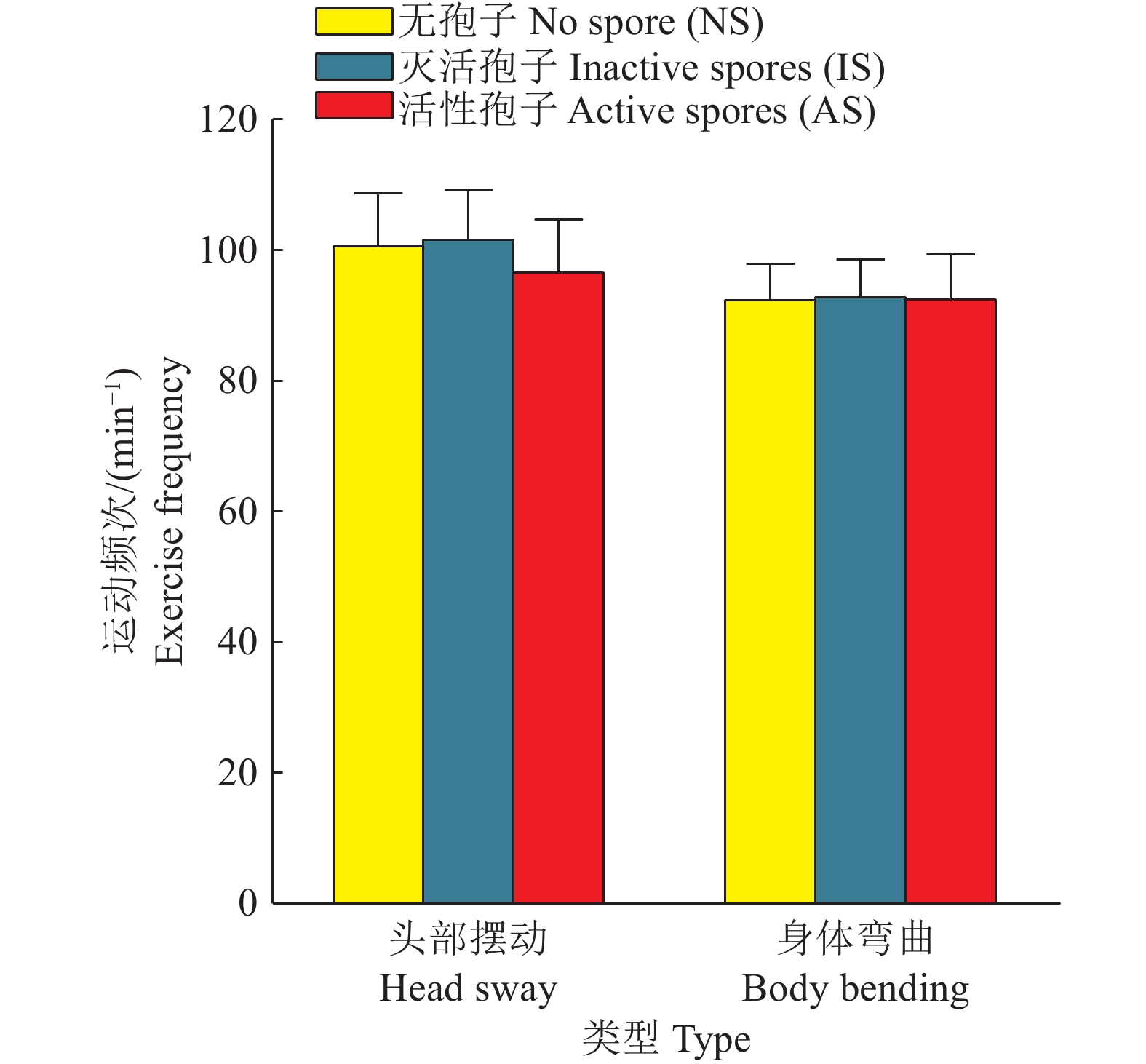
图 7 尖孢镰刀菌对线虫运动能力的影响
Figure 7. Effects of Fusarium oxysporum on locomotor ability of nematodes
![]()
图 8 尖孢镰刀菌毒素对秀丽隐杆线虫生理特征的影响
Figure 8. Effects of Fusarium oxysporum toxin on the physiological characteristics of Caenorhabditis elegans
![]()
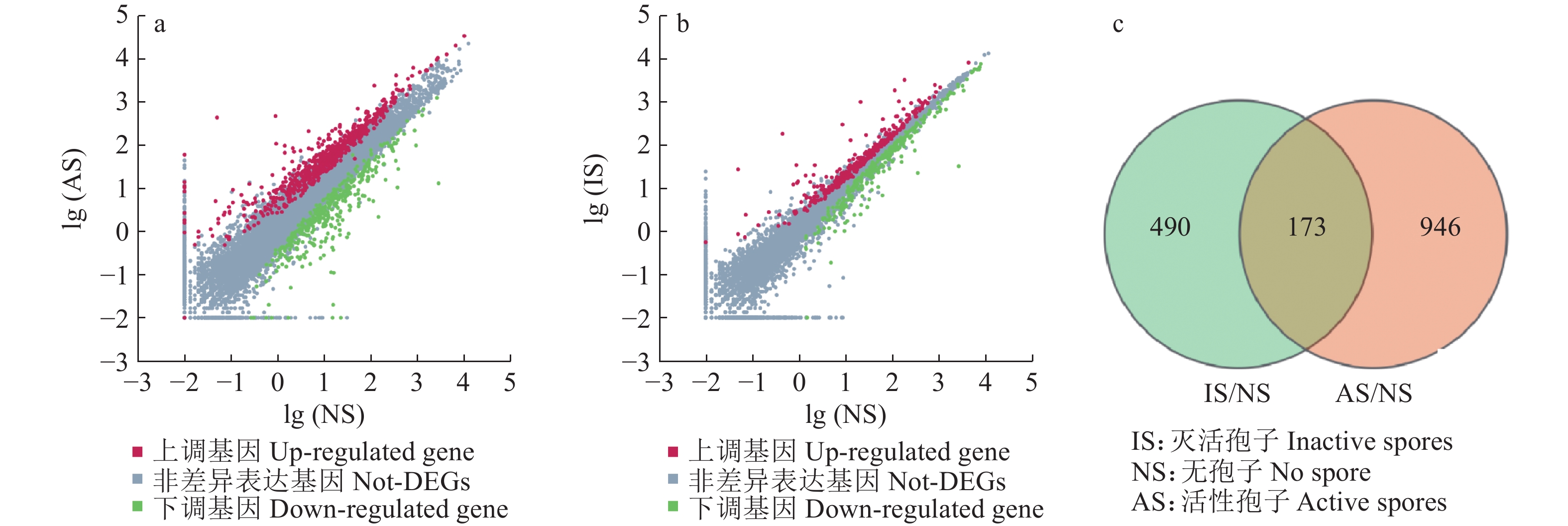
图 9 差异基因散点图(a、b)和韦恩图(c)
Figure 9. Differential gene scatter plot (a, b) and Venn diagram (c)
![]()
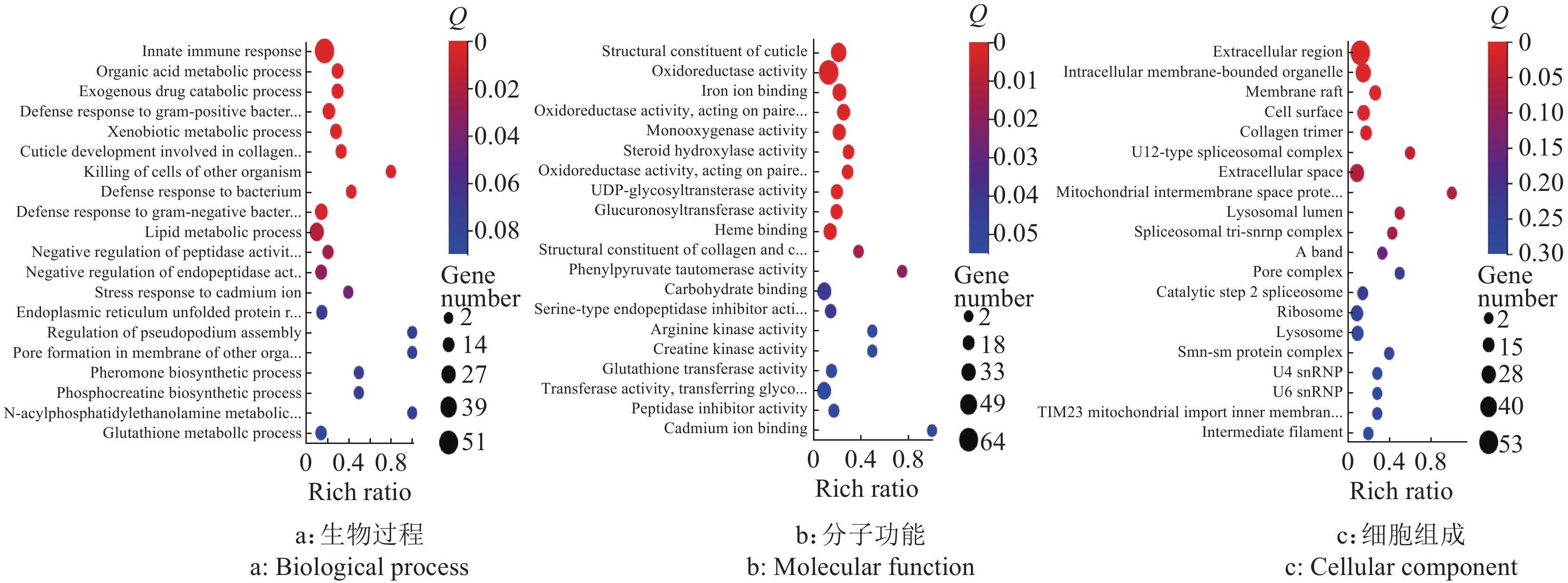
图 10 活性孢子 (Active spores, AS)组和无孢子 (No spore, NS)组差异基因的GO功能注释
Figure 10. GO function annotation of differential genes in AS and NS groups
![]()
图 11 灭活孢子(Inactive spores, IS)组和无孢子(No spore, NS)组差异基因的GO功能注释
Figure 11. GO function annotation of differential genes in IS and NS groups
-
[1] POULIN R, KEENEY D B. Host specificity under molecular and experimental scrutiny[J]. Trends in Parasitology, 2008, 24(1): 24-28. doi: 10.1016/j.pt.2007.10.002
[2] SCHULZE-LEFERT P, PANSTRUGA R. A molecular evolutionary concept connecting nonhost resistance, pathogen host range, and pathogen speciation[J]. Trends in Plant Science, 2011, 16(3): 117-125. doi: 10.1016/j.tplants.2011.01.001
[3] THINES M. An evolutionary framework for host shifts - jumping ships for survival[J]. New Phytologist, 2019, 224(2): 605-617. doi: 10.1111/nph.16092
[4] JIANPING X. Assessing global fungal threats to humans[J]. mLife, 2022, 1(3): 223-240. doi: 10.1002/mlf2.12036
[5] FIGTREE M, WEEKS K, CHAN L, et al. Colletotrichum gloeosporioides sensu lato causing deep soft tissue mycosis following a penetrating injury[J]. Medical Mycology Case Reports, 2013, 2: 40-43. doi: 10.1016/j.mmcr.2013.01.003
[6] 杜宾. 镰刀菌的分类和研究现状[J]. 太原学院学报(自然科学版), 2017, 35(4): 68-70. [7] 叶旭红, 林先贵, 王一明. 尖孢镰刀菌致病相关因子及其分子生物学研究进展[J]. 应用与环境生物学报, 2011, 17(5): 759-762. [8] NÜRNBERGER T, BRUNNER F, KEMMERLING B, et al. Innate immunity in plants and animals: Striking similarities and obvious differences[J]. Immunological Reviews, 2004, 198(1): 249-266. doi: 10.1111/j.0105-2896.2004.0119.x
[9] DARDICK C, RONALD P. Plant and animal pathogen recognition receptors signal through non-RD kinases[J]. PLoS Pathogens, 2006, 2(1): e2. doi: 10.1371/journal.ppat.0020002.
[10] AUSUBEL F M. Are innate immune signaling pathways in plants and animals conserved?[J]. Nature Immunology, 2005, 6(10): 973-979. doi: 10.1038/ni1253
[11] 郭军英. 关于血球计数板的使用及注意事项[J]. 教学仪器与实验, 2009, 25(4): 26-28. [12] 樊金华, 薛皎亮, 谢映平, 等. 布氏白僵菌液体培养条件的优化研究[J]. 山西大学学报(自然科学版), 2013, 36(2): 271-274. [13] STIERNAGLE T. Maintenance of C. elegans[J]. WormBook, 2006: 1-11. doi: 10.1895/wormbook.1.101.1
[14] 台莲梅, 许艳丽, 高凤昌. 尖孢镰刀菌毒素的初步研究[J]. 黑龙江八一农垦大学学报, 2004, 16(4): 9-12. [15] 周鸿媛, 唐莉莉, 路勇, 等. 脱氧雪腐镰刀菌烯醇、黄曲霉毒素B1和玉米赤霉烯酮对秀丽隐杆线虫的联合毒性研究[J]. 生态毒理学报, 2018, 13(3): 112-121. [16] 迟东泽, 何源, 刘芳芳, 等. 鹿鞭醇提物对秀丽隐杆线虫衰老的影响[J]. 食品工业科技, 2021, 42(10): 327-335. [17] YANG J, WAN Q L, MU Q Z, et al. The lifespan-promoting effect of otophylloside B in Caenorhabditis elegans[J]. Natural Products and Bioprospecting, 2015, 5(4): 177-183. doi: 10.1007/s13659-015-0064-4
[18] PUKKILA-WORLEY R, AUSUBEL F M, MYLONAKIS E. Candida albicans infection of Caenorhabditis elegans induces antifungal immune defenses [J]. PLoS Pathogens, 2011, 7(6): e1002074. doi: 10.1371/journal.ppat.1002074.
[19] 肖月, 易杏盈, 张东华, 等. 胶孢炭疽菌侵染秀丽隐杆线虫的特性研究[J]. 生物学杂志, 2023, 40(5): 24-28. [20] 郭倩楠, 周正富, 徐玉泉, 等. 圆锥掘氏梅里霉侵染秀丽隐杆线虫周期的扫描电镜观察[J]. 中国农业科技导报, 2015, 17(2): 81-86. [21] HUANG X, LI D, XI L, et al. Caenorhabditis elegans: A simple nematode infection model for Penicillium marneffei[J]. PLoS One, 2014, 9(9): e108764. doi: 10.1371/journal.pone.0108764.
[22] OKOLI I, BIGNELL E M. Caenorhabditis elegans: Aspergillus fumigatus (nematode-mould) model for study of fungal pathogenesis[J]. British Microbiology Research Journal, 2015, 7(2): 93-99. doi: 10.9734/BMRJ/2015/15838
[23] ARNONI M V, PAULA C R, AULER M E, et al. Infections caused by Fusarium species in pediatric cancer patients and review of published literature[J]. Mycopathologia, 2018, 183(6): 941-949. doi: 10.1007/s11046-018-0257-6
[24] 郭亦晨. 霉变槟榔中霉菌分离、鉴定及其对秀丽隐杆线虫的影响[D]. 长沙: 湖南农业大学, 2020.
 下载:
下载:

计量
- 文章访问数: 223
- HTML全文浏览量: 79
- PDF下载量: 44
