
Interspecific relationships between Rhopalosiphum rufiabdominalis and Sogatella furcifera mediated by rice secondary metabolites
-
摘要:目的
探究白背飞虱Sogatella furcifera与红腹缢管蚜Rhopalosiphum rufiabdominalis种间互作机制,以及2种昆虫互作过程中水稻次生代谢物质的作用。
方法将白背飞虱和红腹缢管蚜按不同比例混合饲养,待白背飞虱长至成虫后分组配对,分析白背飞虱总产卵量及日均产卵量;测定各处理的水稻草酸、黄酮及总酚含量,利用GC/MS仪器分析各处理水稻幼苗次生代谢物质成分的差异。
结果“15头蚜虫 + 5头白背飞虱”处理的白背飞虱总产卵量仅为131.67粒,与“20头白背飞虱”处理(214.60粒)差异显著,日均产卵量也呈现相同的规律;“15头蚜虫 + 5头白背飞虱”处理的水稻黄酮和总酚含量分别为1.98和63.71 mg/L,均显著高于其他处理。GC/MS分析表明,“15头蚜虫 + 5头白背飞虱”处理的水稻中丙丁酚相对含量高达75.78%,而其余处理的水稻中不存在此种酚类物质。
结论红腹缢管蚜可能是通过刺激水稻提高黄酮及总酚含量,进而抑制白背飞虱的生殖能力,其中丙丁酚可能在白背飞虱和红腹缢管蚜种间互作中起着关键作用。
Abstract:ObjectiveTo explore the interaction mechanism between species of Sogatella furcifera (white-backed planthopper) and Rhopalosiphum rufiabdominalis (aphid), and the role of secondary metabolites in rice during the interaction between the two insects.
MethodS. furcifera and R. rufiabdominalis were mixed in different proportions, and the total egg production and average daily spawning of white-backed planthopper adults were analyzed after pairing. The contents of oxalic acid, flavones and total phenols of each treated rice seedlings were determined, and the differences in compositions of secondary metabolites were analyzed by GC/MS instrument.
ResultThe total egg production of white-backed planthoppers treated with “15 aphids + 5 white-backed planthoppers” was only 131.67 grains, which was significantly different from that of “20 white-backed planthoppers” (214.60 grains), and the average daily fecundity was the same. The contents of rice flavones and total phenols treated with “15 aphids + 5 white-backed planthoppers” were 1.98 and 63.71 mg/L, respectively, which were significantly higher than those of other treatments. GC/MS analysis showed that the relative content of probucol in rice treated with “15 aphids + 5 white-backed planthoppers” was as high as 75.78%, while this phenolic substance was not present in the rest of the treatments.
ConclusionR. rufiabdominalis may inhibit the reproductive ability of S. furcifera through stimulating rice to promote the content of flavones and total phenols. The probucol may play a key role in the interaction between S. furcifera and R. rufiabdominalis.
-
在昆虫纲中,昆虫互作的现象频繁发生,其互作关系的类型包括中性、竞争、偏害、捕食、寄生、偏利及互利等[1]。其中,捕食、寄生和竞争是目前较为普遍的类型。例如,邱良妙等[2]研究发现六斑月瓢虫Menochilus sexmaculata 4龄幼虫及成虫对低龄的草地贪夜蛾Spodoptera frugiperda均有较好的防治潜能。张晓明等[3]研究发现海氏浆角蚜小蜂Eretmocerus hayati和浅黄恩蚜小蜂Encarsia sophia均能寄生于烟粉虱Bemisia tabaci体内,两者在3︰1比例数量组合下对烟粉虱的寄生率最高。Kaplan等[4]总结了333种植食性昆虫的种间关系,其中62%属于竞争关系,因此竞争关系在昆虫互作中占有重要地位。刘晓飞等[5]比较了桔小实蝇Bactrocera (Bactrocera) dorsalis及番石榴实蝇Bactrocera (Bactrocera) correcta幼虫在不同温度下存活率的差异,结果表明在33 ℃条件下番石榴实蝇存活率更高,说明番石榴实蝇具有较强的竞争优势。禹云超等[6]发现,西花蓟马Frankliniella occidentalis作为入侵性害虫,与本地的东方花蓟马Frankliniella tritici存在种间竞争关系,东方花蓟马能显著降低西花蓟马若虫的存活率,说明东方花蓟马具有竞争优势。
昆虫间的互作受到其自身和环境因素的影响,有研究表明,许多食草昆虫之间的相互作用与寄主植物相关[7]。例如,李建领[8]研究发现,枸杞瘿螨Aceria pallida通过取食诱导枸杞产生防御酶及类黄酮、总酚等次生物质,进而抑制枸杞木虱Bactericera gobica的生长发育。Zhao等[9]亦研究发现烟粉虱取食烟草,从而促进烟草水杨酸含量的增高,进而提高桃蚜Myzus persicae死亡率,显著降低桃蚜的繁殖力。寄主植物为抵抗昆虫的危害,在危害过程中被诱导产生防御性次生代谢物质,如黄酮类、酚类、有机酸、生物碱等化合物,极大影响了昆虫的取食、生长发育等行为[10-11],以上研究均证明了植物次生代谢物质能够间接参与昆虫的种间互作行为。我们前期研究表明,白背飞虱Sogatella furcifera和红腹缢管蚜Rhopalosiphum rufiabdominalis存在种间互作,将两者混养后,白背飞虱后代数量显著减少。为明确红腹缢管蚜和白背飞虱种间互作的原理,同时验证与植物次生代谢物质的关联性,本研究将红腹缢管蚜和白背飞虱按不同比例混合饲养,探究红腹缢管蚜抑制白背飞虱繁殖的最佳有效比例,以及红腹缢管蚜刺激水稻产生的次生代谢物质反作用抑制白背飞虱生殖的机理,为科学合理利用昆虫互作控制红腹缢管蚜和白背飞虱提供理论依据。
1. 材料与方法
1.1 供试材料
1.1.1 供试水稻
选用四川农业大学水稻研究所提供的TN1水稻品种,种子用水冲洗,浸泡2 d后,置于恒温箱(30±1) ℃中培养备用。
1.1.2 供试虫源
由华中农业大学提供白背飞虱敏感品系,于养虫笼内(45 cm × 35 cm × 22 cm)以TN1水稻繁育。饲养条件:温度(26±1) ℃,相对湿度70%~80%,光周期14 h光照︰10 h黑暗。本试验所需的白背飞虱为3龄若虫。
由四川农业大学无公害农药实验室提供红腹缢管蚜,于透明塑料盒内(33 cm × 18 cm × 14 cm)以TN1水稻繁育。饲养条件:温度(26±1) ℃,相对湿度70%~80%,光周期14 h光照︰10 h黑暗。本试验所需的红腹缢管蚜为3~4龄若虫。
1.1.3 主要设备和仪器
三重四级杆气质联用仪GC-MS-TQ8040(岛津);UV-3000紫外可见分光光度计(上海美谱达仪器有限公司);D37520离心机(美国科峻仪器公司);RXZ型智能人工气候箱(宁波江南仪器厂)。
1.2 试验方法
1.2.1 水稻前期处理
用φ为5%的过氧化氢水溶液浸泡TN1水稻种子2 d,催芽露白后,将种子播种于底部垫有营养液棉花的500 mL塑料杯内,每杯种子15粒,然后置于(26±1) ℃、湿度70%~80%的培养箱内,期间补充适量营养液。待幼苗长至5~6 cm备用。
1.2.2 白背飞虱与红腹缢管蚜互作装置
挑选出3~4龄的无翅红腹缢管蚜和处于3龄期的白背飞虱备用。试验处理见表1,仅有水稻的处理为对照(CK)。培养箱温度控制在(26±1) ℃,相对湿度70%~80%,每天光照14 h,每个处理重复10次。
表 1 各试验处理的白背飞虱和红腹缢管蚜初始数量Table 1. Initial number of white-backed planthopper and aphid in each experimental treatment处理
Treatment蚜虫/头
Aphid白背飞虱/头
White-backed planthopperT1 20 0 T2 15 5 T3 10 10 T4 5 15 T5 0 20 T6(CK) 0 0 1.2.3 白背飞虱产卵量的测定
待上述处理的白背飞虱发育至5龄若虫,准备2株新鲜的水稻苗并转移至玻璃试管内(直径2.0 cm,高度14.5 cm)备用。选取同一天羽化的雄/雌虫进行一一配对,吸取至上述玻璃试管内,然后每天将雌、雄成虫转移至新的试管中,并且定时向旧的稻苗中添加营养液以防枯黄坏死,所有配对的白背飞虱均在同一条件下培养。于10 d后记录试管中白背飞虱的孵化情况,使用镊子剖开整株稻苗统计未孵化的卵数,然后对比T2~T5这4个处理的总产卵量及日均产卵量,筛选出抑制白背飞虱产卵量的最佳配比[12]。每个处理重复15次。
1.3 水稻次生代谢物质含量的测定
待白背飞虱发育至成虫后,移除塑料杯内所有的白背飞虱和红腹缢管蚜,清理水稻叶片上残留的蜕皮及蜜露,选用整株水稻以测定水稻中的草酸、黄酮及总酚含量。
1.3.1 草酸含量的测定
样品制备:参考段立珍等[13]的方法,称取各处理水稻幼苗0.5 g,加入10 mL KCl-HCl缓冲液(pH 2.0),用石英砂研磨数分钟至匀浆,然后用15 mL超纯水清洗转入容量瓶中,在75~80 ℃水浴锅中加热震荡30 min,过滤装入离心管中并用超纯水定容至刻度线,即得到草酸待测液。
水稻草酸含量的测定:参考段立珍等[13]的方法,用样品草酸待测液代替草酸铵标准溶液,室温下静置30 min后在510 nm波长处测定光密度,代入标准曲线计算出水稻草酸含量。
1.3.2 黄酮含量的测定
样品制备:参考李明阳等[14]的方法,称取0.5 g水稻幼苗,在离心管中研磨至粉状,然后加入φ为40%的乙醇溶液10 mL,于40 ℃超声波清洗仪中提取45 min,期间超声波辅助作用15 min(20 kHz),取出后在7 000 r/min下离心5 min,取上清液为待测样品。
水稻黄酮含量的测定:参考李明阳等[14]的方法,用黄酮待测液代替芦丁标准溶液,在510 nm波长下测定光密度,根据芦丁标准曲线计算出水稻中黄酮含量。
1.3.3 总酚含量的测定
待测样品制备方法与“1.3.2”一致。
水稻总酚含量的测定:参考蔡文国等[15]的方法,用上述待测液代替没食子酸标准溶液,在760 nm波长下测定其光密度。测得的光密度代入标准曲线,计算各处理水稻总酚的含量。
1.4 GC/MS法测定水稻化学成分相对含量
取处理T2、T4及T6(CK)的整株稻苗测定水稻中化合物的成分及相对含量。样品处理[14,16]:称取0.1 g整株水稻样品研磨至粉状,装入5 mL离心管中,加入1.4 mL预冷后的甲醇,充分摇匀,在10700 r/min下离心10 min。转移上清液至10 mL离心管,加入0.75 mL三氯甲烷及1.4 mL超纯水(均预冷至–20 ℃)充分摇匀混合,在4800 r/min下离心10 min,然后将上清液转入1.5 mL离心管中备用。
GC/MS仪器程序设定:参照李明阳等[14]的方法。分析仪器:GC-MS-TQ8040,RTX-5MS毛细管柱(30 m × 0.25 mm × 0.25 µm)。以氦气为载体,柱流量为l mL/min。柱温从100 ℃(保持3 min)以5 ℃/min速度升温到280 ℃(保持5 min)。进样体积1 μL,配备一个AS-3000自动取样器,并设置用于采样无分离注射。注射温度250 ℃,轰击电压70 eV,质量扫描范围m/z为50~650 。记录各处理中水稻化学成分的保留时间(Retention time),在NIST98 MS数据库中对比分析并记录各成分的名称及其分子式,找出不同处理下相对含量变化明显的成分。
1.5 数据处理及分析
采用Excel及IBM SPSS Statistics 27软件整理分析数据,采用单因素方差分析(One-way ANOVA)法分析不同处理间白背飞虱总产卵量及水稻草酸、黄酮、总酚含量的差异性,处理间差异显著性采用Duncan’s法检验,作图使用SigmaPlot 10.0软件。
2. 结果与分析
2.1 红腹缢管蚜和白背飞虱不同比例混合对白背飞虱产卵量的影响
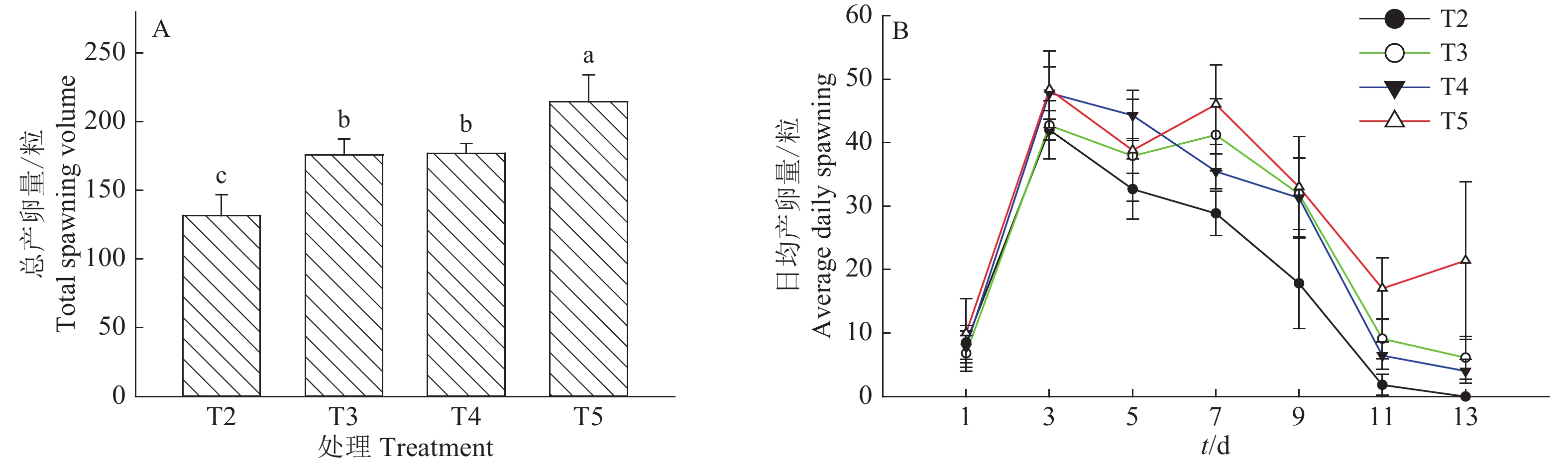
白背飞虱总产卵量见图1A,处理T5总产卵量高达214.60粒,处理T4(177.09粒)和处理T3(175.70粒)次之,处理T2最低,仅131.67粒,且处理间差异显著(P<0.05)(T3、T4间除外)。白背飞虱日均产卵量见图1B,第1天处理T2产卵量与其余3组近似相同,第3~13天每日产卵量均低于其余3组;前期,处理T3产卵量低于T4,但中后期与之相反,且中后期两组产卵量相差较小。处理T5除第5天产卵量稍低于T4以外,其余时间产卵量均高于其他处理,第3天高达48.40粒,且后期产卵量远超过其余处理。可见,红腹缢管蚜能够显著影响白背飞虱的产卵量,且初始红腹缢管蚜数量越多,对白背飞虱产卵量的抑制程度越明显。
![]() 图 1 各处理组白背飞虱的单雌总产卵量及日均产卵量T2:15头蚜虫 + 5头白背飞虱,T3:10头蚜虫 + 10头白背飞虱,T4:5头蚜虫 + 15头白背飞虱,T5:20头白背飞虱;图A中,柱子上方的不同小写字母表示处理间差异显著(P<0.05,Duncan’s法)Figure 1. Total egg production and average daily spawning of white-backed planthopper in each treatmentT2: 15 aphids + 5 white-backed planthoppers, T3: 10 aphids + 10 white-backed planthoppers, T4: 5 aphids + 15 white-backed planthoppers, T5: 20 white-backed planthoppers; In figure A, different lowercase letters above the bars indicate significant differences among treatments (P<0.05, Duncan’s method)
图 1 各处理组白背飞虱的单雌总产卵量及日均产卵量T2:15头蚜虫 + 5头白背飞虱,T3:10头蚜虫 + 10头白背飞虱,T4:5头蚜虫 + 15头白背飞虱,T5:20头白背飞虱;图A中,柱子上方的不同小写字母表示处理间差异显著(P<0.05,Duncan’s法)Figure 1. Total egg production and average daily spawning of white-backed planthopper in each treatmentT2: 15 aphids + 5 white-backed planthoppers, T3: 10 aphids + 10 white-backed planthoppers, T4: 5 aphids + 15 white-backed planthoppers, T5: 20 white-backed planthoppers; In figure A, different lowercase letters above the bars indicate significant differences among treatments (P<0.05, Duncan’s method)2.2 红腹缢管蚜和白背飞虱共存对水稻次生代谢物质含量的影响
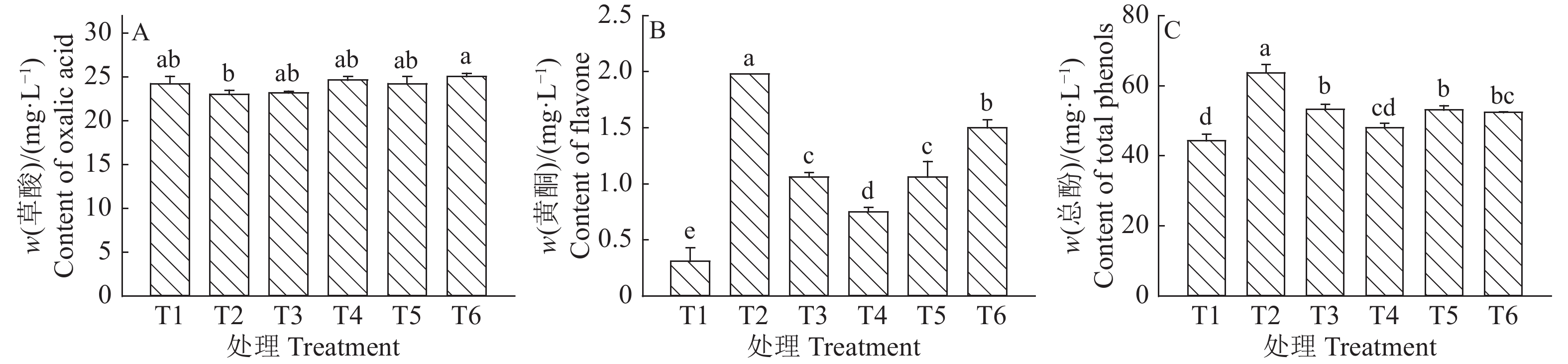
不同处理间水稻草酸含量的测定结果见图2A,处理T6(CK)的草酸含量最高,达到了25.06 mg/L,其余由高到低依次为T4(24.65 mg/L)、T1(24.23 mg/L)、T5(24.23 mg/L)和T3(23.19 mg/L),T2含量最低(23.03 mg/L),除T2以外,其余处理之间差异均不显著。可见,在T2处理下高初始量的红腹缢管蚜能够降低水稻草酸含量。
![]() 图 2 各处理组水稻的草酸、黄酮和总酚含量T1:20头蚜虫,T2:15头蚜虫 + 5头白背飞虱,T3:10头蚜虫 + 10头白背飞虱,T4:5头蚜虫 + 15头白背飞虱,T5:20头白背飞虱,T6(CK):0头蚜虫 + 0头白背飞虱;各图中,柱子上方的不同小写字母表示处理间差异显著(P<0.05,Duncan’s法)Figure 2. Contents of oxalic acid, flavones and total phenols in rice of each treatmentT1: 20 aphids, T2: 15 aphids + 5 white-backed planthoppers, T3: 10 aphids + 10 white-backed planthoppers, T4: 5 aphids + 15 white-backed planthoppers, T5: 20 white-backed planthoppers, T6 (Control treatment): 0 aphid + 0 white-backed planthopper; In each figure, different lowercase letters above the bars indicate significant differences among treatments (P<0.05, Duncan’s method)
图 2 各处理组水稻的草酸、黄酮和总酚含量T1:20头蚜虫,T2:15头蚜虫 + 5头白背飞虱,T3:10头蚜虫 + 10头白背飞虱,T4:5头蚜虫 + 15头白背飞虱,T5:20头白背飞虱,T6(CK):0头蚜虫 + 0头白背飞虱;各图中,柱子上方的不同小写字母表示处理间差异显著(P<0.05,Duncan’s法)Figure 2. Contents of oxalic acid, flavones and total phenols in rice of each treatmentT1: 20 aphids, T2: 15 aphids + 5 white-backed planthoppers, T3: 10 aphids + 10 white-backed planthoppers, T4: 5 aphids + 15 white-backed planthoppers, T5: 20 white-backed planthoppers, T6 (Control treatment): 0 aphid + 0 white-backed planthopper; In each figure, different lowercase letters above the bars indicate significant differences among treatments (P<0.05, Duncan’s method)不同处理间水稻黄酮含量的测定结果见图2B,处理T2的黄酮含量高达1.98 mg/L,其余由高到低依次为T6(CK)(1.50 mg/L)、T5(1.06 mg/L)、T3(1.06 mg/L)和T4(0.75 mg/L),T1最低(0.31 mg/L),除T3与T5之间差异不显著以外,其余处理之间均差异显著。因此,在红腹缢管蚜与白背飞虱共存的情况下,高初始量的红腹缢管蚜能够明显提高水稻黄酮含量,T2处理尤为显著。
不同处理间水稻总酚含量的测定结果见图2C,处理T2的总酚含量最高,达到63.71 mg/L,其余由高到低依次为T3(53.20 mg/L)、T5(53.10 mg/L)、T6(CK)(52.37 mg/L)、T4(48.01 mg/L),T1最低(44.29 mg/L),且处理T2、T3、T4间差异显著。因此,红腹缢管蚜与白背飞虱互作对水稻总酚含量影响显著,且高初始量的红腹缢管蚜能够明显提升水稻总酚含量,T2处理尤为显著。
2.3 GC/MS法分析水稻次生代谢物质成分
经过NIST98 MS数据库对比,发现不同处理间水稻幼苗次生代谢物质种类及相对含量存在较大的差别,共涉及18种化合物(表2)。处理T2中,丙丁酚相对含量最高(75.78%),其余较高的为2,4–二叔丁基苯酚(5.38%)、二苯基砜(4.21%);处理T4中,2,4–二叔丁基苯酚相对含量最高(28.59%),其余为二苯基砜(28.02%)、7,9–二叔丁基–1–氧杂螺(4,5)癸–6,9–二烯–2,8–二酮(15.95%);处理T6(CK)中,二苯基砜相对含量最高(23.68%),其余为5–羟甲基糠醛(16.67%)、2,3–二氢–3,5–二羟基–6–甲基–4H–吡喃–4–酮(13.21%)。其中,区别最明显的为处理T4及T6水稻中均无丙丁酚,而处理T2水稻中的丙丁酚相对含量高达75.78%,表明丙丁酚可能在白背飞虱与红腹缢管蚜互作中起着重要作用,且相对含量与红腹缢管蚜/白背飞虱的初始比可能有密切联系。另外,相对含量差别较大的还有2,3–二氢–3,5–二羟基–6–甲基–4H–吡喃–4–酮、5–羟甲基糠醛、2,4–二叔丁基苯酚、二苯基砜、7,9–二叔丁基–1–氧杂螺(4,5)癸–6,9–二烯–2,8–二酮等成分,这可能与两者互作有一定的关系。
表 2 各处理组水稻主要次生代谢物质成分及相对含量Table 2. The main secondary metabolite components and relative contents in rice of each treatment化合物名称
Compound name分子式
Molecular formulat保留/min
Retention time相对含量1)/% Relative content T2 T4 T6 2,3−二氢−3,5−二羟基−6−甲基−4H−吡喃−4−酮
2,3-Dihydro-3,5-dihydroxy-6-methyl-4H-pyran-4-oneC6H8O4 5.664 0.67 3.71 13.21 萘 Naphthalene C10H8 5.931 0.41 0.95 2.82 2,4−二甲基苯甲醛 2,4-Dimethyl-benzaldehyde C9H10O 6.662 1.53 8.38 3.45 3−乙酰氧基−3−羟基丙酸甲酯
3-Acetoxy-3-hydroxypropionic acid-methyl esterC6H10O5 7.800 0 0 6.39 1−硝基−1−脱氧−d−甘油−l−甘露庚醇
1-Nitro-1-deoxy-d-glycero-l-mannoheptitolC7H15NO8 8.943 0 0 1.87 十二烷醛 Dodecanal C12H24O 11.407 0.08 1.07 1.70 5−羟甲基糠醛 5-Hydroxymethylfurfural C6H6O3 12.881 0.35 0 16.67 2,4−二叔丁基苯酚 2,4-Di-tert-butylphenol C14H22O 14.001 5.38 28.59 2.70 7,9−二叔丁基−1−氧杂螺(4,5)癸−6,9−二烯−2,8−二酮
7,9-Di-tert-butyl-1-oxaspiro(4,5)deca-6,9-diene-2,8-dioneC17H24O3 22.629 2.79 15.95 2.23 二苯基砜 Diphenyl sulfone C12H10O2S 23.036 4.21 28.02 23.68 9−十八烯酰胺 (Z)-9-Octadecenamide C18H35NO 30.918 0 0.73 1.30 β−谷甾醇 Beta-sitosterol C29H50O 33.250 0.24 0.74 1.33 3−[(三甲基硅)氧基]麦角甾−7−烯
3-[(Trimethylsilyl)oxy]ergost-7-eneC31H56OSi 35.274 0 0 0.98 1,6−双[甲基(三甲基)硅氧基]己烷
1,6-Bis[methyl (trimethylene) silyloxy]hexaneC14H30O2Si2 36.375 0 0 0.92 (1R,4R)−4−二丙基−1−甲基环己基−2−烯醇
(1R,4R)-4-Lsopropyl-1-methylcyclohex-2-enolC10H18O 38.780 0 0 0.90 丙丁酚 Probucol C31H48O2S2 38.935 75.78 0 0 3−羟基−(3β,5α,14β,20β,22β,25R)−螺甾−8−烯−11−酮
3-hydroxy-(3β,5α,14β,20β,22β,25R)-Spirost-8-en-11-oneC27H40O4 38.941 0 0 1.05 7−溴−8−氯−2−甲基十八烷
7-Bromo-8-chloro-2-methyloctadecaneC19H38BrCl 41.439 1.12 0 1.53 其他 Other 7.44 11.86 17.27 1) T2:15头蚜虫 + 5头白背飞虱;T4:5头蚜虫 + 15头白背飞虱;T6(CK): 0头蚜虫 + 0头白背飞虱
1) T2: 15 aphids + 5 white-backed planthoppers; T4: 5 aphids + 15 white-backed planthoppers; T6 (Control treatment): 0 aphid + 0 white-backed planthopper3. 讨论与结论
植食性口器昆虫种间互作包含竞争、中性、偏利及互利4类,其中,刺吸式口器昆虫间的种间竞争尤为明显[17-18]。本研究将白背飞虱和红腹缢管蚜在不同比例下进行混养,发现红腹缢管蚜初始量越大,对白背飞虱产卵量的抑制程度更明显,尤其以T2处理(15头蚜虫 + 5头白背飞虱)最为显著,甚至红腹缢管蚜的初始数量处于劣势下也能够显著抑制白背飞虱生殖,且日均产卵量也呈现近似的规律。相互竞争的动力体现在昆虫为争夺同一寄主资源产生了相互作用及影响,当两种昆虫生殖能力差异较大时,往往发生竞争取代现象,即生殖能力强的一方凭着绝对的数量优势取代竞争者[1,19]。蚜虫的生殖方式主要为孤雌生殖,具有严重的世代重叠[20],在生殖能力方面占据绝对的优势,与其他物种争夺同一寄主资源的过程中易凭借数量优势胜出。类似的种间竞争在其他昆虫中也有着广泛的报道。石永秀等[18]研究了不同异种共存密度对豌豆蚜Acyrthosiphon pisum与黑豆蚜Aphis fabae生长发育及生殖的影响,结果表明黑豆蚜能够显著抑制豌豆蚜的成蚜体质量及生殖能力,而黑豆蚜不受影响。王健立等[21]的研究结果表明,烟蓟马Thrips tabaci具有更强的繁殖能力且可以显著降低西花蓟马的产卵量,甚至西花蓟马在初始混合种群中占据较大比例的情况下,烟蓟马也可以经繁育后完全取代西花蓟马,与本研究的结论基本一致。但本文仅侧重于白背飞虱,未对红腹缢管蚜生长发育及繁殖力参数进行测定,同时仅测定了对当代白背飞虱繁殖力的影响,没有深究互作效应是否会对后代的生长发育及繁殖能力造成同等的影响,这些均需进一步研究。
此外,本研究通过测定水稻次生代谢物质含量,发现在T2处理(15头蚜虫 + 5头白背飞虱)中,高初始量的红腹缢管蚜能够降低水稻草酸含量,同时明显提高水稻黄酮及总酚含量,且与此对应的白背飞虱总产卵量显著降低。该结果表明昆虫之间的竞争除直接竞争外,还存在以寄主植物为纽带的间接竞争,即昆虫取食能够诱导寄主产生次生代谢物质,在调节物种间的竞争关系中起着关键作用[22-24]。一部分植食性昆虫的种间互作通常是由寄主植物介导的[17],纪祥龙等[25]发现桃蚜取食后可以改变甘蓝Brassica oleracea var. capitata中游离氨基酸含量,显著降低后取食的萝卜蚜Lipaphis erysimi种群数量。另外有研究表明,取食叶部的大菜粉蝶Pieris brassicae对取食根部的甘蓝地种蝇Delia radicum的生长发育有明显影响,与根组织中化感物质水平增加有一定的关联性[26]。张茂新等[11]已经从寄主植物中分离鉴定出多个具有抑制昆虫产卵的化合物,主要包含黄酮类、酚类、有机酸等化合物。枸杞瘿螨和枸杞木虱共同为害能够引起寄主植物总酚含量升高,进而抑制枸杞木虱种群数量的发展[8]。在马广民[27]的研究中,朱砂叶螨Tetranychus cinnabarinus取食后会诱导棉花Gossypium spp.的水杨酸信号途径增强,造成棉蚜Aphis gossypii发育历期延长及产卵量降低,施用外源性水杨酸亦引起相同的现象。本文因红腹缢管蚜可能是通过刺激寄主植物水稻产生防御性的次生代谢物质黄酮及总酚,其含量的积累能够抑制白背飞虱产卵量,从而证实水稻间接参与了种间互作,与前人的研究结果一致。但处理T5(20头白背飞虱)与处理T3(10头蚜虫 + 10头白背飞虱)的黄酮与总酚含量无差异性,而在产卵量方面差异显著,推测白背飞虱与红腹缢管蚜危害后诱导水稻产生了不同类型的防御性次生代谢物质,不同昆虫危害诱导寄主植物产生的防御途径有差异性[28]。另外,研究发现红腹缢管蚜与白背飞虱的数量初始比可能与水稻黄酮和总酚含量的变化有一定的联系,有待进一步研究。结合GC/MS仪器检测到了与酚类相关的化合物,其中丙丁酚物质仅存在于处理T2(15头蚜虫 + 5头白背飞虱),其规律也与上述的结论相匹配,其他相对含量有明显变化的成分也可能参与了种间互作。然而,仪器未检测到与草酸相关的次生代谢物质,这表明红腹缢管蚜与白背飞虱的互作效应可能对草酸没有明显影响。另外本文仅筛选出相对含量变化较大的成分,未明确其具体的作用,后续需进一步将上述变化幅度较大的成分添至人工饲料中,筛选并验证具有抑制白背飞虱生殖能力的成分。
本研究仅从植物的生理生化层面解释种间互作现象,没有深入到分子层面,因此在后续开展的试验中可以进一步对植物进行代谢组分析,以明确红腹缢管蚜与白背飞虱互作的分子基础,同时分析研究白背飞虱和红腹缢管蚜中与生殖发育相关基因表达情况,找出内在互作机制,为害虫的综合防控提供理论参考。
-
![]()
图 1 各处理组白背飞虱的单雌总产卵量及日均产卵量
T2:15头蚜虫 + 5头白背飞虱,T3:10头蚜虫 + 10头白背飞虱,T4:5头蚜虫 + 15头白背飞虱,T5:20头白背飞虱;图A中,柱子上方的不同小写字母表示处理间差异显著(P<0.05,Duncan’s法)
Figure 1. Total egg production and average daily spawning of white-backed planthopper in each treatment
T2: 15 aphids + 5 white-backed planthoppers, T3: 10 aphids + 10 white-backed planthoppers, T4: 5 aphids + 15 white-backed planthoppers, T5: 20 white-backed planthoppers; In figure A, different lowercase letters above the bars indicate significant differences among treatments (P<0.05, Duncan’s method)
![]()
图 2 各处理组水稻的草酸、黄酮和总酚含量
T1:20头蚜虫,T2:15头蚜虫 + 5头白背飞虱,T3:10头蚜虫 + 10头白背飞虱,T4:5头蚜虫 + 15头白背飞虱,T5:20头白背飞虱,T6(CK):0头蚜虫 + 0头白背飞虱;各图中,柱子上方的不同小写字母表示处理间差异显著(P<0.05,Duncan’s法)
Figure 2. Contents of oxalic acid, flavones and total phenols in rice of each treatment
T1: 20 aphids, T2: 15 aphids + 5 white-backed planthoppers, T3: 10 aphids + 10 white-backed planthoppers, T4: 5 aphids + 15 white-backed planthoppers, T5: 20 white-backed planthoppers, T6 (Control treatment): 0 aphid + 0 white-backed planthopper; In each figure, different lowercase letters above the bars indicate significant differences among treatments (P<0.05, Duncan’s method)
表 1 各试验处理的白背飞虱和红腹缢管蚜初始数量
Table 1 Initial number of white-backed planthopper and aphid in each experimental treatment
处理
Treatment蚜虫/头
Aphid白背飞虱/头
White-backed planthopperT1 20 0 T2 15 5 T3 10 10 T4 5 15 T5 0 20 T6(CK) 0 0  下载: 导出CSV
下载: 导出CSV
表 2 各处理组水稻主要次生代谢物质成分及相对含量
Table 2 The main secondary metabolite components and relative contents in rice of each treatment
化合物名称
Compound name分子式
Molecular formulat保留/min
Retention time相对含量1)/% Relative content T2 T4 T6 2,3−二氢−3,5−二羟基−6−甲基−4H−吡喃−4−酮
2,3-Dihydro-3,5-dihydroxy-6-methyl-4H-pyran-4-oneC6H8O4 5.664 0.67 3.71 13.21 萘 Naphthalene C10H8 5.931 0.41 0.95 2.82 2,4−二甲基苯甲醛 2,4-Dimethyl-benzaldehyde C9H10O 6.662 1.53 8.38 3.45 3−乙酰氧基−3−羟基丙酸甲酯
3-Acetoxy-3-hydroxypropionic acid-methyl esterC6H10O5 7.800 0 0 6.39 1−硝基−1−脱氧−d−甘油−l−甘露庚醇
1-Nitro-1-deoxy-d-glycero-l-mannoheptitolC7H15NO8 8.943 0 0 1.87 十二烷醛 Dodecanal C12H24O 11.407 0.08 1.07 1.70 5−羟甲基糠醛 5-Hydroxymethylfurfural C6H6O3 12.881 0.35 0 16.67 2,4−二叔丁基苯酚 2,4-Di-tert-butylphenol C14H22O 14.001 5.38 28.59 2.70 7,9−二叔丁基−1−氧杂螺(4,5)癸−6,9−二烯−2,8−二酮
7,9-Di-tert-butyl-1-oxaspiro(4,5)deca-6,9-diene-2,8-dioneC17H24O3 22.629 2.79 15.95 2.23 二苯基砜 Diphenyl sulfone C12H10O2S 23.036 4.21 28.02 23.68 9−十八烯酰胺 (Z)-9-Octadecenamide C18H35NO 30.918 0 0.73 1.30 β−谷甾醇 Beta-sitosterol C29H50O 33.250 0.24 0.74 1.33 3−[(三甲基硅)氧基]麦角甾−7−烯
3-[(Trimethylsilyl)oxy]ergost-7-eneC31H56OSi 35.274 0 0 0.98 1,6−双[甲基(三甲基)硅氧基]己烷
1,6-Bis[methyl (trimethylene) silyloxy]hexaneC14H30O2Si2 36.375 0 0 0.92 (1R,4R)−4−二丙基−1−甲基环己基−2−烯醇
(1R,4R)-4-Lsopropyl-1-methylcyclohex-2-enolC10H18O 38.780 0 0 0.90 丙丁酚 Probucol C31H48O2S2 38.935 75.78 0 0 3−羟基−(3β,5α,14β,20β,22β,25R)−螺甾−8−烯−11−酮
3-hydroxy-(3β,5α,14β,20β,22β,25R)-Spirost-8-en-11-oneC27H40O4 38.941 0 0 1.05 7−溴−8−氯−2−甲基十八烷
7-Bromo-8-chloro-2-methyloctadecaneC19H38BrCl 41.439 1.12 0 1.53 其他 Other 7.44 11.86 17.27 1) T2:15头蚜虫 + 5头白背飞虱;T4:5头蚜虫 + 15头白背飞虱;T6(CK): 0头蚜虫 + 0头白背飞虱
1) T2: 15 aphids + 5 white-backed planthoppers; T4: 5 aphids + 15 white-backed planthoppers; T6 (Control treatment): 0 aphid + 0 white-backed planthopper
下载: 导出CSV
-
[1] 陈堑. 麦长管蚜与禾谷缢管蚜种间竞争的研究[D]. 杨凌: 西北农林科技大学, 2010. [2] 邱良妙, 刘其全, 陈秀琴, 等. 六斑月瓢虫对草地贪夜蛾低龄幼虫的捕食作用[J]. 中国生物防治学报, 2023, 39(2): 471-477. [3] 张晓明, 徐海云, 杨念婉, 等. 两种蚜小蜂对烟粉虱MED隐种的田间笼罩控效评价[J]. 植物保护学报, 2018, 45(6): 1281-1288. [4] KAPLAN I, DENNO R F. Interspecific interactions in phytophagous insects revisited: A quantitative assessment of competition theory[J]. Ecology Letters, 2007, 10(10): 977-994. doi: 10.1111/j.1461-0248.2007.01093.x
[5] 刘晓飞, 陈强, 叶辉. 桔小实蝇与番石榴实蝇幼虫的种间竞争研究[J]. 环境昆虫学报, 2014, 36(1): 33-38. [6] 禹云超, 郅军锐, 曾广, 等. 入侵种西花蓟马与其它昆虫的种间竞争[J]. 环境昆虫学报, 2020, 42(1): 94-100. [7] ANDERSON P, SADEK M M, WÄCKERS F L. Root herbivory affects oviposition and feeding behavior of a foliar herbivore[J]. Behavioral Ecology, 2011, 22(6): 1272-1277. doi: 10.1093/beheco/arr124
[8] 李建领. 寄主植物介导的枸杞瘿螨和枸杞木虱种间互作关系研究[D]. 北京: 北京协和医学院, 2019. [9] ZHAO H P, ZHANG X Y, XUE M, et al. Feeding of whitefly on tobacco decreases aphid performance via increased salicylate signaling[J]. PLoS One, 2015, 10(9): e0138584. doi: 10.1371/journal.pone.0138584.
[10] 陈华才. 挥发物在水稻−二化螟、稻纵卷叶螟−二化螟绒茧蜂、螟蛉绒茧蜂相互关系中的作用[D]. 杭州: 浙江大学, 2002. [11] 张茂新, 凌冰, 庞雄飞. 非嗜食植物中的昆虫产卵驱避物及其利用[J]. 昆虫天敌, 2003, 25(1): 28-36. [12] 张钰明. 三氟苯嘧啶介导的核受体USP过表达对白背飞虱生殖发育的影响[D]. 雅安: 四川农业大学, 2022. [13] 段立珍, 汪建飞, 赵建荣. 比色法测定菠菜中草酸含量的条件研究[J]. 安徽农业科学, 2007, 35(3): 632-633. [14] 李明阳, 姚宇波, 徐翔, 等. 几种水稻对褐飞虱的抗性鉴定及抗性相关次生物质分析[J]. 应用昆虫学报, 2020, 57(6): 1375-1384. [15] 蔡文国, 吴卫, 邵金凤, 等. Folin-Ciocalteu法测定鱼腥草多酚的含量[J]. 食品科学, 2010, 31(14): 201-204. [16] PENG L, ZHAO Y, WANG H Y, et al. Comparative metabolomics of the interaction between rice and the brown planthopper[J]. Metabolomics, 2016, 12(8): 28746-28764.
[17] DENNO R F, MCCLURE M S, OTT J R. Interspecific interactions in phytophagous insects: Competition reexamined and resurrected[J]. Annual Review of Entomology, 1995, 40: 297-331. doi: 10.1146/annurev.en.40.010195.001501
[18] 石永秀, 上官超智, 王婷婷, 等. 黑豆蚜与豌豆蚜的种间竞争及密度效应[J]. 应用昆虫学报, 2022, 59(4): 862-873. [19] 吴佳昊, 黄波, 王德辉, 等. 光肩星天牛与星天牛种间竞争行为研究[J]. 环境昆虫学报, 2022, 44(3): 651-657. doi: 10.3969/j.issn.1674-0858.2022.03.16 [20] 张祥, 刘长仲, 宋维虎. 不同CO2浓度条件下两种色型豌豆蚜的种群密度效应[J]. 甘肃农业大学学报, 2019, 54(3): 78-83+92. [21] 王健立, 李洪刚, 马志国, 等. 西花蓟马与烟蓟马在紫甘蓝上的种间竞争[J]. 中国农业科学, 2011, 44(24): 5006-5012. doi: 10.3864/j.issn.0578-1752.2011.24.005 [22] 闫文静, 王俊刚, 张玉栋, 等. 棉长管蚜和棉蚜对受蚜虫取食胁迫棉花植株的选择行为[J]. 新疆农业科学, 2019, 56(1): 52-60. [23] PAPADOPOULOU G V, DAM N M. Mechanisms and ecological implications of plant-mediated interactions between belowground and aboveground insect herbivores[J]. Ecological Research, 2017, 32(1): 13-26. doi: 10.1007/s11284-016-1410-7
[24] NGUYEN D, RIEU I, MARIANI C, et al. How plants handle multiple stresses: Hormonal interactions underlying responses to abiotic stress and insect herbivory[J]. Plant Molecular Biology, 2016, 91(6): 727-740. doi: 10.1007/s11103-016-0481-8
[25] 纪祥龙, 刘长庆, 胡玲玲, 等. 桃蚜与萝卜蚜交互为害对寄主氮营养及蚜虫种间竞争的调节[J]. 中国农学通报, 2019, 35(4): 97-101. [26] SOLER R, BEZEMER T M, CORTESERO A M, et al. Impact of foliar herbivory on the development of a root-feeding insect and its parasitoid[J]. Oecologia, 2007, 152(2): 257-264. doi: 10.1007/s00442-006-0649-z
[27] 马广民. 朱砂叶螨取食诱导的棉花防御反应及其对棉蚜发育的影响[D]. 北京: 中国农业大学, 2017. [28] WALLING L L. Avoiding effective defenses: Strategies employed by phloem-feeding insects[J]. Plant Physiology, 2008, 146(3): 859-866. doi: 10.1104/pp.107.113142
-
期刊类型引用(0)
其他类型引用(1)
计量
- 文章访问数: 99
- HTML全文浏览量: 34
- PDF下载量: 22
- 被引次数: 1
