Analysis of germination characteristics and differential gene expression in neo-tetraploid rice under NaCl stress
-
摘要:目的
评价新型四倍体水稻在不同浓度NaCl处理下的种子萌发特性、幼苗建成期形态指标和根部细胞结构差异,评估盐胁迫相关基因的表达差异。
方法在不同浓度NaCl (0、50、100、150、200和250 mmol/L)处理下,以‘华多1号’为研究材料,统计其种子在萌发时间、发芽势、发芽率和萌发指数指标的差异。在幼苗形成期,统计和比较‘华多1号’在不同浓度NaCl处理下的总根数、幼苗苗长、根长、幼苗鲜质量、含水率和根冠比参数的差异。利用WE-CLSM和塑料半薄切片技术观察根尖组织细胞的形态。利用基因组重测序、生物信息学分析和qRT-PCR技术对‘华多1号’与双亲相关的盐胁迫基因进行分析和定量验证。
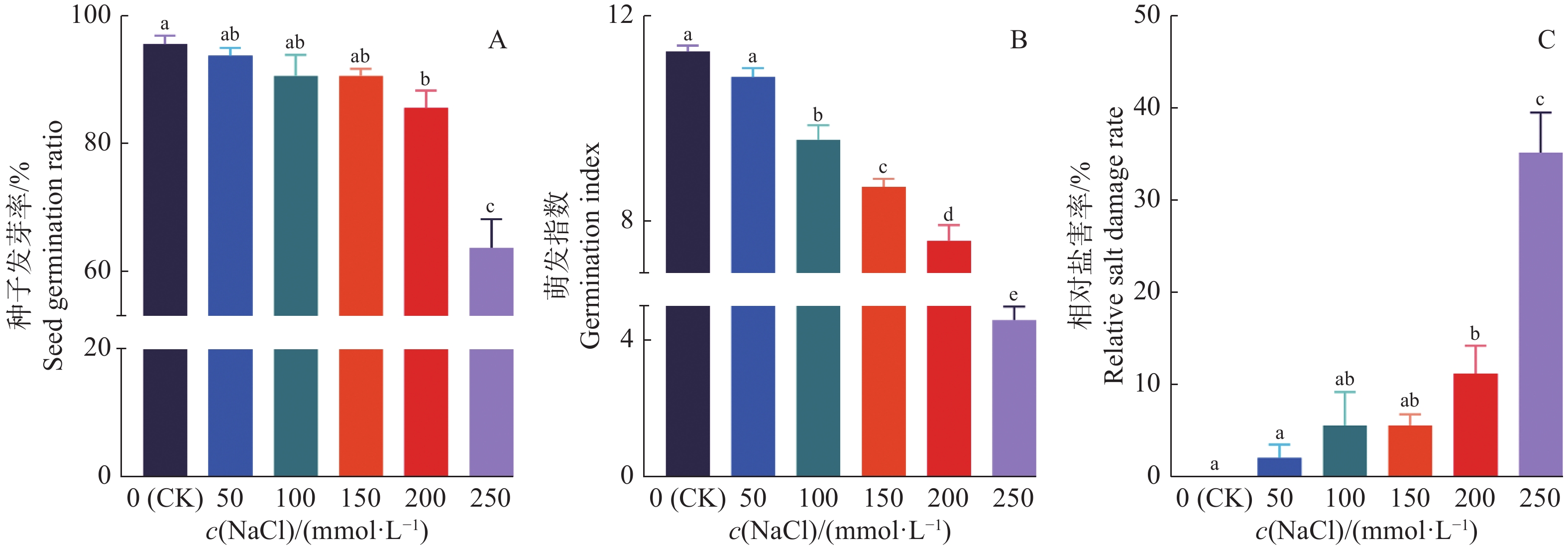
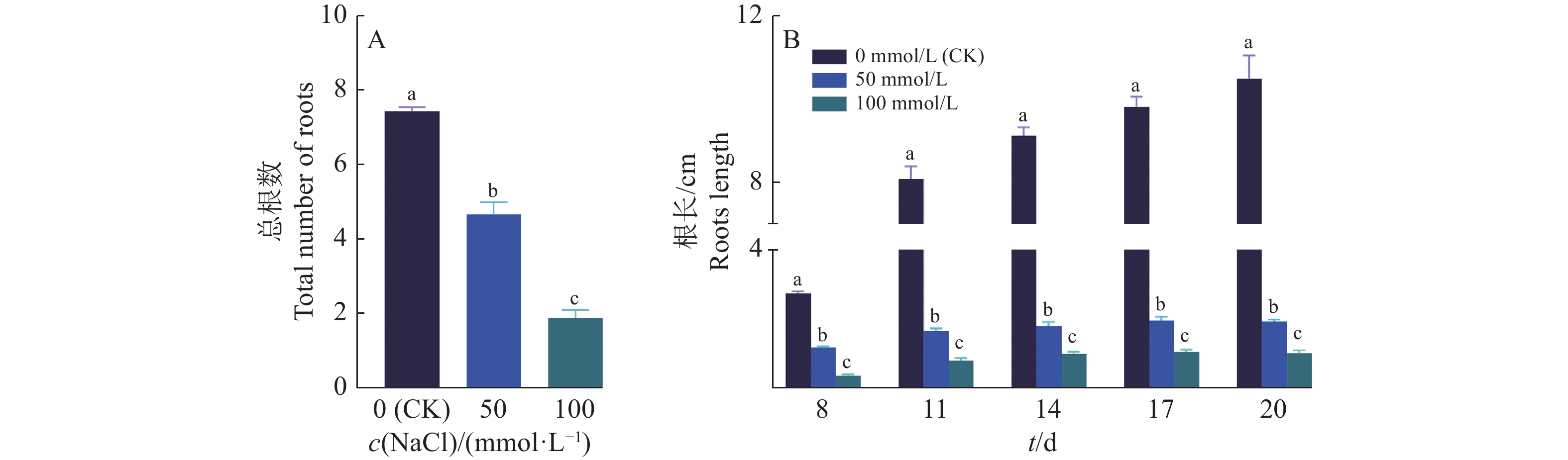
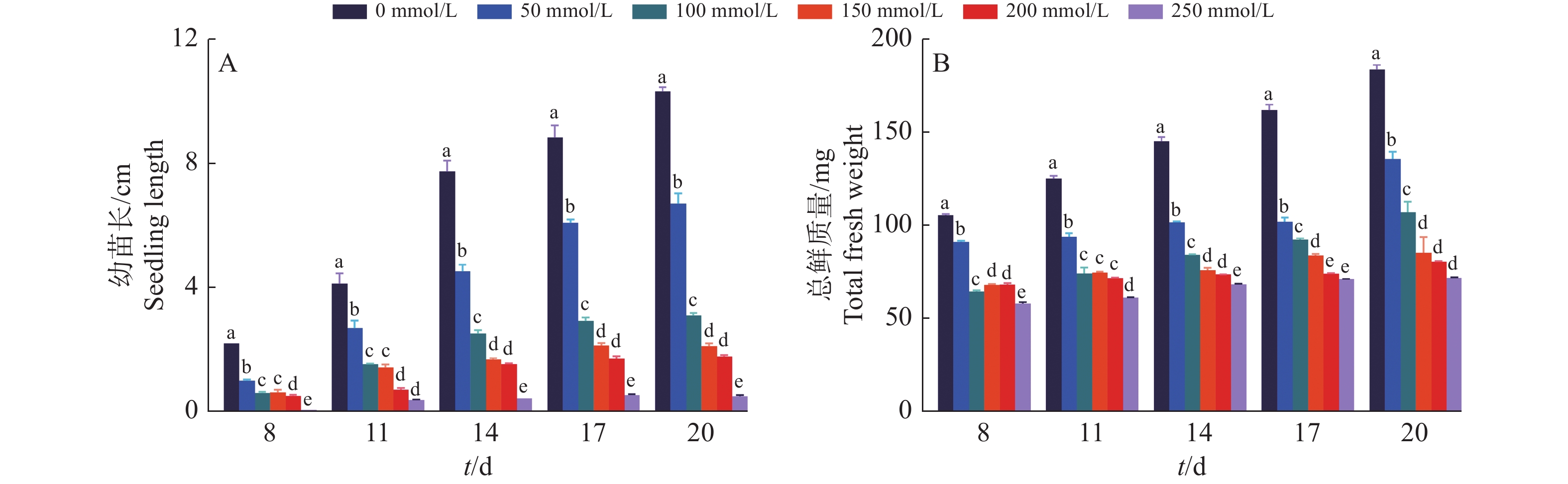
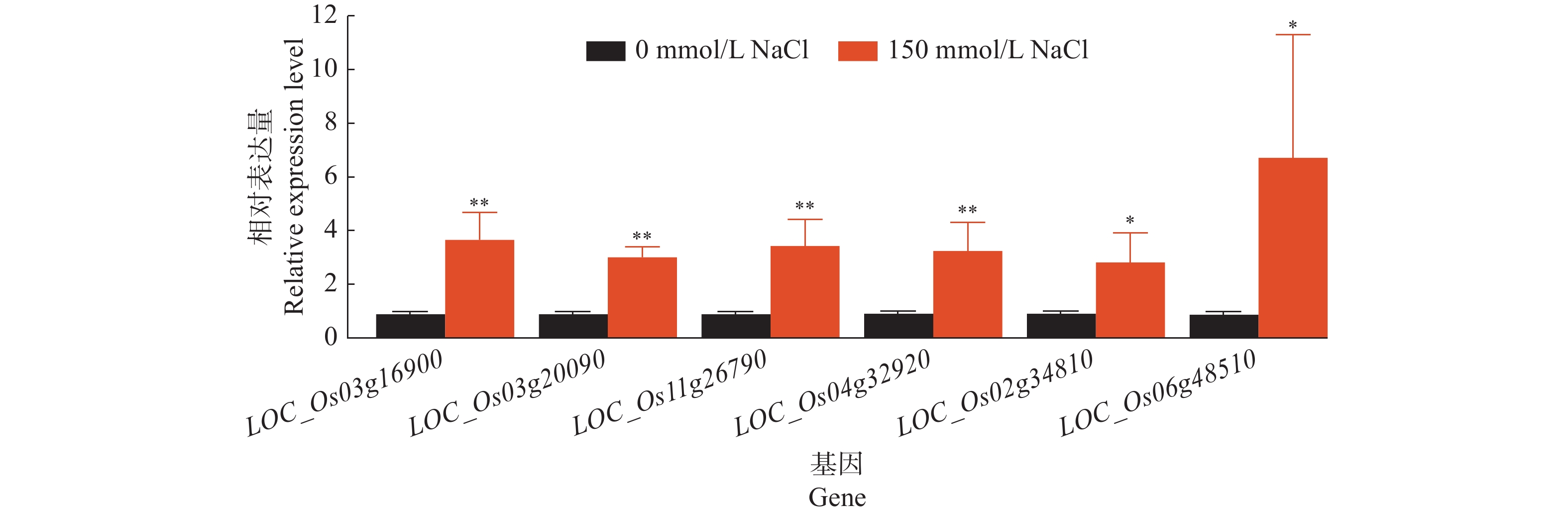
结果当NaCl浓度升高时,‘华多1号’种子发芽率和萌发指数呈递减趋势,但相对盐害率和平均发芽时间呈递增趋势。当NaCl浓度高达250 mmol/L时,‘华多1号’种子相对盐害率为35%,表明其具有较强的耐盐性。在NaCl处理8~20 d后,‘华多1号’幼苗随着NaCl浓度升高,总根数呈减少趋势,同时幼苗苗长、根长、幼苗鲜质量和含水率呈下降趋势,但根冠比呈上升趋势。NaCl胁迫会抑制‘华多1号’幼苗生长,导致其根尖细胞体积变小,细胞皱缩和细胞排列松散;盐胁迫条件下,根尖伸长区的表皮和外皮层的细胞会增厚,中间皮层的细胞层数增加,中柱直径缩小,木质部导管分化数目增加。在‘华多1号’中发现有85个盐胁迫基因在CDS区中较双亲存在差异,包括53个盐敏感相关基因、32个盐胁迫抗性相关基因。qRT-PCR结果表明:与对照相比,‘华多1号’的4个盐敏感基因(LOC_Os03g16900、LOC_Os06g48510、LOC_Os02g34810和LOC_Os04g32920)和2个耐盐性基因(LOC_Os03g20090和LOC_Os11g26790)表达量显著上升。
结论该研究可为后续系统评价四倍体水稻种质资源的耐盐性、揭示四倍体水稻的耐盐性调控机理,以及后续筛选耐盐多倍体水稻品种提供理论依据。
Abstract:ObjectiveThe purpose of this research was to evaluate differences in seed germination characteristics, morphological indexes, and root cell structure in ‘Huaduo1’ under different concentrations of NaCl and to evaluate the gene expression differences of salt stress related genes in ‘Huaduo1’.
MethodUsing ‘Huaduo1’ as the research material, the differences in seed germination time, germination potential, germination rate, and germination index under various concentrations of NaCl (0, 50, 100, 150, 200, and 250 mmol/L) were analyzed. The differences in total root number, seedling length, root length, seedling fresh weight, water content, and root-to-head ratio of ‘Huaduo1’ at seedling forming stage under various concentrations of NaCl were summarized and compared. Root tip cell morphology of ‘Huaduo1’ was observed by the WE-CLSM and semi-thin section analysis. Genomic resequencing, bioinformatics analysis, and qRT-PCR analysis were conducted to analyze and quantify the expression of the salt stress genes in ‘Huaduo1’ related to its parents.
ResultThe seed germination rate and germination index of ‘Huaduo1’ decreased with increased NaCl concentration, while the relative salt damage rate and average germination time increased. ‘Huaduo1’ showed a strong salt tolerance with the relative salt damage rate reached 35% under 250 mmol/L NaCl treatment. After 8 to 20 days of NaCl treatment, the total root number of ‘Huaduo1’ seedlings decreased with increased NaCl concentration. Meanwhile, the seedling length, root length, fresh weight and water content of seedlings under NaCl stress showed a decreasing trend, and the root-to-head ratio showed an increasing trend. NaCl stress inhibited the growth of the ‘Huaduo1’ seedlings, resulting in smaller cell volume, cell shrinkage, and loose cell arrangement in the root tip elongation zone. Moreover, the cells of the epidermis and outer cortex layer in the root tip elongation zone were thickened, the number of cell layers in the mesocortex increased, the diameter of the middle column decreased, and the number of xylem ductal differentiation increased under salt stress. Totally 85 salt stress-related genes of ‘Huaduo1’ showed variance in the CDS region compared with its two parents, including 53 salt sensitivity related genes and 32 salt stress tolerance related genes. The qRT-PCR results showed that the expressions of four salt sensitivity genes (LOC_Os03g16900, LOC_Os06g48510, LOC_Os02g34810, and LOC_Os04g32920) and two salt tolerance genes (LOC_Os03g20090 and LOC_Os11g26790) were significantly increased compared with the control.
ConclusionThe study can be used to evaluate the salt tolerance of tetraploid rice germplasm, reveal the regulation mechanism of salt tolerance in tetraploid rice, and provide a theoretical basis of the salt-tolerant germplasm selection in polyploidy rice.
-
Keywords:
- Neo-tetraploid rice /
- NaCl stress /
- Seed germination /
- Gene expression
-
动物肠道存在数量众多和种类各异的微生物,既包括乳酸杆菌和双歧杆菌等有益菌,也存在沙门菌和大肠埃希菌等致病菌或条件致病菌[1].肠道内正常微生物菌群不仅是肠道内抵抗外来病原菌以及内源条件致病菌的第一道防线,而且也是影响动物肠道发育和消化吸收功能的重要因素[2-3].仔猪,尤其是从出生到断奶前后的仔猪,由于肠道发育尚不成熟,其肠道微生态的平衡容易受到气候、断奶应激和饲料等因素的影响,从而造成微生物菌群失调,病原微生物感染,进而出现腹泻甚至导致仔猪死亡[4].因此,如何调节仔猪肠道微生物区系,改变有益菌和病原菌的比例,对养猪生产具有重要意义.
长期以来,抗生素一直被认为是抑制猪肠道病原微生物的首选添加剂.但抗生素滥用所造成的药物残留和细菌耐药性等问题也日益凸显.近年来的大量动物试验研究表明,酸制剂在降低肠道pH、提高饲料蛋白消化率和抑制有害微生物生长等方面均发挥着重要的作用[5],因而被认为是能有效替代抗生素的饲料添加剂之一[6].目前养殖生产中常用的酸制剂的种类和组合多样,既包括各种单一有机酸及其盐和无机酸,又包括各种复合酸[7].由于动物活体试验存在干扰因素多,动物个体差异大和试验重复性差等限制,导致不同酸制剂以及不同酸制剂组合之间的效果往往难以进行客观的评价和系统性地比较.
为快速便捷地测定有机酸对有害微生物的抑菌效应,研究人员多选择标准菌株进行药敏试验[8-9].但标准菌株既不能完全代表肠道野生菌株,同时又忽略了肠道内多种微生物之间的互作效应,其研究结果往往也很难在活体动物试验中得到重复验证.因此,本试验利用体外厌氧共培养技术,分离培养仔猪小肠内微生物,并利用16S rRNA定量分析食糜内微生物菌群的数量.研究结果不仅可以为有机酸的效价分析提供有效的体外模型,而且对于养猪生产中有机酸种类和剂量的选择同样具有指导意义.
1. 材料与方法
1.1 试验材料
有机酸:甲酸、乙酸、丙酸、丁酸、乳酸、富马酸、柠檬酸、苯甲酸、苹果酸和山梨酸,均为广州吉祥生物科技有限公司产品,为分析纯.
1.2 试验动物和厌氧培养
每次选择2头35日龄健康断奶仔猪(华南农业大学原种猪场),于代谢笼中饲养(饲粮配方见表 1)5 d后,屠宰取其空肠和回肠内食糜,放入冰浴的烧杯中,用保鲜膜封口后置于厌氧工作站(试验开始前用氮气和二氧化碳混合气体将厌氧工作站内氧气排尽,温度调节至39 ℃)中,并搅拌混匀,按照每管300 μL食糜分装于40个2 mL离心管中.40个离心管随机分成4组,即未培养组、空白培养组、5 mg·mL-1氨苄青霉素组(5 mg·mL-1 AMP)和10 mg·mL-1氨苄青霉素组(10 mg·mL-1 AMP).其中,未培养组和空白培养组添加100 μL无菌生理盐水,抗生素组添加不同剂量氨苄青霉素.除未培养组外,其余3组均厌氧培养4 h后立即置于-20 ℃保存备用.采用定量PCR检测微生物菌群,每个处理设10个重复.
表 1 饲粮配方表(风干基础)Table 1. Diet formulations (air dry basis)
1.3 有机酸对全食糜厌氧培养体系微生物菌群的影响
按照“1.2”方法制备仔猪肠道食糜,分别采用30 mmol·L-1的甲酸、乙酸、丙酸、丁酸、乳酸、富马酸、柠檬酸、苯甲酸、苹果酸和山梨酸培养食糜4 h.采用定量PCR检测有机酸对食糜内微生物菌群的影响.随后选择抑菌活性较强的甲酸、乙酸、丁酸、乳酸、柠檬酸,每种有机酸分别设置5、10、15、20、25和30 mmol·L-1浓度梯度,研究这些有机酸抑菌活性的浓度梯度效应.有机酸与食糜厌氧条件下共培养4 h后立即置于-20 ℃保存备用.每个处理设10个重复.
1.4 引物设计
根据NCBI中GenBank发表的基因序列,用Primer Premier 5.0软件设计不同菌属16S rRNA的上、下游引物(表 2),引物由北京奥科鼎盛生物科技有限公司合成.
表 2 荧光定量PCR引物一览表Table 2. Primer sequences used in real-time PCR analyses
1.5 细菌DNA的提取
食糜解冻后,定量称取300 mg于离心管中,加入50 μL溶菌酶,轻轻振荡混匀,于37 ℃水浴放置30 min,随后加入40 μL的0.1 g·mL-1 SDS和10 μL蛋白酶K,65 ℃水浴20 min后加入5 mol·L-1 NaCl 100 μL于65 ℃水浴10 min.采用酚/三氯甲烷/异戊醇法提取DNA.所得DNA用体积分数为75%的预冷乙醇洗涤沉淀并干燥,最后用含有终质量浓度为20 μg·μL-1 RNAase的灭菌TE Buffer溶解沉淀,于37 ℃水浴中孵育30 min.所得DNA样品置于-20 ℃保存备用.
1.6 16S rRNA定量分析
采用Mx3005P型实时荧光定量PCR仪进行扩增与分析,20 μL反应体系包括基因组DNA 1 μL,上、下游引物(10 μmol · L-1)1 μL,RealtimePCR Master Mix(含荧光染料SYBR green)10 μL,ddH2O 8 μL.反应程序为:95 ℃预变性1 min;95 ℃变性15 s,58 ~ 61 ℃退火15 s,72 ℃延伸30 s,共40个循环后结束.PCR产物胶回收后制作标准曲线.扩增结果用Mx3005P型实时荧光定量PCR仪进行分析并制作标准曲线.细菌数量(以拷贝数计)采用下列公式计算:

式中,C表示各样品根据标准曲线计算得到的DNA浓度,单位μg·μL-1;S表示待测细菌基因组DNA的定量PCR产物的碱基数;V表示从单位质量食糜中获取的DNA溶液体积,单位μL.
1.7 数据统计
数据采用SPSS19.0软件进行统计分析,检验试验数据的正态分布性,对不同处理组间的数值进行单因素方差分析(Analysis of variance,ANOVA)和独立t检验(Independent samples t test).数值用平均值±标准误表示.
2. 结果与分析
2.1 断奶仔猪肠道食糜厌氧培养效果鉴定
经过4 h厌氧培养后,食糜中总菌和各菌属的数量均急剧增加,添加5和10 mg·mL-1氨苄青霉素可极显著抑制总菌、沙门菌属和大肠埃希菌的数量(表 3).氨苄青霉素对乳杆菌属和双歧杆菌属也有显著抑制作用,但抑菌活性相对较弱.上述结果表明该体外培养模型可用于后续试验.
表 3 断奶仔猪肠道食糜体外厌氧培养效果1)Table 3. The construction of in vitro anaerobic culture system for total chyme from the small intestine of weaned piglets
2.2 不同种类有机酸对仔猪全食糜培养体系微生物菌群的影响
试验选取的10种有机酸对微生物菌群的影响如表 4所示.结果发现:与对照组相比,30 mmol·L-1甲酸、乙酸、丙酸、丁酸、乳酸和山梨酸能提高乳杆菌属数量2倍以上,而丁酸、乳酸和山梨酸能显著提高双歧杆菌属数量1.4 ~ 2.0倍.对于病原菌,柠檬酸、甲酸、乙酸和苹果酸表现出对大肠埃希菌和沙门菌属较强的抑菌效应.与对照组相比,柠檬酸、甲酸、乙酸、苹果酸、丁酸和乳酸处理组沙门菌数仅分别为对照组的5.2%、7.8%、10.0%、10.1%、12.6%和17.4%,大肠埃希菌数仅分别为对照组的5.5%、11.2%、12.7%、16.1%、20.2%和40.0%.因此,30 mmol·L-1情况下综合各种有机酸对有益菌和有害菌的效应,甲酸和乙酸可以有效提高乳杆菌的数量,抑制有害菌的繁殖,但对双歧杆菌没有显著的促进作用;柠檬酸对有害菌具有最强的抑制作用,但是对有益菌也表现出抑制作用;只有丁酸和乳酸既能够有效促进有益菌的繁殖,又能较有效地抑制有害菌.
表 4 不同种类有机酸对仔猪空肠和回肠中厌氧培养微生物菌群的影响1)Table 4. The effects of organic acids on intestinal bacteria profiles in vitro anaerobic culture system
2.3 有机酸调控仔猪全食糜培养体系微生物菌群的剂量效应
为进一步研究有机酸调控猪肠道微生物菌群的剂量效应,试验选择甲酸、乙酸、丁酸、乳酸和柠檬酸,分别设置6个浓度梯度进行试验.试验结果如图 1所示,甲酸、丁酸和柠檬酸能呈现剂量依赖性地抑制细菌增殖,而乙酸对总菌数量影响不大(图 1A).对益生菌而言,柠檬酸能显著抑制乳杆菌属和双歧杆菌属的增殖,但低剂量的乳酸(10 mmol·L-1)能显著促进乳杆菌属和双歧杆菌属的增殖(图 1B、1C),且低剂量的乙酸(15 ~ 20 mmol·L-1)也能促进双歧杆菌属的增殖(图 1C).所选择的5种有机酸在浓度达到10 ~ 15 mmol·L-1时,均能有效抑制病原菌(沙门菌和大肠埃希菌)的增殖(图 1D、1E).因此,体外厌氧培养条件下使用单一有机酸时:乙酸应使用约15 mmol·L-1,丁酸约15 mmol · L-1,乳酸约10 ~ 15 mmol·L-1,柠檬酸只能使用约5 mmol·L-1.
![图 1 不同浓度有机酸对断奶仔猪全食糜培养体系微生物菌群的影响]() 图 1 不同浓度有机酸对断奶仔猪全食糜培养体系微生物菌群的影响图中柱上*表示与对照组差异显著(P<0.05,t检验).Figure 1. The effects of different levels of concentration on organic acids on intestinal bacteria profiles in vitro total chyme anaerobic culture system
图 1 不同浓度有机酸对断奶仔猪全食糜培养体系微生物菌群的影响图中柱上*表示与对照组差异显著(P<0.05,t检验).Figure 1. The effects of different levels of concentration on organic acids on intestinal bacteria profiles in vitro total chyme anaerobic culture system综上所述,体外厌氧培养条件下使用单一有机酸调节微生物菌群,15 mmol · L-1乙酸和10 ~ 15 mmol·L-1乳酸是较好的选择.
3. 讨论与结论
3.1 不同有机酸对肠道有害菌的影响
多数研究显示,有机酸在抑制肠道病原菌(大肠埃希菌、沙门菌)方面作用显著[10-12],本研究待筛选的10种酸所用浓度为30 mmol·L-1,换算成质量分数后结果与部分文献的结果也基本一致[6, 13],但关于有机酸作用效果的报道仍存在差异,主要原因包括有机酸种类的不同、添加剂量存在差异.有学者认为有机酸抑制病原菌的机制主要有2个方面,首先是有机酸电离产生的氢离子,可以通过降低消化道pH间接减少细菌数量;其次,有机酸在某些复杂的酶作用下直接进入细菌的细胞从而抑制革兰阴性菌[14-15].现有的有机酸中,已有研究表明富马酸、柠檬酸、苹果酸和乳酸主要通过改变pH来抑菌,而甲酸、乙酸、丙酸和山梨酸等则可以进入细菌内,从而杀灭或抑制细菌[16].本文中的甲酸和乙酸等对大肠埃希菌和沙门菌的抑菌活性明显高于乳酸和富马酸等,说明甲酸和乙酸等直接添加在体外培养食糜中,可能同时通过两方面不同的机制发挥抑菌作用,因而表现出更强的抑菌活性.此外,有机酸电离氢离子的部位对其抑菌活性也有不同的影响.Gauthier等[17]则将肠道细菌分为pH敏感型(大肠埃希菌等)和pH不敏感型(乳酸杆菌和双歧杆菌).研究表明,未解离状态的有机酸能够穿透某些细菌的细胞膜,在细菌体内发生解离,释放出氢离子和阴离子.细胞内pH降低,无法耐受细胞内外pH变化的pH敏感型细菌则会由于消耗大量能量用来维持正常pH而被杀灭[18].由此可以推测,由于柠檬酸的pKa值较低,将保留更多的未解离状态,从而可能产生更强的抑菌活性.
3.2 不同有机酸对仔猪肠道有益菌的影响
本文研究结果发现,短链脂肪酸和乳酸能显著提高乳杆菌属数量,而丁酸、乳酸和山梨酸则对双歧杆菌属增殖有明显的促进效应.陈宝江等[19]通过试验也发现,添加酸化剂后,回肠中乳酸杆菌、厌氧菌数量呈上升趋势,且随着添加剂量增加,乳酸杆菌随之上升.而且多数报道也指出,添加单一有机酸或者复合酸化剂后,肠道内乳酸杆菌数量呈上升趋势[20-21],这主要是由于肠道pH降低,抑制有害微生物的活动和繁殖,从而促使有益菌繁殖[22],由此可见,不同种类和配比的酸化剂对仔猪肠道微生物区系的影响也不尽相同.
目前对有机酸的研究大多是通过动物试验的方式来进行,添加的有机酸也都通过不同的工艺方式进行加工处理,常用的酸化剂的加工工艺有吸附混合、流化床包衣和脂埋等技术,而随着工艺的改进,酸化剂的使用效果将不受饲料因素影响,且缓释性能得到极大的提高,从而导致不同研究中的添加剂量差异很大[23].本文通过体外研究模型,筛选出每种酸的最佳处理浓度,以期为生产实践中添加剂量的选择提供参考.而生产实践中,通过合理设置酸化剂的组合是增加肠道有益菌,降低病原菌,改善肠道健康的有效途径.
3.3 结论
综上所述,本试验采用仔猪食糜离体厌氧培养体系,研究各种有机酸对肠道微生物菌群的影响,并初步筛选确定了体外厌氧培养条件下使用单一有机酸调节微生物菌群,15 mmol · L-1乙酸和10 ~ 15 mmol·L-1乳酸是较好的选择.研究结果为进一步优化仔猪用有机酸制剂的配比提供了直接的试验依据.
-
![]()
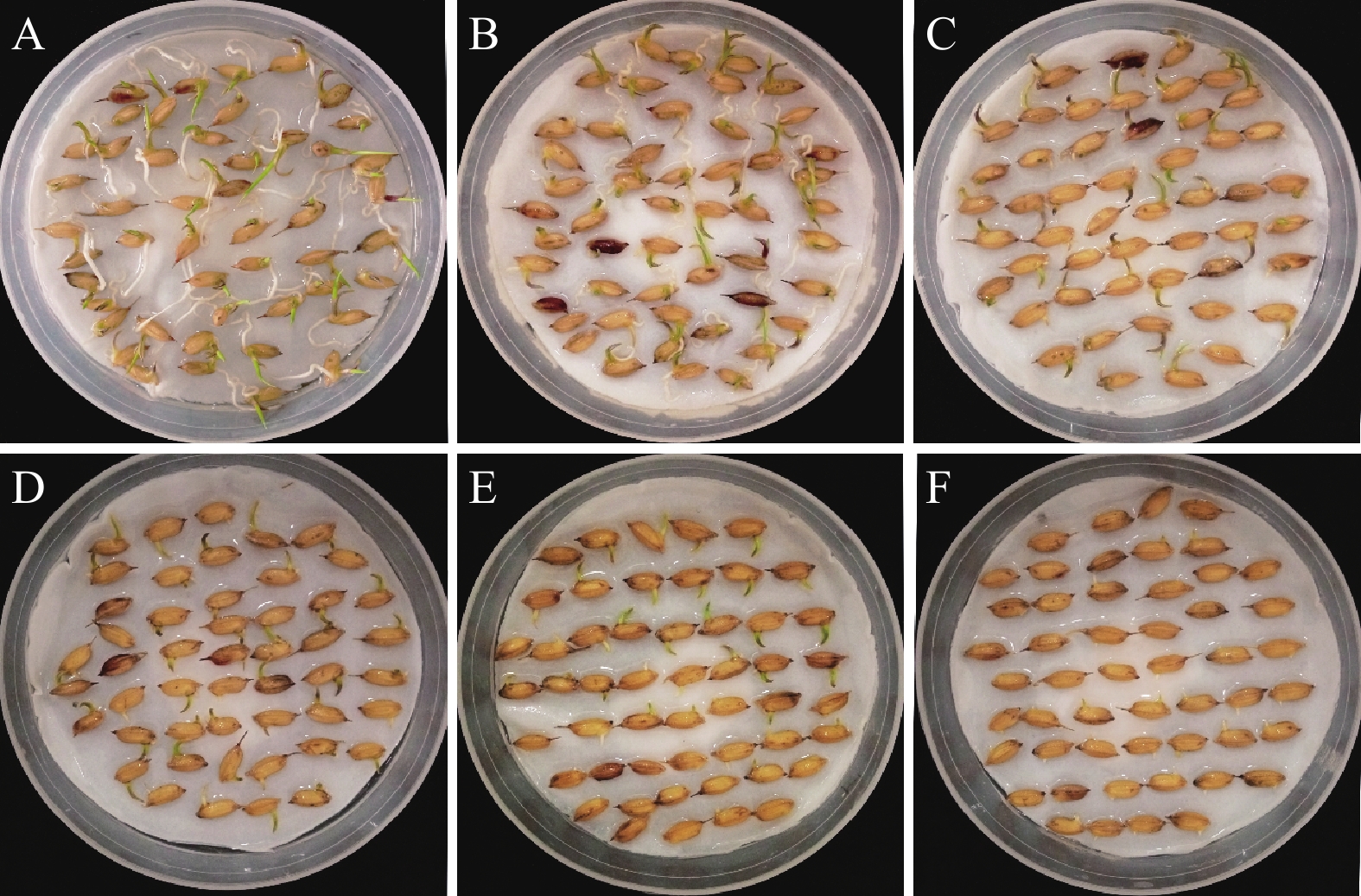
图 1 ‘华多1号’种子在不同浓度NaCl处理至7 d后的萌发情况比较
Figure 1. Comparison of seed germination of ‘Huaduo1’ after seven days under different NaCl concentrations
A~F:0、50、100、150、200、250 mmol/L NaCl
![]()
图 2 ‘华多1号’种子在不同浓度NaCl处理下的萌发特性
各小图中,柱子上方的不同小写字母表示差异显著(P<0.05,LSD法)
Figure 2. Germination characteristics of ‘Huaduo1’ under different NaCl concentrations
In each figure, different lowercase letters on bars indicate significant differences (P<0.05, LSD method)
![]()
图 3 ‘华多1号’幼苗在不同浓度NaCl处理下的根部生长情况
各小图中,柱子上方的不同小写字母表示差异显著(P<0.05,LSD法)
Figure 3. Root growth statistics of ‘Huaduo1’ seedlings under different NaCl concentrations
In each figure, different lowercase letters on bars indicate significant differences (P<0.05, LSD method)
![]()
图 4 ‘华多1号’在不同浓度NaCl处理下的幼苗长度及总鲜质量的变化情况
各小图中,柱子上方的不同小写字母表示差异显著(P<0.05,LSD法)
Figure 4. Changes of ‘Huaduo1’ seedling length and total fresh weight under different NaCl concentrations
In each figure, different lowercase letters on bars indicate significant differences (P<0.05, LSD method)
![]()
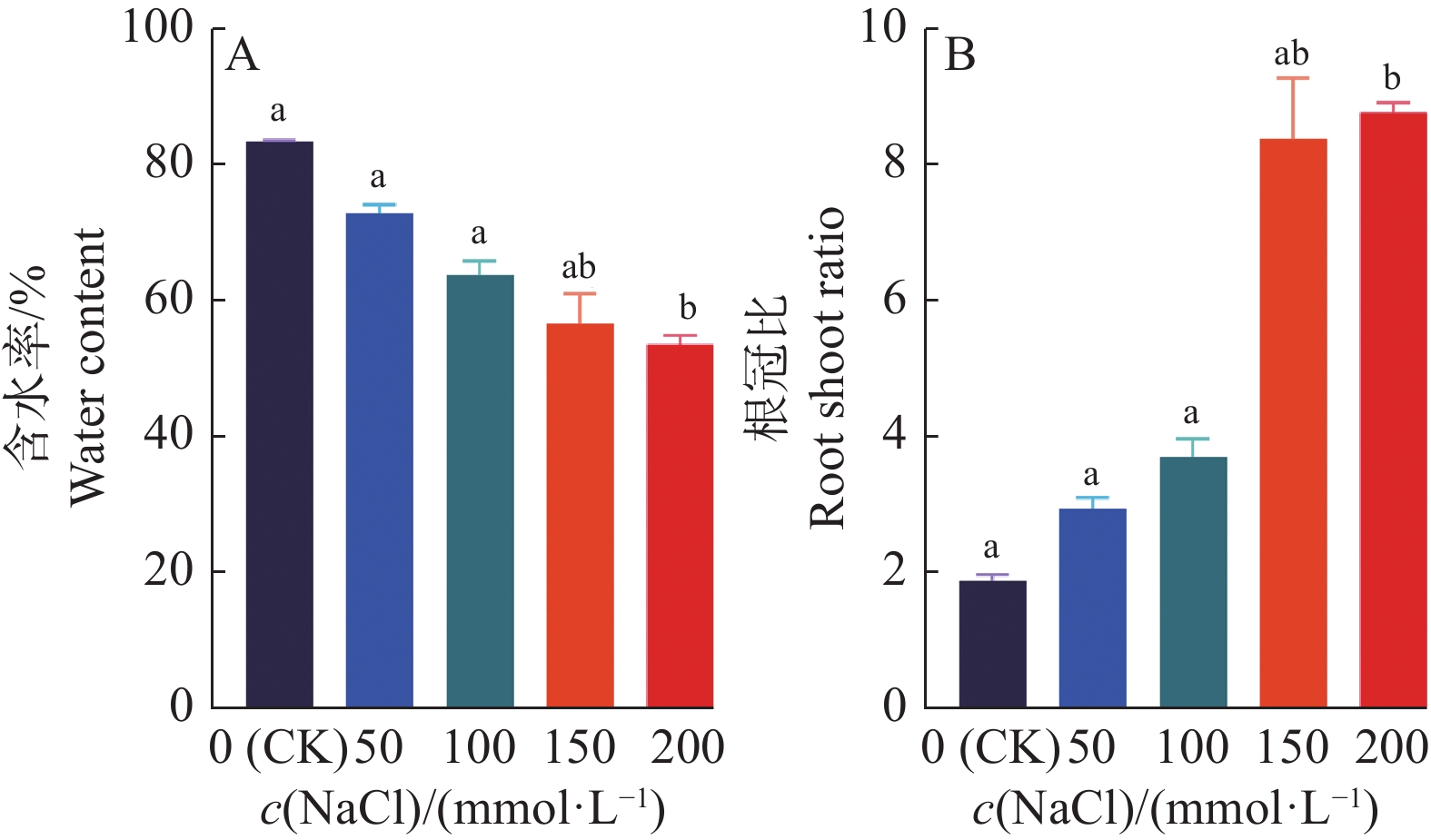
图 5 ‘华多1号’幼苗在不同浓度NaCl处理下的含水率及根冠比变化情况
各小图中,柱子上方的不同小写字母表示差异显著(P<0.05,LSD法)
Figure 5. Changes of water content and root/shoot ratio in ‘Huaduo1’ seedlings under different NaCl concentrations
In each figure, different lowercase letters on bars indicate significant differences (P<0.05, LSD method)
![]()
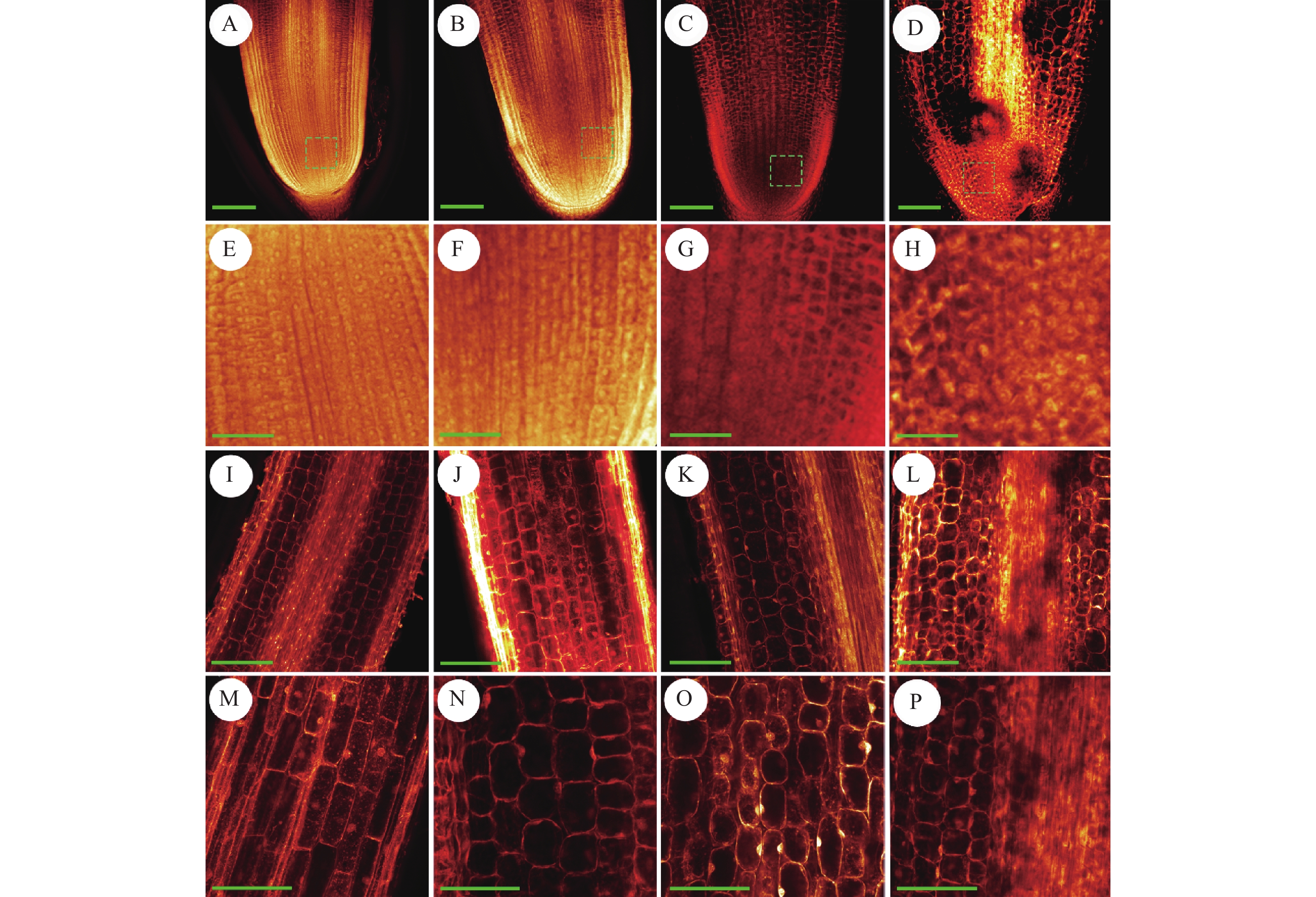
图 6 不同浓度NaCl处理下根尖细胞的形态差异
A~D分别为0、50、100和150 mmol/L的NaCl溶液处理后的根尖细胞,标尺= 20 µm;E~H分别为A~D对应的分生区细胞,标尺= 40 µm;I~L分别为A~D对应的伸长区细胞,标尺= 40 µm;M~P分别为A~D对应的伸长区细胞,标尺= 100 µm
Figure 6. Morphological differences of root tip cells under different NaCl concentrations
A−D: Root tip cells under NaCl concentration of 0, 50, 100, and 150 mmol/L, respectively, bar = 20 μm; E−H: Root tip meristematic zone cells corresponding to A−D respectively, bar = 40 μm; I−L: Root tip elongation zone cells corresponding to A−D respectively, bar =40 μm; M−P: Root tip elongation zone cells corresponding to A−D respectively, bar =100 μm
![]()
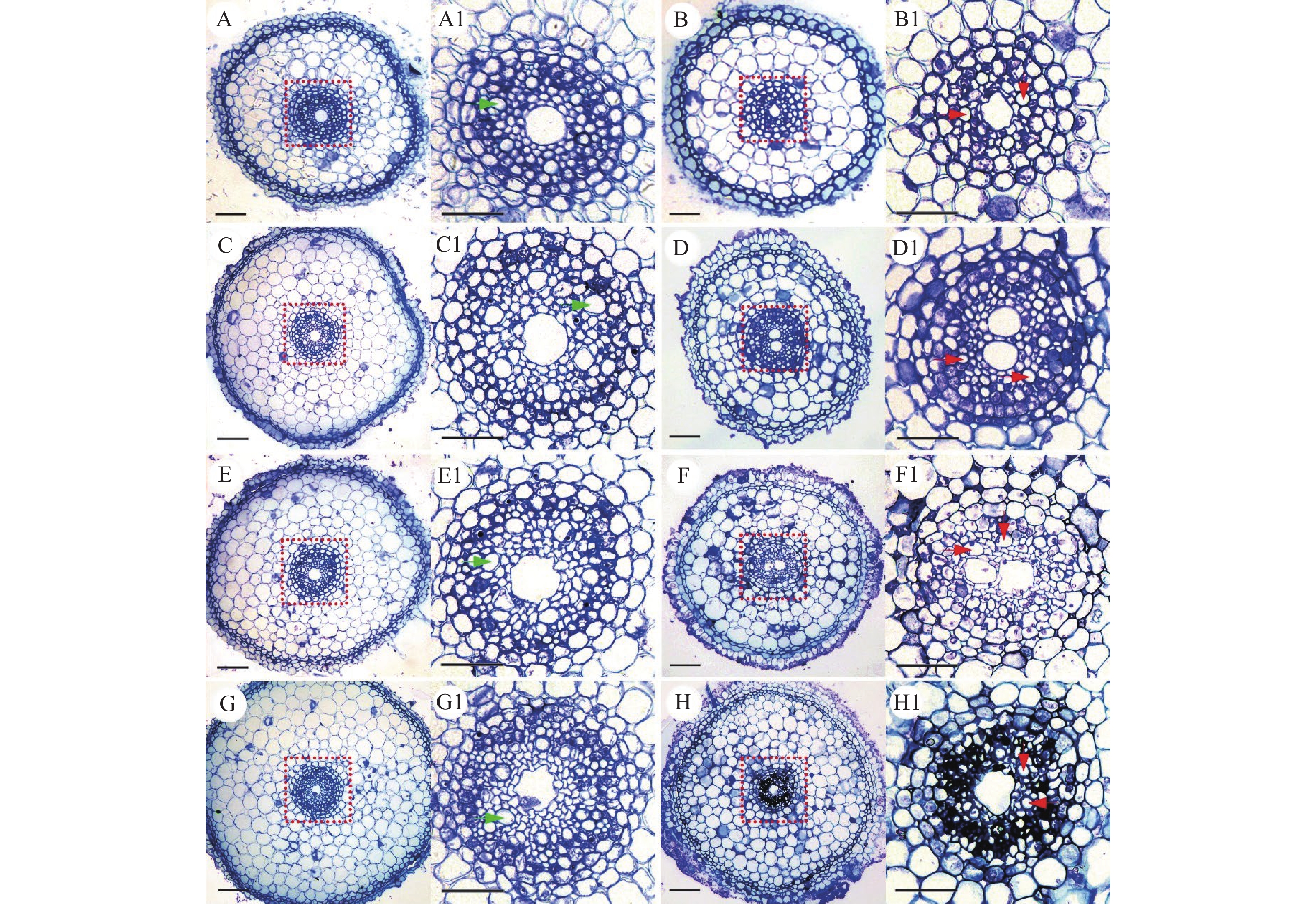
图 7 不同浓度NaCl处理下根尖伸长区横切面差异
A、B分别表示0 mmol/L NaCl溶液中处理8和20 d;C、D分别表示在50 mmol/L NaCl溶液中处理8和20 d;E、F分别表示100 mmol/L NaCl溶液中处理8和20 d;G、H分别表示150 mmol/L NaCl溶液中处理8和20 d;A1~H1:分别对应图A~H的根尖伸长区中柱结构细节;绿色和红色箭头分别表示盐胁迫处理下8和20 d后的根尖伸长区木质部;A~H:标尺=100 μm,A1-H1:标尺=20 μm
Figure 7. Transection of root tip elongation zone under different NaCl concentrations
A, B: Transection of root tip elongation zone under the 0 mmol/L NaCl treatment after the 8th day and 20th day; C, D: Transection of root tip elongation zone under the 50 mmol/L NaCl treatment afterthe 8th day and 20th day; E, F Transection of root tip elongation zone under the 100 mmol/L NaCl treatment after the 8th day and 20th day; G, H: Transection of root tip elongation zone under the 150 mmol/L NaCl treatment after the 8th day and 20th day; A1−H1: Detailed diagram of the column structure in the root tip elongation zone corresponding to figure A−H; The green arrow indicates the xylem of the root tip elongation zone under salt stress treatment after the 8th day, and the red arrow indicates the xylem of the root tip elongation zone under salt stress treatment after the 20th day; Bar = 100 μm in figure A−H, bar = 20 μm in figure A1−H1
![]()
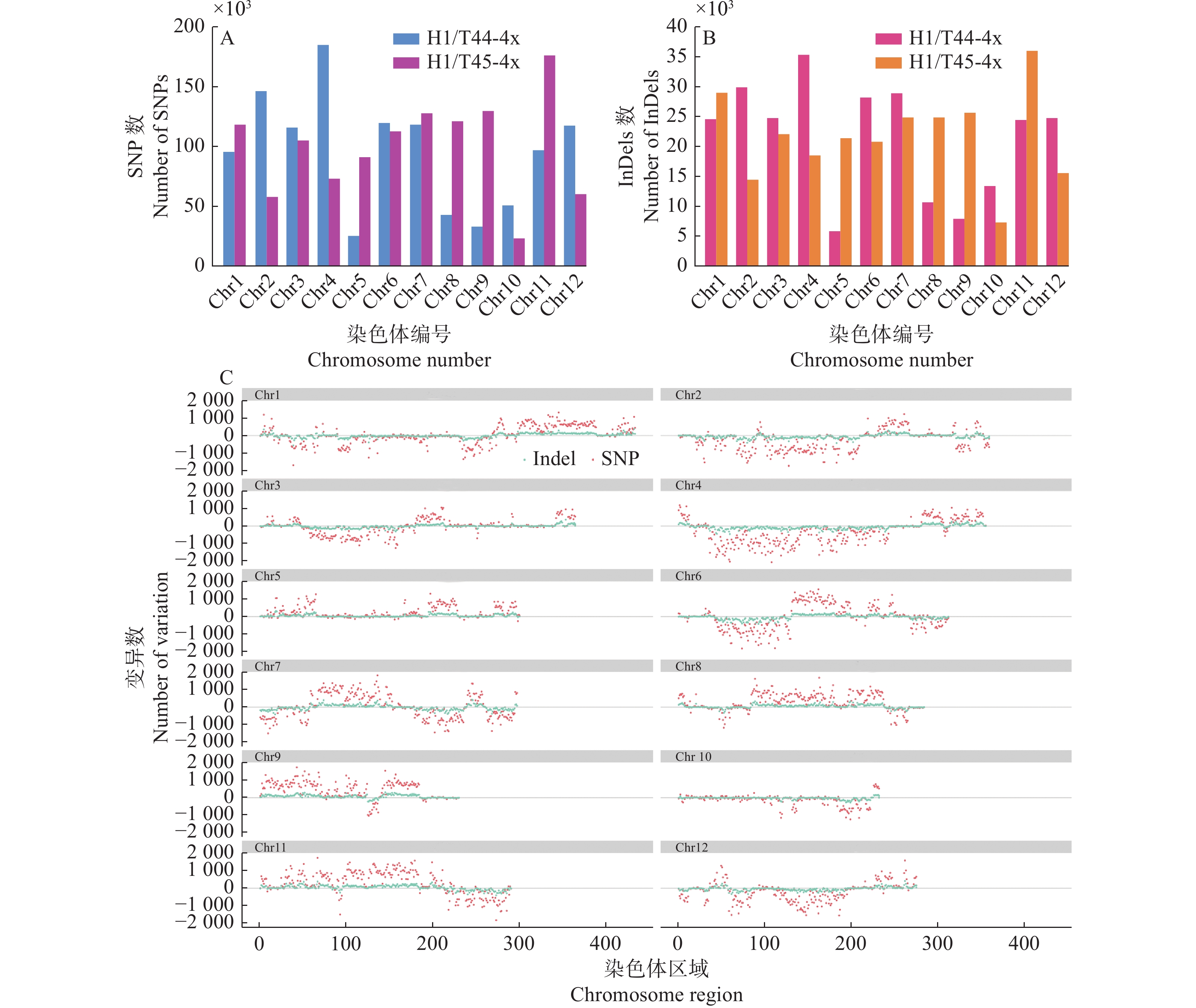
图 8 ‘华多1号’及双亲在各染色体的基因组变异情况
图C中,‘华多1号’分别与T44-4x和T45-4x的SNPs(粉红色)和InDels(浅绿色)的差值显示在纵轴上,正数表示H1和T44-4x之间的变化,负数表示H1和T45-4x之间的差异
Figure 8. Genomic variation of ‘Huaduo1’ and its parents on each chromosome
In figure C, the difference of SNPs (pink) and InDels (light green) in ‘Huaduo1’ is listed on the vertical axis, compared with T44-4x and T45-4x, respectively, the positive numbers represent the change between ‘Huaduo1’ and T44-4x, and the negative numbers represent the differences between ‘Huaduo1’ and T45-4x
![]()
图 9 ‘华多1号’中基因组变异基因的GO富集分析
Figure 9. GO enrichment analysis of genomic variant genes in ‘Huaduo1’
![]()
图 10 ‘华多1号’中盐胁迫相关基因的差异比较
“*”和“**”分别表示处理间存在0.05和0.01水平的显著差异(t检验)
Figure 10. Expression comparison of salt stress related genes in ‘Huaduo1’
“*” and “**” indicate significant differences between treatments at the levels of 0.05 and 0.01, respectively(t test)
表 1 盐胁迫相关基因的qRT-PCR引物序列
Table 1 Primer sequence for qRT-PCR of salt stress related genes
基因名称
Gene name引物序列 (5'→3')
Primer sequenceLOC_Os03g16900 F: ATAATGAAGCATCCGATT R: ATATGAGCAACAGAACAC LOC_Os03g20090 F: ACAACGAGATCAAGAACTACTG R: TTGAACTGCTGGCTGTTG LOC_Os11g26790 F: TCCAGCTCAAGCTCGTCT R: AGCTTCTCCTTGATCTTCTCCTT LOC_Os04g32920 F: TCCTCTCCCTCATCCTCT R: CATTGTCGTTGGCGTAGA LOC_Os02g34810 F: GCTTATCCAACCAATCAA R: ACATCAACTCGTCCATAT LOC_Os06g48510 F: ACACCTCGTTGCCTCATA R: ATAGCCACCATCCAGATAGTT Ubiquition F: ACCACTTCGACCGCCACTACT R: ACGCCTAAGCCTGCTGGTT  下载: 导出CSV
下载: 导出CSV
表 2 不同浓度NaCl胁迫对‘华多1号’种子发芽时间的影响1)
Table 2 Effects of salt stress with different NaCl concentration on seed germination time of ‘Huaduo1’
c(NaCl)/(mmol·L−1) 发芽起始时间/d
Germination initiation time发芽终止时间/d
Germination termination time平均发芽时间/d
Average germination time0 4 5 4.25±0.030a 50 4 5 4.36±0.009a 100 4 6 4.77±0.113b 150 4 7 4.97±0.154b 200 4 7 5.19±0.182c 250 5 8 6.81±0.409d 1)同列数据后不同小写字母表示差异显著(P<0.05,LSD法)
1)Different lowercase letters in the same column indicate significant differences (P<0.05, LSD method )
下载: 导出CSV
-
[1] 蔡得田, 袁隆平, 卢兴桂. 二十一世纪水稻育种新战略Ⅱ: 利用远缘杂交和多倍体双重优势进行超级稻育种[J]. 作物学报, 2001, 27(1): 110-116. [2] WU J W, HU C Y, SHAHID M Q, et al. Analysis on genetic diversification and heterosis in autotetraploid rice[J]. SpringerPlus, 2013, 2: 439. doi: 10.1186/2193-1801-2-439
[3] YANG P M, HUANG, Q C, QIN, G Y, et al. Different drought-stress responses in photosynthesis and reactive oxygen metabolism between autotetraploid and diploid rice[J]. Photosynthetica, 2014, 52: 193-202. doi: 10.1007/s11099-014-0020-2
[4] 阮华强, 黄群策, 杨鹏鸣. 同源四倍体水稻光合特性初步研究[J]. 杂交水稻, 2016, 31(2): 76-80. [5] 宋文昌, 张玉华. 水稻四倍化及其对农艺性状和营养成分的影响[J]. 作物学报, 1992, 18(2): 137-144. [6] XIAN L, LONG Y, YANG M, et al. iTRAQ-based quantitative glutelin proteomic analysis reveals differentially expressed proteins in the physiological metabolism process during endosperm development and their impacts on yield and quality in autotetraploid rice[J]. Plant Science, 2021, 306: 110859. doi: 10.1016/j.plantsci.2021.110859
[7] GUO H B, MENDRIKAHY J N, XIE L, et al. Transcriptome analysis of neo-tetraploid rice reveals specific differential gene expressions associated with fertility and heterosis[J]. Scientific Reports, 2017, 7: 40139. doi: 10.1038/srep40139
[8] BEI X, SHAHID M Q, WU J, et al. Re-sequencing and transcriptome analysis reveal rich DNA variations and differential expressions of fertility-related genes in neo-tetraploid rice[J]. PLoS One, 2019, 14(4): e0214953. doi: 10.1371/journal.pone.0214953
[9] GHALEB M A A, LI C, SHAHID M Q, et al. Heterosis analysis and underlying molecular regulatory mechanism in a wide-compatible neo-tetraploid rice line with long panicles[J]. BMC Plant, 2020, 20: 83. doi: 10.1186/s12870-020-2291-z
[10] CHEN L, YUAN Y, WU J, et al. Carbohydrate metabolism and fertility related genes high expression levels promote heterosis in autotetraploid rice harboring double neutral genes[J]. Rice, 2019, 12(1): 34. doi: 10.1186/s12284-019-0294-x
[11] YU H, SHAHID M Q, LI Q, et al. Production assessment and genome comparison revealed high yield potential and novel specific alleles associated with fertility and yield in neo-tetraploid rice[J]. Rice, 2020, 13(1): 32. doi: 10.1186/s12284-020-00387-3
[12] YU H, LI Q, LI Y, et al. Genomics analyses reveal unique classification, population structure and novel allele of neo-tetraploid rice[J]. Rice, 2021, 14(1): 16. doi: 10.1186/s12284-021-00459-y
[13] HUANG Y, HUANG Q, LI J, et al. Photosynthetic physiology and molecular response mechanisms of indica-japonica intersubspecific tetraploid rice seedlings to ion beams[J]. Journal of Plant Growth Regulation, 2021, 40(2): 722-735. doi: 10.1007/s00344-020-10136-x
[14] TU Y, JIANG A, GAN L, et al. Genome duplication improves rice root resistance to salt stress[J]. Rice, 2014, 7(1): 15. doi: 10.1186/s12284-014-0015-4
[15] 刘向东, 吴锦文, 陆紫君, 等. 同源四倍体水稻: 低育性机理、改良与育种展望[J]. 遗传, 2023. doi: 10.16288/j.yczz.23-074. [16] 汪宗立, 李建坤, 王志霞. 水稻耐盐性的生理研究 IV. 盐渍对超氧物歧化酶和过氧化氢酶活性的影响[J]. 江苏农业学报, 1990, 6(2): 1-6. [17] 顾兴友, 郑少玲, 严小龙, 等. 盐浓度对水稻苗期耐盐指标变异度的影响[J]. 华南农业大学学报[J], 1998, 19(1): 33-37. [18] ZENG Y X, HU C Y, LU Y G, et al. Diversity of abnormal embryo sacs in indica/japonica hybrids in rice demonstrated by confocal microscopy of ovaries[J]. Plant Breeding, 2010, 126: 574-580.
[19] WU J W, CHEN Y, LIN H, et al. Comparative cytological and transcriptome analysis revealed the normal pollen development process and up-regulation of fertility-related genes in newly developed tetraploid rice[J]. International of Molecular Science, 2020, 21(19): 7046. doi: 10.3390/ijms21197046
[20] WU J W, SHAHID M Q, GUO H B, et al. Comparative cytological and transcriptomic analysis of pollen development in autotetraploid and diploid rice[J]. Plant Reproduction, 2014, 27(4): 181-196. doi: 10.1007/s00497-014-0250-2
[21] LIVAK K J, SCHMITTGEN T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method[J]. Methods, 2001, 25(4): 402-408.
[22] COMAI L. The advantages and disadvantages of being polyploid[J]. Nature Reviews Genetics, 2005, 6(11): 836-846. doi: 10.1038/nrg1711
[23] VAN DE PEER Y, MIZRACHI E, MARCHAL K. The evolutionary significance of polyploidy[J]. Nature Reviews Genetics, 2017, 18(7): 411-424. doi: 10.1038/nrg.2017.26
[24] WANG L, CAO S, WANG P, et al. DNA hypomethylation in tetraploid rice potentiates stress- responsive gene expression for salt tolerance[J]. Proceedings of the National Academy of Sciences, 2021, 118(13): e2023981118. doi: 10.1073/pnas.2023981118
[25] 刘向东, 吴锦文, SHAHID M Q. 新型四倍体水稻创制及其杂种优势利用研究进展[J]. 生物技术通报, 2021, 38(1): 44-50. [26] 钟静, 陈大鹏. NaCl胁迫对水稻种子萌发和幼苗生长的影响[J]. 黑龙江农业科学, 2016, 5: 18-20. [27] 贺奇, 杨锋, 王昕, 等. NaCl胁迫对水稻宁粳48号种子萌发特性的影响[J]. 宁夏农林科技, 2017, 58(3): 4-6. [28] 信彩云, 马惠, 赵庆雷, 等. 不同浓度NaCl胁迫对水稻种子发芽及幼苗生长的影响[J]. 大麦与谷类科学, 2019, 36(3): 7-10. [29] 刘胜群, 宋凤斌. 不同耐旱性玉米根系解剖结构比较研究[J]. 干旱地区农业研究, 2007, 2: 86-91. [30] GANGULY M, DATTA K, ROYCHOUDHURY A, et al. Overexpression of Rab16A gene in indica rice variety for generating enhanced salt tolerance[J]. Plant Signaling and Behavior, 2012, 7(4): 502-509. doi: 10.4161/psb.19646
[31] YANG A, DAI X Y, ZHANG W H. A R2R3-type MYB gene, OsMYB2, is involved in salt, cold, and dehydration tolerance in rice[J]. Journal of Experimental Botany, 2012, 63(7): 2541-2556. doi: 10.1093/jxb/err431