
Response of growth and physiological characteristics of Pennisetum hydridum to water content change in dredged soil
-
摘要:目的
研究在不同含水量疏浚土中皇竹草Pennisetum hydridum的生长、生理特性及动态变化规律,探明皇竹草在疏浚土上生长的最适含水量,为新近吹填疏浚土的生态固化提供理论依据。
方法选用皇竹草为供试植物,通过盆栽试验,设置20%(T1)、30%(T2)、40%(T3)、50%(T4) 4个含水量(w)处理,分析皇竹草的生长、养分吸收、水分蒸腾、叶片和根系生理特性。
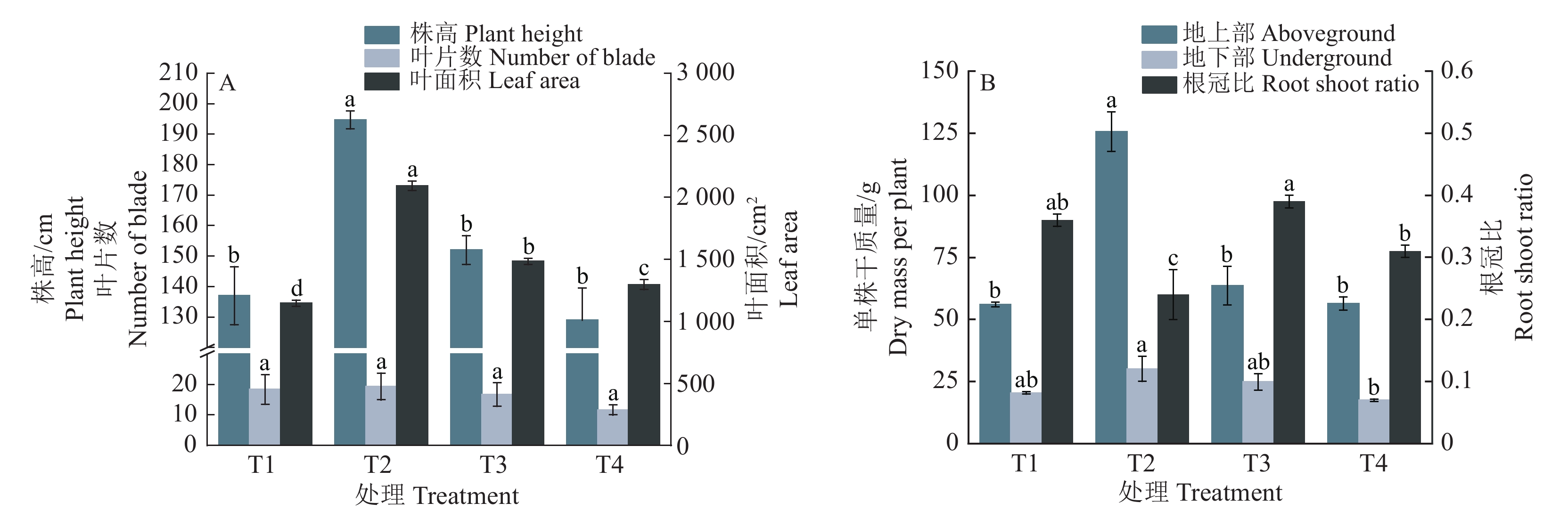
结果皇竹草在T2处理中生长表现最佳,T2处理的株高(152 cm)、单株干质量(88.51 g)以及单株氮、磷、钾吸收量(505.72、99.39、1703.45 mg)等数值均显著高于其他处理(P<0.05)。随着疏浚土含水量的增加,皇竹草的日耗水量、耗水速率、净光合速率、气孔导度和蒸腾速率均呈先升后降趋势,且均在T2处理达到最大值,并显著高于其他处理(P<0.05);皇竹草酶活性指标在T1和T4处理均高于其他处理,在T2处理达到最低。综合评价结果表明,皇竹草生长对疏浚土含水量(w)适应性强弱排序为:30%>40%>50%>20%。
结论综合各项指标,30%(w)疏浚土含水量最有利于皇竹草的生长、养分吸收和蒸腾耗水,皇竹草在水淹胁迫(T3、T4)中的各项生长指标优于干旱胁迫(T1)。以上结果可供实际生产应用中疏浚土的浅层固化、植物资源利用等参考。
Abstract:ObjectiveTo study the growth, physiological characteristics and their dynamic changes of Pennisetum hydridum in dredged soils with different water contents, find out the optimal water content for the growth of P. hydridum in dredged soils, and provide a theoretical reference for the ecological solidification method of newly blown and filled dredged soil.
MethodFour water content (w) treatments of 20% (T1), 30% (T2), 40% (T3) and 50% (T4) were set up to analyze the growth, nutrient absorption, water transpiration, leaf and root physiological characteristics of P. hydridum through potting experiment.
ResultThe growth performance of P. hydridum was the best in T2, and the plant height (152 cm), dry mass per plant (88.51 g) and N, P and K uptake per plant (505.72, 99.39, 1 703.45 mg) of T2 were significantly higher than those of other treatments (P<0.05). The daily water consumption, water consumption rate, net photosynthetic rate, stomatal conductance and transpiration rate of P. hydridum all increased first and then decreased with the increase of soil water content, and all reached the maximum in T2 and were significantly higher than those of other treatments (P<0.05). The enzyme activity indexes of P. hydridum were higher in T1 and T4 than those of other treatments, and reached the lowest in T2. The comprehensive evaluation results showed that the growth adaptability of P. hydridum to water content in dredged soil was as follows: 30% > 40% > 50% > 20%.
ConclusionBased on the various indicators, the water content of 30% in dredged soil is the most conducive to growth, nutrient absorption and transpiration and water consumption of P. hydridum, and the growth performance of P. hydridum in flooding stress (T3, T4) was better than that of drought stress (T1). These results can be used as a reference for shallow solidification of dredged soil and utilization of plant resources in practical production applications.
-
Keywords:
- Water content /
- Dredged soil /
- Pennisetum hydridum /
- Growth /
- Physiology
-
黄梁木Anthocephalus chinensis是茜草科团花属植物,产于我国广西和云南南部,东南亚及印度、斯里兰卡也有分布, 具有生长快、易管理、抗风能力强和木材产量高等特性。黄梁木不仅速生,而且材质良好,适用于作箱侧板、火柴杆、茶叶箱或其他包装箱;在建筑上可用作门窗、檀条、椽子、天花板、室内装修等[1]。因黄梁木生长速度快,被人们誉为“奇迹之树”[2]、“宝石之树”,是热带亚热带地区发展人工造林最理想的树种之一。在福建南部、广东、广西、云南、海南等大部分地区,黄梁木有望成为继桉树之后的又一个极具发展潜力的速生树种。在广东,黄梁木目前种植地区有云浮、江门、普宁、揭西、河源、湛江等地,部分地区开始用于上山造林。
林分密度是指单位面积林地上林木的数量,是反映林木对其所占空间利用程度的指标[3]。在森林培育的过程中,林分密度是制约林木生长的关键因素,对人工林生态系统的稳定性及生物生产力有重要影响。合理的造林密度既能增强林分的生长稳定性和抗逆性,又能最大限度地利用其营养空间,使林分获得最高的经济、生态效益[4]。探索合理的造林密度已成为森林培育工作的核心问题之一[5]。国内外许多学者就生产上主栽的小黑杨Populus × xiaohei、樟子松Pinus sylvestris var. mongolica和非洲破布树Mukumari Cordia africana等树种人工林的林分密度与其生长的关系进行了系统的探讨,表明造林密度对人工林树高、胸径、冠幅、单株材积等均产生显著影响。由于树种、林木年龄以及生长环境等因素各不相同,所以根据具体情况才能得出合理的生长密度[6-8]。
目前对黄梁木林分密度效应的研究尚不多见。本文对黄梁木人工幼林的个体生长与林分密度的关系进行分析,从而深入了解该树种林分密度对幼林生长和材积的影响,以期为黄梁木的人工林培育和高效利用提供科学依据。
1. 材料与方法
1.1 试验地概况
试验地设在广东省云浮市新兴县东城镇(22°22′~22°50′N,111°57′~112°31′E),气候常年温和湿润,雨量充沛,年平均气温22.1 ℃,年平均降水量1 546.5 mm,降雨主要集中在夏秋季;而冬春两季则降雨较少,经常出现旱情。年日照1 478.2 h,山地土壤多为赤红壤,呈酸性,pH5.0~5.5, 母岩为花岗岩。黏粒矿物以高岭石为主,伴生黏粒矿物有针铁矿和少量水云母,极少三水铝石。
1.2 试验方法
试验林采用完全随机区组设计,共设5个造林密度:625、667、833、1 667和2 500株·hm-2,株行距分别为4 m×4 m、3 m×5 m、3 m×4 m、3 m×2 m和2 m×2 m。设25株小区(每个小区5行,每行种植5株树),4次重复。每个小区最外围的植株为保护株,内部9株为调查(试验)株。于2012年1月布设试验地, 全面整地后,根据试验设计拉线确定植树穴位置。机械挖穴,穴规格为60 cm×60 cm×60 cm。回填土壤20 cm,每穴施堆置发酵后的猪粪15 kg(鲜质量),回填表土,使土堆略高出地面5 cm左右。2012年3月中旬,用苗龄2个月的黄梁木组培苗造林。造林后发现缺株及时补植。造林当年7月人工除草1次,每穴追施复合肥0.1 kg。之后林分很快郁闭,不再追肥和抚育。
1.3 观测指标
2014年11月,对试验林进行调查,采用每木检尺法,测量每个小区内9株试验树的胸径、树高和冠幅。因目前还未有黄梁木的材积公式,本研究中采用与黄梁木干形相近的大青杨Populus ussuriensis的立木二元材积公式:V=0.287 063D2H+0.920 037D2,计算黄梁木单株材积(V), 式中,D为胸径,H为树高[9]。
1.4 数据处理
用Excel 2003对数据进行整理,采用SPSS 18.0进行统计分析。采用单因素方差分析和Duncan’s多重极差检验法比较不同处理数据组间的差异,采用Pearson相关性分析对不同数据组间的相关性进行分析。
2. 结果与分析
2.1 造林密度对树木生长的影响
2.1.1 不同造林密度对树高的影响
由图 1A可知,黄梁木造林密度为2 500和1 667株·hm-2林分的平均树高无显著差异,但均显著高于造林密度为667和625株·hm-2林分的平均树高,且造林密度833株·hm-2林分的平均树高与造林密度为667和625株·hm-2林分的平均树高均无显著差异。造林密度为667株·hm-2的黄梁木平均树高为8.97 m, 密度增加到2 500株·hm-2时,平均树高增加10.4%。本研究中,黄梁木林分树高总体随造林密度的增加而增高。
![]() 图 1 不同造林密度下黄梁木的生长状况各图的柱子上方,凡具有一个相同小写字母者, 表示不同造林密度间差异不显著(P>0.05, Duncan’s法,n=180)Figure 1. The growth of Anthocephalus chinensis under different planting densities
图 1 不同造林密度下黄梁木的生长状况各图的柱子上方,凡具有一个相同小写字母者, 表示不同造林密度间差异不显著(P>0.05, Duncan’s法,n=180)Figure 1. The growth of Anthocephalus chinensis under different planting densities2.1.2 不同造林密度对冠幅的影响
由图 1B可知,黄梁木造林密度为667和1 667株·hm-2林分的平均冠幅与其他密度林分均无明显差异;造林密度为625和833株·hm-2林分的平均冠幅无显著差异,但均显著高于造林密度2 500株·hm-2林分的平均冠幅,625株·hm-2密度下林木的平均冠幅最大(5.8 m), 密度增加到2 500株·hm-2时,平均冠幅降低8.6%。本研究中,冠幅总体随着造林密度的增加而降低。
2.1.3 不同造林密度对枝下高的影响
由图 1C可知,黄梁木造林密度为625、667和833株·hm-2林分的平均枝下高均无明显差异,均与造林密度为1 667和2 500株·hm-2的林分平均枝下高存在显著差异,且造林密度为1 667和2 500株·hm-2林分之间平均枝下高也存在显著差异。1 667株· hm-2密度下,枝下高最高,密度由833株·hm-2增至1 667株·hm-2,枝下高增长26.6%,密度由1 667株·hm-2增至2 500株·hm-2,枝下高降低15.6%。
2.1.4 不同造林密度对胸径的影响
由图 1D可知,黄梁木造林密度为667、833、1 667和2 500株·hm-2的林分平均胸径均无显著差异(P>0.05),但均显著低于造林密度为625株·hm-2的林分(P<0.05)。造林密度为625株·hm-2的黄梁木平均胸径最大(16.3 cm), 密度增加到2 500株·hm-2,平均胸径降低12.9%。本研究中,黄梁木林分胸径总体随造林密度的增加而降低。
2.2 造林密度对材积的影响
2.2.1 不同造林密度对单株立木材积的影响
由图 2A可知,黄梁木造林密度为625株·hm-2的林分单株材积与密度为1 667株·hm-2的林分单株材积无显著差异,但显著高于造林密度为667、833和2 500株·hm-2的林分单株材积。造林密度为625株·hm-2的黄梁木单株材积为0.095 m3, 密度增加到2 500株·hm-2, 单株材积降低20%。本研究中,黄梁木造林密度对单株材积的影响和对胸径的影响是一致的。
![]() 图 2 不同造林密度下黄梁木的单株材积(A)和林分蓄积(B)各图的柱子上方,凡具有一个相同小写字母者,表示不同造林密度间差异不显著(P>0.05, Duncan’s法,n=180)Figure 2. Individual tree volumes (A) and stand volumes (B) of Anthocephalus chinensis under different planting densities
图 2 不同造林密度下黄梁木的单株材积(A)和林分蓄积(B)各图的柱子上方,凡具有一个相同小写字母者,表示不同造林密度间差异不显著(P>0.05, Duncan’s法,n=180)Figure 2. Individual tree volumes (A) and stand volumes (B) of Anthocephalus chinensis under different planting densities2.2.2 不同造林密度对林分蓄积的影响
由图 2B可知,黄梁木造林密度为2 500株·hm-2的林分蓄积最大,显著高于其他林分。密度为1 667株·hm-2的林分蓄积显著低于2 500株·hm-2的林分,但显著高于其他林分。密度为625和667株·hm-2的林分蓄积无显著差异,密度为833和625株·hm-2的林分蓄积无显著差异。造林密度为667株·hm-2的林分蓄积为50.396 m3·hm-2, 密度增加到2 500株·hm-2时,林分蓄积增加73.9%。本研究中,黄梁木林分蓄积总体随造林密度的增加而增加。
2.3 各生长性状间及与造林密度的相关性
黄梁木树高、枝下高和林分蓄积分别与造林密度呈极显著正相关关系(表 1),即林分树高、枝下高和林分蓄积随造林密度的增大而增加,方差分析结果表明,造林密度对以上3个生长性状的影响极显著(P<0.01);冠幅与造林密度呈极显著负相关(表 1),即冠幅随造林密度的增大而减少,方差分析结果表明,造林密度对冠幅的影响极显著。各生长性状之间的相关性分析表明,林分蓄积与树高、胸径与冠幅、单株材积与胸径以及林分蓄积与枝下高之间均存在着显著的正相关关系(表 1),方差分析结果表明,树高对林分蓄积、冠幅对胸径、胸径对单株材积以及枝下高对林分蓄积的影响极显著(表 1)。
表 1 黄梁木各生长性状间及与造林密度的相关性分析和回归方程Table 1. Correlation analysis of different growth traits and planting density of Anthocephalus chinensis and the regression equations
3. 讨论与结论
本研究中,不同造林密度对黄梁木的树高、冠幅、枝下高和林分蓄积有极显著影响。树高和林分蓄积均与密度呈极显著正相关关系; 冠幅与密度呈极显著负相关关系。随着造林密度的增加,平均树高和林分蓄积显著增高,平均冠幅显著下降,胸径和单株材积总体呈下降趋势。造林密度在625~1 667株·hm-2范围内,枝下高随密度的增长而增加;在1 667~2 500株·hm-2范围内,枝下高随密度的增长而降低。
造林密度对林分平均树高的影响较复杂,以往的研究结论也不一致。有些研究表明,密度对树高生长影响较弱,和林木的其他生长指标相比可认为是不受密度影响的[10-12]。而有些研究则认为树高随着密度的增大而降低[13-14]。黄梁木幼林树高随着造林密度的增加而总体增高,与徐勃等[15]对青杨Populus cathayana的研究结果一致。因为黄梁木属喜光速生树种, 对光的竞争促进了树高的生长。密度大的林分郁闭较早, 光只能从树冠上方照到树木上, 从而形成了对林木向上生长的有利条件[15]。
本研究中,黄梁木的冠幅、胸径和单株材积总体上均随造林密度的增加而降低。因在密度较小的林分中,林木的枝干有足够的伸展空间和光照条件,故胸径和冠幅生长比较快,随着林分密度的增大,林木对土壤有机质、水分以及光照等营养成分的激烈争夺,使得林木个体的生长相对减慢,这种现象与密度竞争效应的一般规律相一致[15]。本研究中,黄梁木冠幅与造林密度呈极显著负相关关系(r=-0.935,P=0.002)。林木胸径与冠幅呈极显著正相关关系(r=0.847,P=0.000),与以往研究结论一致[16-17]。密度对单株材积的影响和对胸径的影响可认为是一致的,造林密度对林木单株材积生长的影响主要取决于其胸径[18]。胸径的生长受造林密度的影响较大,因而胸径为不同密度下单株材积的决定因子。有研究表明,植株枝下高随着造林密度增大而增加,这是因为随着造林密度增大,林分地上部分平均占有生长空间减小而形成自然整枝[19]。在本研究中,造林密度为1 667株·hm-2时林分的枝下高最高,密度增大到2 500株·hm-2时,枝下高降低,因为在造林密度增加以后,一定的空间内每株黄梁木分配的营养含量降低,所以影响了枝下高的生长。
林分蓄积受单株材积和造林密度2个因子制约,幼龄林阶段造林密度为主导因子。故本次试验测量的黄梁木林分蓄积主要受造林密度的影响,两者呈极显著正相关关系(r=0.995,P=0.000)。大量的密度试验表明,在相同造林条件下,密度本身起主要作用,林分蓄积随造林密度的增大而增大,但当造林密度增大到一定程度时,密度的竞争效应明显,密度过大导致林木个体生长能力降低,产量下降[20]。有研究表明,林分蓄积随造林密度的增大而呈抛物线变化的趋势[4]。探索林木合理造林密度一直是林业研究关注的热点问题,马尾松Pinus massoniana和桉树Eucalyptus spp.等树种造林密度研究结果显示,单株材积随密度的增大而减小,林分蓄积随密度的增大而增大[21-22],本研究中亦得出相似结论。
木材是重要的自然资源,但由于大部分树木的生长周期过长,自然条件下成材时间不能满足人们对木材的需求。因此大力发展速生林木树种是解决木材资源短缺的基本途径之一[23]。本研究中,设置的黄梁木人工林的最大密度为2 500株·hm-2,且在此密度下林分蓄积和树高均最大,不但充分利用营养空间,且人工林产量高,树干笔直高挺,获得最高的经济效益。人工种植黄梁木这一具发展潜力的速生树种,具有推广应用价值。
本试验的黄梁木造林密度范围为625~2 500株·hm-2,结果只能初步确定黄梁木幼林的合理密度。随着林龄的增大,林分内树种种间关系及外界环境要求在不断变化,因此不同造林密度对黄梁木生长的影响也处在不断变化之中,应进行长期动态研究。目前对黄梁木林分密度效应的研究尚不多见,故还有待于进一步研究。
-
![]()
图 1 疏浚土不同含水量对皇竹草生长状况的影响
各图中,同一指标柱子上方的不同小写字母表示处理间差异显著(P<0.05,Duncan’s法)
Figure 1. Effects of different dredged soil water contents on the growth status of Pennisetum hydridum
In each figure, different lowercase letters on the bars of the same index indicated significant differences among different treatments (P<0.05, Duncan’s method)
![]()
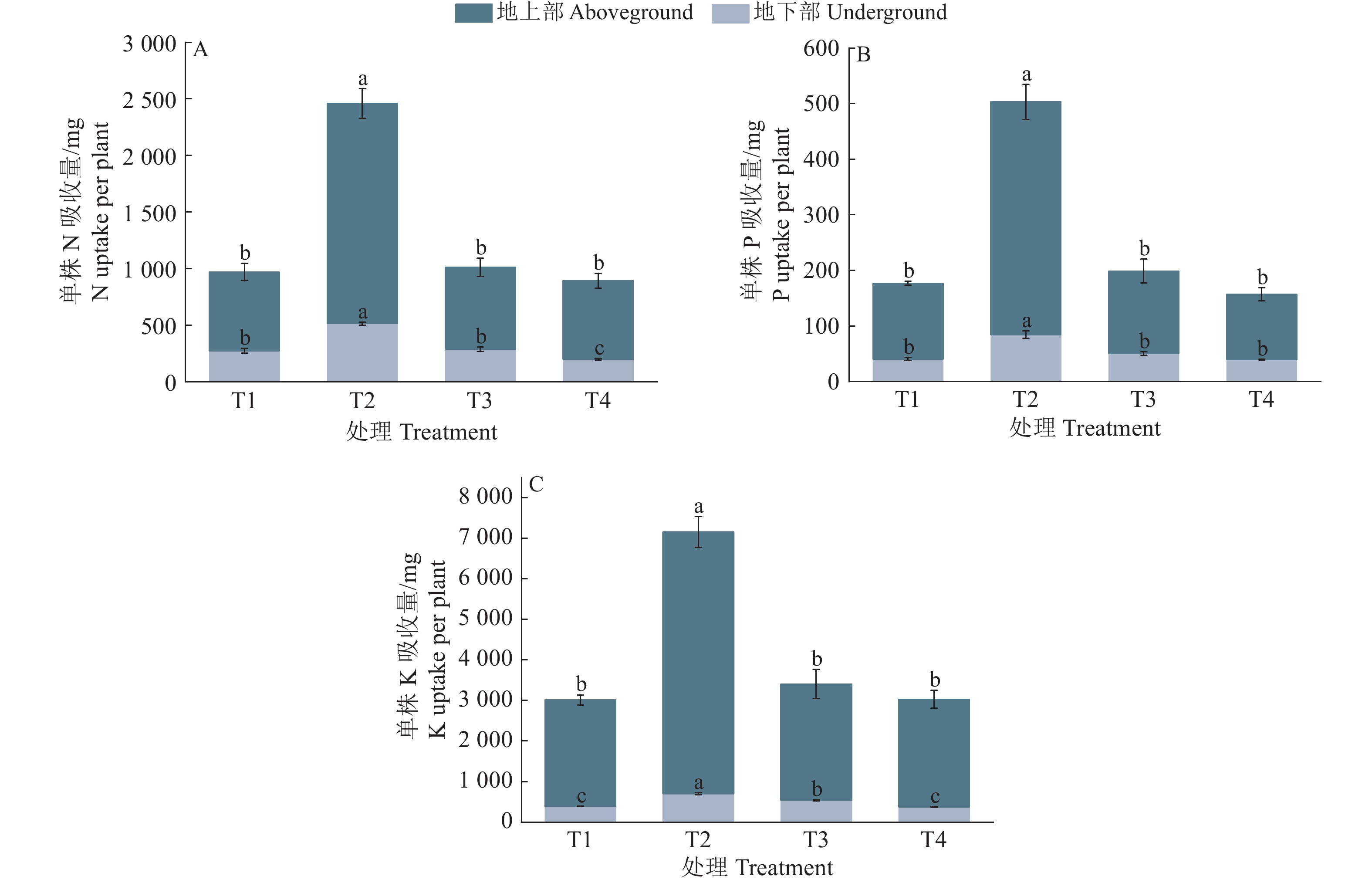
图 2 疏浚土不同含水量对皇竹草养分元素吸收量的影响
各图中,同一指标柱子上方的不同小写字母表示处理间差异显著(P<0.05,Duncan’s法)
Figure 2. Effects of different dredged soil water contents on nutrient uptake of Pennisetum hydridum
In each figure, different lowercase letters on the bars of the same index indicated significant differences among different treatments (P<0.05, Duncan’s method)
![]()
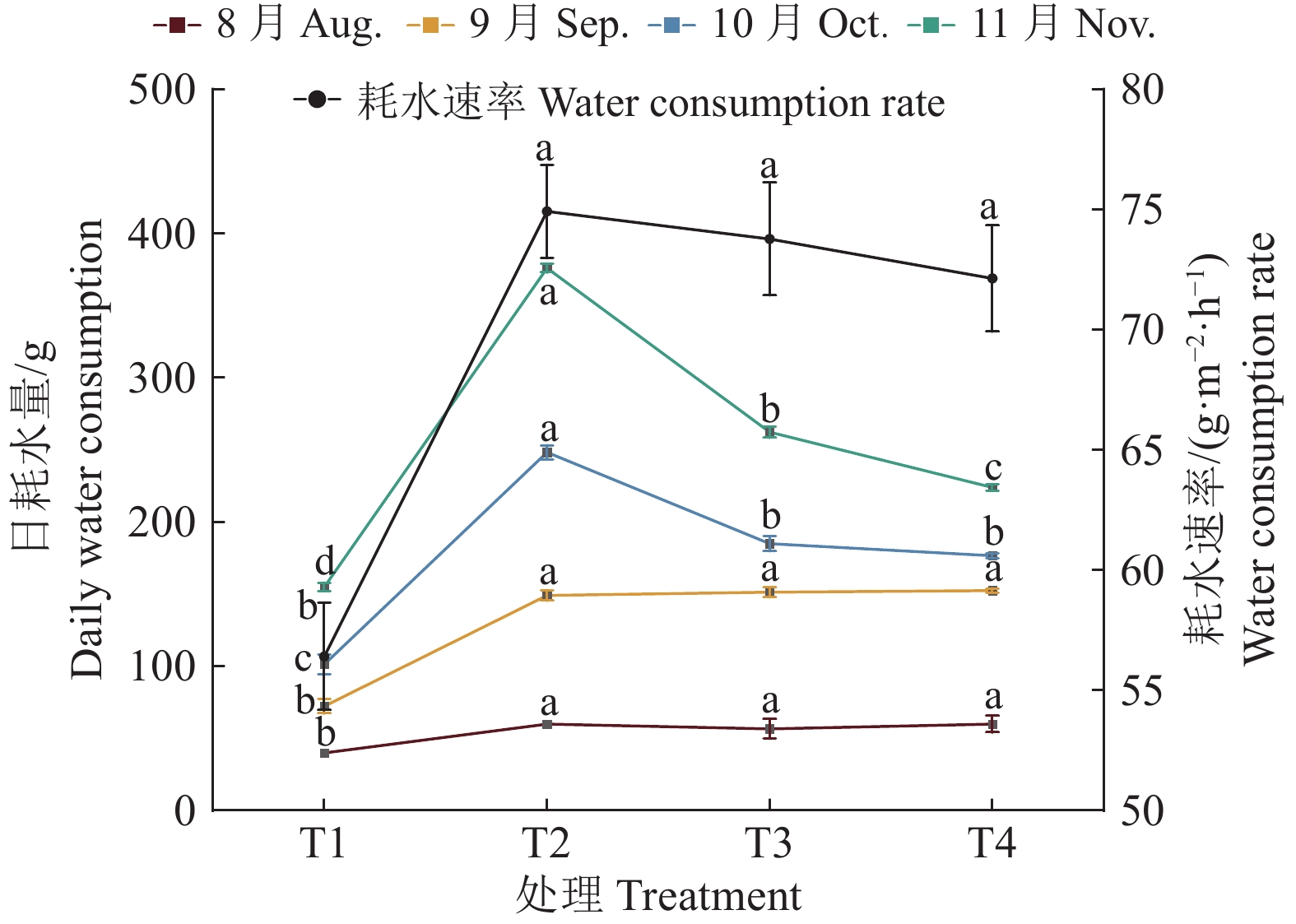
图 3 疏浚土不同含水量对皇竹草耗水量和耗水速率的影响
各折线上不同小写字母表示处理间差异显著(P<0.05,Duncan’s法)
Figure 3. Effects of different dredged soil water contents on water consumption and water consumption rate of Pennisetum hydridum
Different lowercase letters on each line indicated significant differences among different treatments (P<0.05, Duncan’s method)
![]()
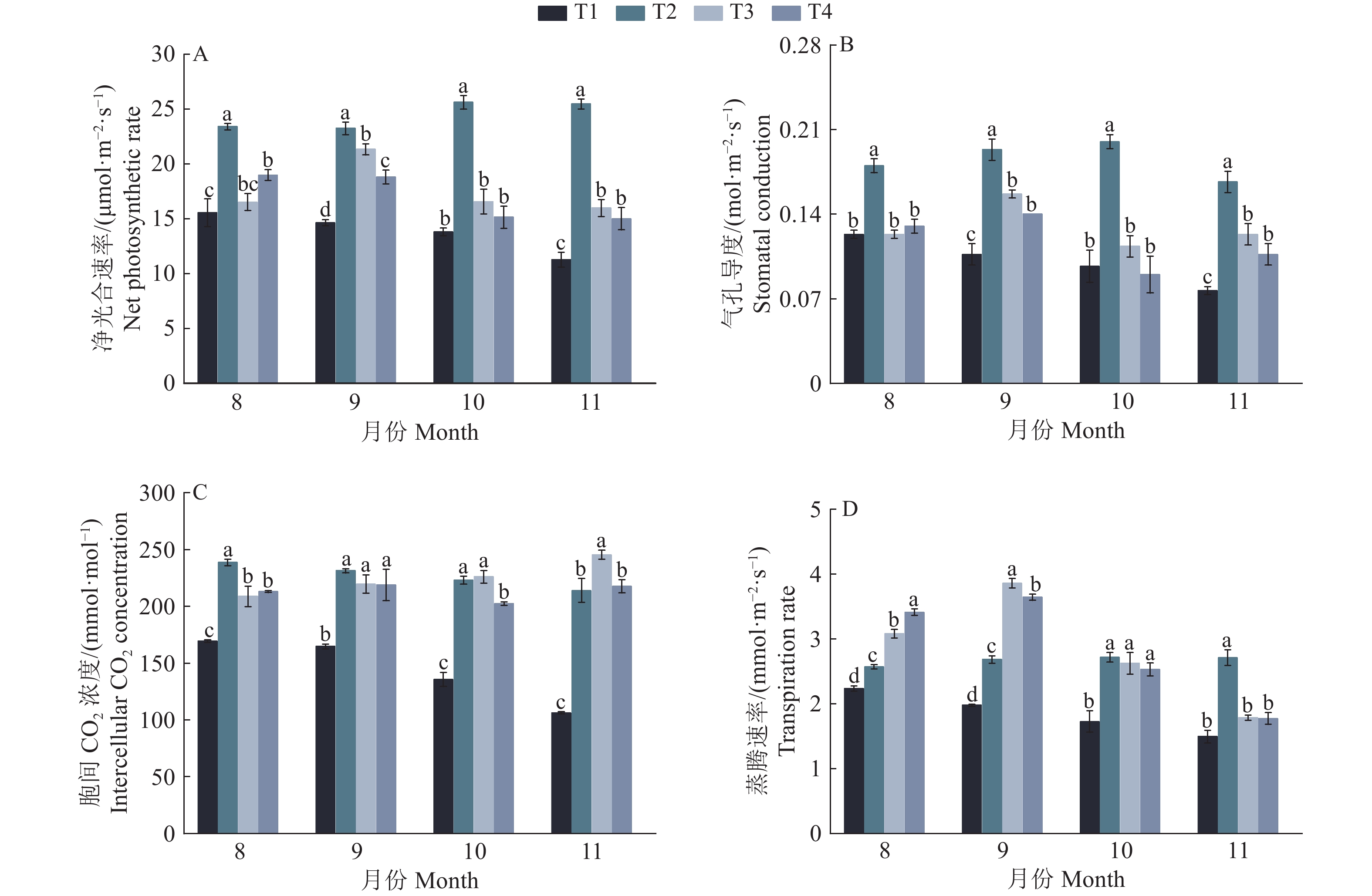
图 4 疏浚土不同含水量对皇竹草光合特性的影响
各图中,相同月份柱子上方的的不同小写字母表示处理间差异显著(P<0.05,Duncan’s法)
Figure 4. Effects of different dredged soil water contents on photosynthetic characteristics of Pennisetum hydridum
In each figure, different lowercase letters on the bars of the same month indicated significant differences among different treatments (P<0.05, Duncan’s method)
![]()
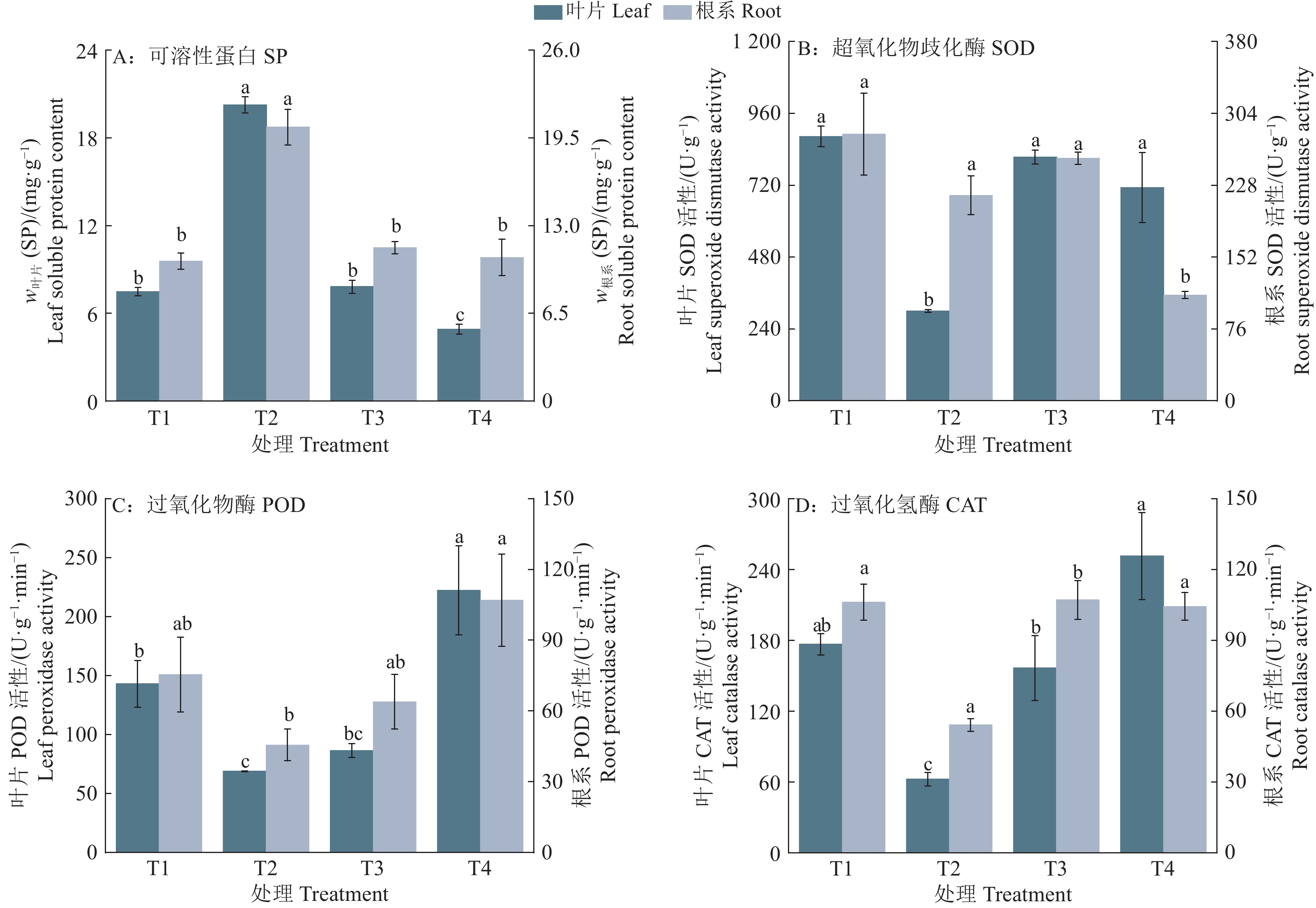
图 5 疏浚土不同含水量对皇竹草叶片和根系生理的影响
各图中,相同部位柱子上方的不同小写字母表示处理间差异显著(P<0.05,Duncan’s法)
Figure 5. Effects of different dredged soil water contents on leaf and root physiology of Pennisetum hydridum
In each figure, different lowercase letters on the bars of the same plant part indicated significant differences among different treatments (P<0.05, Duncan’s method)
表 1 主成分分析成分矩阵
Table 1 Matrix of principal component analysis component
指标
Index主成分 Principal component 1 2 3 耗水量 Water consumption 0.942 0.240 0.199 耗水速率 Water consumption rate 0.535 0.554 0.411 净光合速率 Net photosynthetic rate 0.941 0.239 0.064 气孔导度 Stomatal conduction 0.878 0.288 0.258 胞间CO2浓度 Intercellular CO2 concentration 0.369 0.648 0.629 蒸腾速率 Transpiration rate 0.955 0.202 −0.051 根系可溶性蛋白含量 Root soluble protein content 0.955 0.008 −0.119 根系超氧化物歧化酶活性 Root superoxide dismutase activity 0.022 −0.910 0.257 根系过氧化物酶活性 Root peroxidase activity −0.589 0.419 −0.220 根系过氧化氢酶活性 Root catalase activity −0.910 −0.031 0.306 叶片可溶性蛋白含量 Leaf soluble protein content 0.968 −0.176 −0.121 叶片超氧化物歧化酶活性 Leaf superoxide dismutase activity −0.869 −0.193 0.125 叶片过氧化物酶活性 Leaf peroxidase activity −0.635 0.526 −0.422 叶片过氧化氢酶活性 Leaf catalase activity −0.817 0.360 0.015 株高 Plant height 0.914 −0.098 0.014 叶片数 Number of blade 0.329 −0.507 −0.043 叶面积 Leaf area 0.969 0.119 0.109 地上部干质量 Aboveground dry mass 0.979 −0.003 −0.151 地下部干质量 Underground dry mass 0.949 −0.182 0.128 根冠比 Root shoot ratio −0.579 −0.354 0.634 地上部N吸收量 Aboveground N uptake 0.960 −0.037 −0.196 地上部P吸收量 Aboveground P uptake 0.972 −0.077 −0.157 地上部K吸收量 Aboveground K uptake 0.965 0.008 −0.198 地下部N吸收量 Underground N uptake 0.937 −0.232 −0.059 地下部P吸收量 Underground P uptake 0.969 −0.042 0.002 地下部K吸收量 Underground K uptake 0.956 −0.099 0.236 特征值 Eigenvalue 18.33 2.96 1.73 贡献率/% Contribution rate 70.51 11.37 6.63 累积贡献率/% Cumulative contribution rate 70.51 81.88 88.51  下载: 导出CSV
下载: 导出CSV
表 2 主成分分析综合得分
Table 2 Composite score of principal component analysis
处理
Treatment主成分得分 Principal component score 综合得分
Comprehensive score排序
Sort1 2 3 T1 −3.26 −2.11 −1.03 −2.61 4 T2 6.86 −0.01 −0.39 4.81 1 T3 −0.73 −0.3 2.02 −0.42 2 T4 −2.86 2.41 −0.59 −1.79 3
下载: 导出CSV
-
[1] DEVELIOGLU I, PULAT H F. Compressibility behaviour of natural and stabilized dredged soils in different organic matter contents[J]. Construction and Building Materials, 2019, 228: 116787. doi: 10.1016/j.conbuildmat.2019.116787.
[2] CHU S H, YAO J J. A strength model for concrete made with marine dredged sediment[J]. Journal of Cleaner Production, 2020, 274: 122673. doi: 10.1016/j.jclepro.2020.122673.
[3] WANG L, YEUNG T L K, LAU A Y T, et al. Recycling contaminated sediment into eco-friendly paving blocks by a combination of binary cement and carbon dioxide curing[J]. Journal of Cleaner Production, 2017, 164: 1279-1288. doi: 10.1016/j.jclepro.2017.07.070
[4] WANG L, SHAO Y, ZHAO Z, et al. Optimized utilization studies of dredging sediment for making water treatment ceramsite based on an extreme vertex design[J]. Journal of Water Process Engineering, 2020, 38: 101603. doi: 10.1016/j.jwpe.2020.101603.
[5] PARK J, SON Y, NOH S, et al. The suitability evaluation of dredged soil from reservoirs as embankment material[J]. Journal of Environmental Management, 2016, 183: 443-452.
[6] TODA K, KIKUCHI R, OTAKE T, et al. Effect of soil organic matters in dredged soils to utilization of their mixtures made with a steel slag[J]. Materials, 2020, 13(23): 5450. doi: 10.3390/ma13235450.
[7] 吴华林, 赵德招, 程海峰. 我国疏浚土综合利用存在问题及对策研究[J]. 水利水运工程学报, 2013(1): 8-14. [8] 张更生, 徐继涛, 尹崧宇. 疏浚吹填粉土固化室内试验[J]. 水运工程, 2018(9): 54-58. [9] 陈彦霖. 疏浚淤泥的固化处理技术与资源化利用[J]. 中国设备工程, 2020(20): 11-12. [10] STEPPE K, VANDEGEHUCHTE M W, TOGNETTI R, et al. Sap flow as a key trait in the understanding of plant hydraulic functioning[J]. Tree Physiology, 2015, 35(4): 341-345. doi: 10.1093/treephys/tpv033
[11] ZHAO W, DONG X, WU Z, et al. Using infrared thermal imaging technology to estimate the transpiration rate of citrus trees and evaluate plant water status[J]. Journal of Hydrology, 2022, 615: 128671. doi: 10.1016/j.jhydrol.2022.128671.
[12] 陈广明, 张祝培, 毕世明, 等. 利用皇竹草生态固化浅表层疏浚土试验研究[J]. 水运工程, 2021(9): 16-22. [13] ZHAO J, ZHANG W, WANG K, et al. Responses of the soil nematode community to management of hybrid napiergrass: The trade-off between positive and negative effects[J]. Applied Soil Ecology, 2014, 75: 134-144. doi: 10.1016/j.apsoil.2013.10.011
[14] 何琳, 祝其丽, 王彦伟, 等. 多年生禾草皇竹草的综合利用研究进展[J]. 应用与环境生物学报, 2020, 26(3): 705-712. [15] KANCHI G M, NEERAJA V S, SIVAKUMAR BABU G L S. Effect of anisotropy of fibers on the stress-strain response of fiber-reinforced soil[J]. International Journal of Geomechanics, 2015, 15(1): 06014016. doi: 10.1061/(asce)gm.1943-5622.0000392.
[16] 生态环境部, 国家市场监督管理总局. 土壤环境质量 农用地土壤污染风险管控标准(试行): GB 15618—2018[S]. 北京: 中国标准出版社, 2018. [17] 鲍士旦. 土壤农化分析[M]. 3版. 北京: 中国农业出版社, 2000. [18] 李合生, 王学奎. 现代植物生理学[M]. 4版. 北京: 高等教育出版社, 2019. [19] SUN Y, WANG C, CHEN H Y H, et al. Response of plants to water stress: A meta-analysis[J]. Frontiers in Plant Science, 2020, 11: 978. doi: 10.3389/fpls.2020.00978.
[20] YANG W, LIN K, WU C, et al. Effects of waterlogging with different water resources on plant growth and tolerance capacity of four herbaceous flowers in a bioretention basin[J]. Water, 2020, 12(6): 1619. doi: 10.3390/w12061619.
[21] KARIMI M, AHMADI A, HASHEMI J, et al. Plant growth retardants (PGRs) affect growth and secondary metabolite biosynthesis in Stevia rebaudiana Bertoni under drought stress[J]. South African Journal of Botany, 2019, 121: 394-401. doi: 10.1016/j.sajb.2018.11.028
[22] XIAO C, SUN O J, ZHOU G, et al. Interactive effects of elevated CO2 and drought stress on leaf water potential and growth in Caragana intermedia[J]. Trees - Structure and Function, 2005, 19(6): 711-720.
[23] MEZGEBE A, AZEREFEGNE F. Effect of water stress on glucosinolate content of Brassica carinata and performance of Brevicoryne brassicae and Myzus persicae[J]. International Journal of Tropical Insect Science, 2021, 41(2): 953-960. doi: 10.1007/s42690-020-00340-3
[24] PATEL J, MISHRA A. Plant aquaporins alleviate drought tolerance in plants by modulating cellular biochemistry, root-architecture, and photosynthesis[J]. Physiologia Plantarum, 2021, 172(2): 1030-1044. doi: 10.1111/ppl.13324
[25] ZHOU G, ZHOU X, NIE Y, et al. Drought-induced changes in root biomass largely result from altered root morphological traits: Evidence from a synthesis of global field trials[J]. Plant, Cell & Environment, 2018, 41(11): 2589-2599.
[26] ARANJUELO I, MOLERO G, ERICE G, et al. Plant physiology and proteomics reveals the leaf response to drought in alfalfa (Medicago sativa L. )[J]. Journal of Experimental Botany, 2011, 62(1): 111-123. doi: 10.1093/jxb/erq249
[27] 崔东阳, 贺亮亮, 刘崇怀, 等. 淹水胁迫对夏黑葡萄生理特性和品质的影响[J]. 生态学杂志, 2022, 41(12): 2344-2351. [28] PETERS E B, MCFADDEN J P, MONTGOMERY R A. Biological and environmental controls on tree transpiration in a suburban landscape[J]. Journal of Geophysical Research, 2010, 115(G4): G04006. doi: 10.1029/2009jg001266.
[29] 张婵, 洪希群, 吴承祯, 等. 干旱胁迫对闽北乡土树种耗水及光合特性的影响[J]. 应用与环境生物学报, 2023, 29(1): 212-219. [30] 吴俊文, 刘珊, 李吉跃, 等. 干旱胁迫下广东石漠化地区造林树种光合和耗水特性[J]. 生态学报, 2016, 36(11): 3429-3440. [31] BLANKENAGEL S, YANG Z, AVRAMOVA V, et al. Generating plants with improved water use efficiency[J]. Agronomy, 2018, 8(9): 194. doi: 10.3390/agronomy8090194.
[32] 欧芷阳, 庞世龙, 谭长强, 等. 干旱胁迫对桂西南石漠化地区主要造林树种光合与耗水特性的影响[J]. 生态学杂志, 2020, 39(10): 3237-3246. [33] 李吉跃, 周平, 招礼军. 干旱胁迫对苗木蒸腾耗水的影响[J]. 生态学报, 2002(9): 1380-1386. [34] 陈玉, 贾剑波, 颜成正, 等. 干旱胁迫下杉木叶片吸水及水分逆向运移特征[J]. 生态学杂志, 2023, 42(9): 2091-2099. [35] 常宏达, 吕德生, 王振华, 等. 滴灌量和有机无机肥等氮配施对葡萄产量及品质的影响[J]. 干旱地区农业研究, 2022, 40(4): 116-123. [36] 罗成威, 王若水, 王莉莎, 等. 不同水热调控方式对苹果−大豆间作系统土壤温度和根系分布的影响[J]. 水土保持学报, 2023, 37(2): 217-226. [37] 赵经华, 杨庭瑞, 胡文军, 等. 水氮互作对滴灌小麦土壤硝态氮运移、氮平衡及水氮利用效率的影响[J]. 中国农村水利水电, 2021(4): 141-149. [38] CALZADILLA P I, CARVALHO F E L, GOMEZ R, et al. Assessing photosynthesis in plant systems: A cornerstone to aid in the selection of resistant and productive crops[J]. Environmental and Experimental Botany, 2022, 201: 104950. doi: 10.1016/j.envexpbot.2022.104950.
[39] WERTIN T M, BELNAP J, REED S C. Experimental warming in a dryland community reduced plant photosynthesis and soil CO2 efflux although the relationship between the fluxes remained unchanged[J]. Functional Ecology, 2017, 31(2): 297-305. doi: 10.1111/1365-2435.12708
[40] 李志鹏, 万素梅, 胡守林, 等. 不同灌水频率和灌溉定额对南疆无膜棉蕾铃时空分布及产量形成的影响[J]. 棉花学报, 2022, 34(5): 383-400. [41] DE OLLAS C, DODD I C. Physiological impacts of ABA-JA interactions under water-limitation[J]. Plant Molecular Biology, 2016, 91(6): 641-650. doi: 10.1007/s11103-016-0503-6
[42] ZHANG X, SUN W, CHEN X, et al. Integrated physiological and transcriptomic analysis reveals mechanism of leaf in Phellodendron chinense Schneid. seedlings response to drought stress[J]. Industrial Crops and Products, 2023, 198: 116679. doi: 10.1016/j.indcrop.2023.116679.
[43] 张蓉, 董禄信, 孙长红, 等. 秸秆粉碎覆盖沟播对旱地冬小麦产量及土壤水热特征的影响[J]. 节水灌溉, 2021(7): 60-64. [44] 赵文哲, 刘晓, 杜桂英, 等. 干旱胁迫及复水对M9T337苹果砧木苗生理特性的影响[J]. 山东农业科学, 2022, 54(4): 49-54,61. [45] RAKHRA G, KAUR T, VYAS D, et al. Molecular cloning, characterization, heterologous expression and in-silico analysis of disordered boiling soluble stress-responsive wBsSRP protein from drought tolerant wheat cv. PBW 175[J]. Plant Physiology and Biochemistry, 2017, 112: 29-44. doi: 10.1016/j.plaphy.2016.12.017
[46] 桑子阳, 马履一, 陈发菊. 干旱胁迫对红花玉兰幼苗生长和生理特性的影响[J]. 西北植物学报, 2011, 31(1): 109-115. [47] DE CÁSSIA ALVES R, OLIVEIRA K R, LUCIO J C B, et al. Exogenous foliar ascorbic acid applications enhance salt-stress tolerance in peanut plants through increase in the activity of major antioxidant enzymes[J]. South African Journal of Botany, 2022, 150: 759-767. doi: 10.1016/j.sajb.2022.08.007
[48] ABABAF M, OMIDI H, BAKHSHANDEH A. Changes in antioxidant enzymes activities and alkaloid amount of Catharanthus roseus in response to plant growth regulators under drought condition[J]. Industrial Crops and Products, 2021, 167: 113505. doi: 10.1016/j.indcrop.2021.113505.
[49] YAVUZ D, KILIC E, SEYMEN M, et al. The effect of irrigation water salinity on the morph-physiological and biochemical properties of spinach under deficit irrigation conditions[J]. Scientia Horticulturae, 2022, 304: 111272. doi: 10.1016/j.scienta.2022.111272.
[50] ÁVILA-LOVERA E, WINTER K, GOLDSMITH G R. Evidence for phylogenetic signal and correlated evolution in plant-water relation traits[J]. New Phytologist, 2023, 237(2): 392-407. doi: 10.1111/nph.18565
[51] DING C, XU C, LU B, et al. Comprehensive evaluation of rice qualities under different nitrogen levels in South China[J]. Foods, 2023, 12(4): 697. doi: 10.3390/foods12040697.


计量
- 文章访问数: 231
- HTML全文浏览量: 45
- PDF下载量: 46
