Effects of ridge irrigation and nitrogen reduction on paddy field CH4 emission, soil organic acid content and expression of enzyme encoding genes
-
摘要:目的
研究不同垄面宽度的垄作灌溉和不同时期减施氮肥对稻田甲烷(CH4)排放、土壤有机酸含量以及CH4形成和转化相关酶编码基因(产CH4古菌中甲基辅酶M还原酶编码基因mcrA和CH4氧化菌中CH4单加氧酶编码基因sMMO)表达量的影响,揭示土壤有机酸以及mcrA和sMMO表达量对稻田CH4通量的影响。
方法开展3种灌溉模式(淹水灌溉,垄宽分别为80、100 cm的垄作灌溉)和3种施氮处理(常规施氮:135 kg·hm−2,其中苗肥47.25 kg·hm−2、分蘖肥54.00 kg·hm−2和穗肥33.75 kg·hm−2;返青期减氮:110 kg·hm−2,其中苗肥22.25 kg·hm−2、分蘖肥54.00 kg·hm−2和穗肥33.75 kg·hm−2;孕穗期减氮:110 kg·hm−2,其中苗肥47.25 kg hm−2、分蘖肥54.00 kg hm−2和穗肥8.75 kg hm−2)的田间试验,测定生育期内稻田CH4通量、土壤有机酸含量以及mcrA和sMMO表达量,分析稻田CH4通量、土壤有机酸含量以及mcrA和sMMO表达量之间的相互关系。
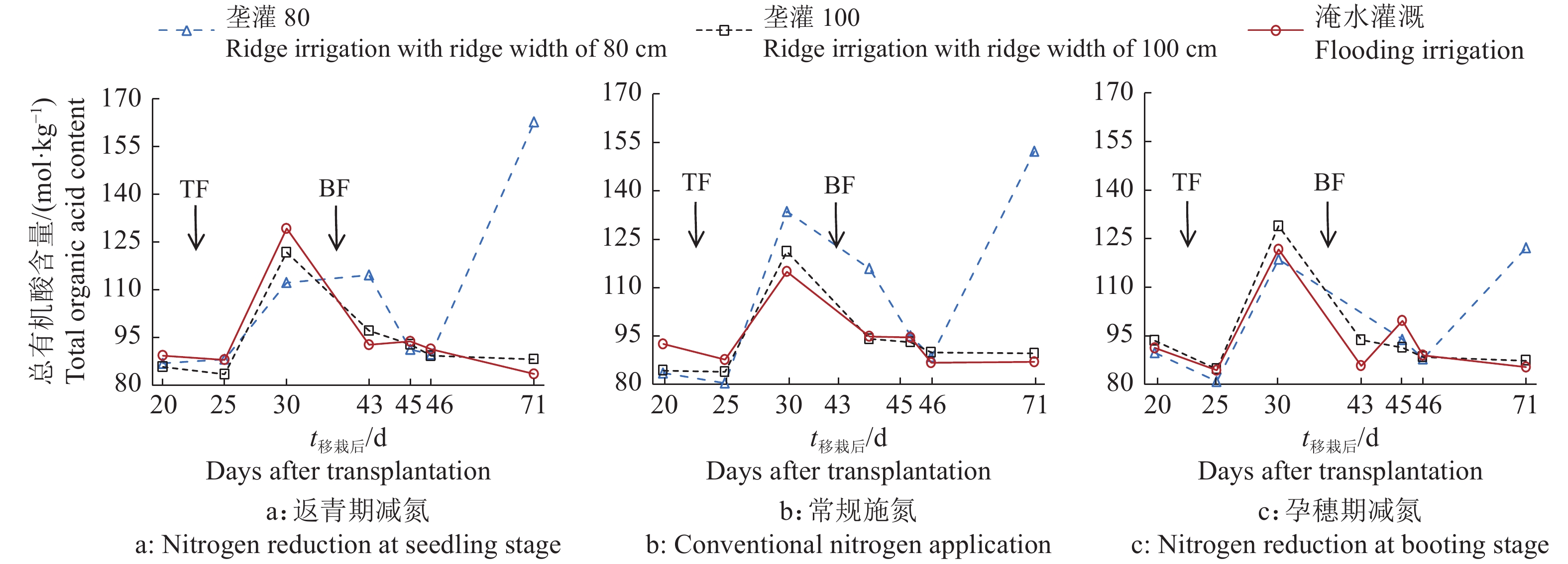
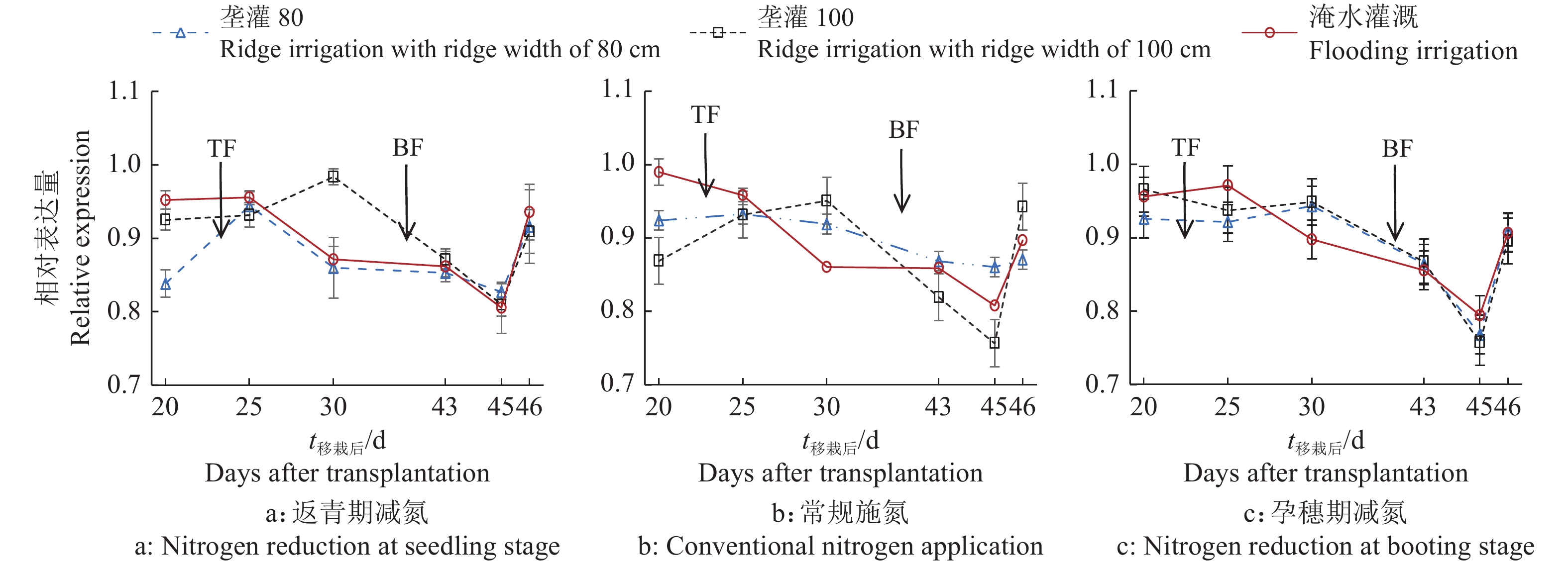
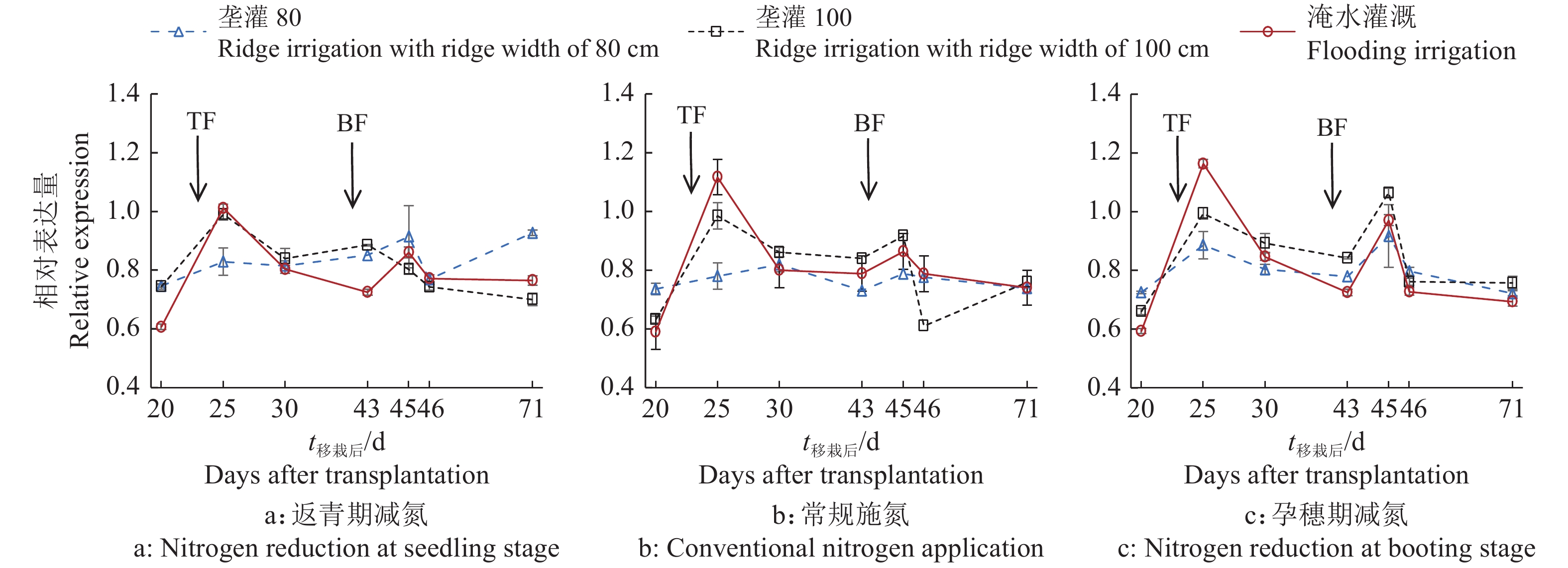
结果相同施氮处理下,垄作灌溉稻田CH4排放量较淹水灌溉显著降低,相同灌水模式下,返青期减氮稻田CH4排放量较常规施氮显著降低。垄宽为80 cm的垄作灌溉+常规施氮处理在施肥后10 d土壤总有机酸含量较施肥前高71.7%,淹水灌溉和常规施氮处理高28.8%。返青期减氮下,垄宽为80 cm的垄作灌溉土壤mcrA表达量整体低于淹水灌溉,sMMO表达量除移栽后25 d外均高于淹水灌溉。稻田CH4通量与土壤mcrA表达量和总有机酸含量显著相关(P<0.01),相关系数分别为0.644和−0.348,土壤总有机酸含量与mcrA、sMMO表达量显著相关(P<0.05),相关系数分别为−0.240和0.197。
结论垄宽80 cm垄作灌溉+返青期减氮处理降低了稻田CH4通量,土壤总有机酸含量和mcrA表达量显著影响稻田CH4通量,sMMO表达量可能间接影响稻田CH4通量。
Abstract:ObjectiveTo investigate the effects of ridge irrigation with different ridge widths and reduced nitrogen fertilizer at different growth stages on methane (CH4) emissions, soil organic acid content and expression of enzyme encoding genes related to CH4 formation and transformation (including methyl coenzyme M reductase gene mcrA in methanogenic archaea, and methane monooxygenase gene sMMO in methane oxidizing bacteria) in paddy fields, and reveal the effects of soil organic acid content and the expression of mcrA, sMMO on CH4 fluxes in paddy fields.
MethodA field experiment was conducted with three irrigation modes (flooding irrigation, ridge irrigation with ridge widths of 80, 100 cm) and three nitrogen treatments (conventional nitrogen application: 135 kg·hm−2, including seedling fertilizer 47.25 kg hm−2, tillering fertilizer 54.00 kg hm−2 and booting fertilizer 33.75 kg hm−2; Nitrogen reduction at seedling stage: 110 kg·hm−2, including seedling fertilizer 22.25 kg hm−2, tillering fertilizer 54.00 kg hm−2 and booting fertilizer 33.75 kg hm−2; Nitrogen reduction at booting stage: 110 kg·hm−2 , including seedling fertilizer 47.25 kg hm−2, tillering fertilizer 54.00 kg hm−2 and booting fertilizer 8.75 kg hm−2). The paddy field CH4 fluxes, soil organic acid content and mcrA, sMMO expression levels in different treatments were measured, and the relationships among them were analyzed.
ResultUnder the same nitrogen treatment, ridge irrigation significantly reduced CH4 emission from paddy fields compared with flooding irrigation. Under the same irrigation mode, nitrogen reduction at seedling stage significantly reduced CH4 emission from paddy field compared with conventional nitrogen application. The total organic acid content of soil in ridge irrigation with ridge width of 80 cm + nitrogen reduction at seedling stage treatment was 71.7% higher than that before fertilization on the 10th day after fertilization, and 28.8% higher in flooding irrigation + conventional nitrogen application treatment. Under the condition of nitrogen reduction at seedling stage, the expression of mcrA in ridge irrigation with ridge width of 80 cm soil was overall lower than that in flooding irrigation soil, and the expression of sMMO was higher than that in flooding irrigation except 25 days after transplanting. CH4 fluxes were significantly correlated with mcrA expression and total organic acid content in soils (P<0.01), with correlation coefficients of 0.644 and −0.348, respectively. There were significant correlations between soil total organic acid content and the expressions of mcrA and sMMO (P < 0.05), with correlation coefficients of −0.240 and 0.197, respectively.
ConclusionThe ridge irrigation with the ridge width of 80 cm + nitrogen reduction at seedling stage treatment can reduce CH4 flux from paddy fields. Soil total organic acid content and mcrA expression significantly affect paddy field CH4 flux, while soil sMMO expression may indirectly affect CH4 flux.
-
烟草青枯病是一种由青枯雷尔菌Ralstonia solanacearum引起的系统性侵染病害,现已成为世界各产烟区最主要的病害之一[1]。该病在我国河南、山东、江苏、云南、广西、广东、福建等地普遍发生,烟株一旦染病,往往整株死亡,其危害是毁灭性的,给生产造成重大经济损失[2]。因此,研究青枯菌致病机理和寻找有效的防治青枯病的方法是当前植物病理学研究的重要课题之一, 其中抗病育种防治青枯病的生化机制最为重要。国内外学者对大量的相关病害进行研究,发现植物在受病害侵袭过程中表现出一系列复杂的生理生化变化,包括体内代谢的变化、细胞内活性氧的积累与清除、抗病信号的产生与转导、防卫反应的表达与调控等[3-4]。在这一复杂过程中,一些与植物抗病性相关的酶类起着很重要的调控作用[5-6],如苯丙氨酸解氨酶(Phenylalanine ammonium lyase, PAL)、超氧化物歧化酶(Superoxide dismutase,SOD)、过氧化物酶(Peroxidase,POD)和多酚氧化酶(Polyphenol oxidase,PPO)等。同工酶谱分析作为一种认识基因存在和表达的工具已被大量运用于种质分类与种间遗传差异分析[7-9],其规律性变化可以作为一种早期的鉴别手段,用来研究植物的抗病性问题。因此,笔者选择了2个不同抗性的烟草品种,进行青枯病菌的接种试验,在接菌后不同时段及时采样,测定与病程相关的防御性酶活性以及代谢组分,从而了解烟草抗、感青枯病的变化过程,旨为烟草抗青枯病育种提供生化方面的依据。
1. 材料与方法
1.1 材料
试验所用烟草品种:高度感病品种长脖黄及高度抗病品种粤烟97,均由广东南雄烟草科学研究所提供;烟草青枯雷尔菌由广东省农科院何自福研究员惠赠;病菌的保存与增殖参照匡传富等[10]的方法。
仪器:LRH型生化培养箱(广东省韶关市鑫腾科普有限公司);FC—18R台式高速冷冻离心机(厦门仪达仪器有限公司);DYCZ-24DN型垂直电泳槽(北京六一仪器厂);DYY 11B三恒电泳仪(北京六一仪器厂);10 μL微量进量器(德国Eppendorf AG);UV-8500PC型紫外-可见分光光度计(上海天美科学仪器公司);1702-MP8型电子天平(德国Startorius公司);岛津GC-MS Q2010 Ultra(日本Shimadzu公司)。
1.2 试验方法
选取长势一致、3~4叶龄的健康烟草植株供试,将其移栽至装有灭菌土的18 cm×23 cm的塑料盆中,每个处理为5株,设置3个重复,放置于室外环境下。待烟苗生长7 d后,接菌组参照黎定军等[11]茎部注射法,用注射器将1 mL青枯雷尔菌液(5×108 cfu·mL-1)从叶腋处注入烟茎基部的维管束里,并在接菌处注意保湿,对照组为清水处理的烟苗。分别在处理1、3、5、7、9、11 d后,取同一叶位处的烟叶进行相关生化指标的测定,其中代谢组分差异分析选择接菌后的第3天进行。
1.2.1 防御性酶活性测定
PAL的提取及活性测定参照薛应龙等[12]方法;SOD的提取与测定参照邹琦[13]163-165的方法;POD的提取与测定参照张志良[14]的方法;PPO的提取与活性测定参照李靖等[15]的方法。PAL以每分钟D290 nm变化0.01所需要的酶量定义为1个酶活性单位(U);POD和PPO分别以每分钟D470 nm、D410 nm变化1.00所需要的酶量定义为1个酶活性单位(U);SOD以每分钟D560 nm抑制50 %所需要的酶量定义为1个酶活性单位(U)。
1.2.2 POD和PPO同工酶电泳
酶液制备:取0.1 g样品,加入0.1 mol·L-1预冷的Tris-HCl (pH 6.8)缓冲液1.5 mL,冰浴研磨,12 000 r·min-1、4 ℃条件下离心15 min,取上清液,于-70 ℃冰箱保存备用。
电泳:采用不连续的聚丙烯酰胺凝胶垂直板电泳技术[13]131-135,上样缓冲液不加SDS和β-巯基乙醇,电极缓冲液为pH 8.3的Tris-Gly缓冲液,样品不经加热处理。同工酶电泳采用聚丙烯酰胺为7.5%的分离胶、4%的浓缩胶。样品上样量为10 μL[V(样品缓冲液): V(粗酶液)=1: 1],于4 ℃条件下电泳,浓缩胶80 V、电泳30 min,分离胶120 V、电泳1.5 h。以溴酚蓝作指示剂,电泳结束后,取下胶进行染色。
POD同工酶染色:称取0.1 g联苯胺,加少量无水乙醇溶解,依次加入5 mol·L-1的醋酸溶液10 mL,1.5 mol·L-1的醋酸钠溶液10 mL,水70 mL,最后加入3~5滴质量分数为30%的过氧化氢溶液。将凝胶电泳胶带放入装有染色液的塑料小盆中,并轻轻晃动,待条带清晰后,立即用相机拍照记录。
PPO同工酶染色:3份质量分数为1%邻苯二酚溶液,1份0.05 mol·L-1磷酸缓冲液(pH6.8),1份质量分数为0.06%对苯二胺溶液,混合均匀后,将胶带放入染色液中3~5 min,即可见到棕红色的多酚氧化酶酶带。
1.2.3 代谢组分差异分析
烟草样品前处理方法:参考Tikunov等[16]和Roessner等[17]建立的方法。
气相色谱条件:进样口温度250 ℃。升温程序:初始温度50 ℃,保持1 min,以5 ℃·min-1升温速度升到150 ℃保持2 min,最后以10 ℃·min-1升温速度升到250 ℃,保持10 min。以高纯氦气(体积分数大于99.999%)为载气采用不分流进样方式,进样量1 μL,载气流速1 mL·min-1。
质谱条件:电子轰击源(EI),离子源温度200 ℃,四极杆温度150 ℃,接口温度250 ℃,电子能量70 eV,溶剂延迟时间4 min;调谐方式为标准调谐,质谱扫描方式为全扫描,扫描范围为50~500 aum。使用NIST 2011谱库进行图谱检索。
1.3 数据分析
试验数据采用Microsoft Excel 2003和SPSS11.5数据处理软件进行数据统计分析,酶活性数值均为平均值±标准差。应用Duncan's新复极差法进行差异显著性分析。
2. 结果与分析
2.1 几种防御性酶活性变化
2.1.1 PAL活性变化
从图 1可知,抗病品种粤烟97的PAL活性在整个测定时期,接菌组与对照组均高于感病品种长脖黄。抗病品种粤烟97在接菌后第7天酶活性显著高于对照组(P<0.05),其余时间酶活性均低于对照组;感病品种长脖黄在接菌后第5天,PAL活性显著高于对照组(P<0.05),其余时间酶活性均低于对照组。
![图 1 不同抗病烟草品种接种青枯雷尔菌后叶片PAL活性(以鲜质量计)的变化]() 图 1 不同抗病烟草品种接种青枯雷尔菌后叶片PAL活性(以鲜质量计)的变化相同时间同一烟草品种不同柱子上方凡是有一个相同小写字母者,表示该品种对照组和接菌组间差异不显著(P>0.05,Duncan's法)。Figure 1. Changes of PAL activities (based on fresh mass) in leaves of resistant and susceptible tobacco cultivars after inoculation of Ralstonia solanacearum
图 1 不同抗病烟草品种接种青枯雷尔菌后叶片PAL活性(以鲜质量计)的变化相同时间同一烟草品种不同柱子上方凡是有一个相同小写字母者,表示该品种对照组和接菌组间差异不显著(P>0.05,Duncan's法)。Figure 1. Changes of PAL activities (based on fresh mass) in leaves of resistant and susceptible tobacco cultivars after inoculation of Ralstonia solanacearum2.1.2 SOD活性变化
从图 2可知,抗病品种粤烟97的SOD活性在整个测定时期,接菌组与对照组均高于感病品种长脖黄。抗病品种粤烟97在接菌后第3、5、7、9天,SOD活性均高于对照组,其中第7天差异显著(P<0.05);感病品种长脖黄在接菌后第1、3、5天,SOD活性均高于对照组,其中第5天差异显著(P<0.05)。由此得知,抗、感品种在受到青枯菌侵害后,均能使SOD迅速被激活,从而起到清除或阻止病原菌所造成伤害的作用。
![图 2 不同抗病烟草品种接种青枯雷尔菌后SOD活性(以鲜质量计)的变化]() 图 2 不同抗病烟草品种接种青枯雷尔菌后SOD活性(以鲜质量计)的变化相同时间同一烟草品种不同柱子上方凡是有一个相同小写字母者,表示该品种对照组和接菌组间差异不显著(P>0.05,Duncan's法)。Figure 2. Changes of SOD activities (based on fresh mass) in leaves of resistant and susceptible tobacco cultivars after inoculation of Ralstonia solanacearum
图 2 不同抗病烟草品种接种青枯雷尔菌后SOD活性(以鲜质量计)的变化相同时间同一烟草品种不同柱子上方凡是有一个相同小写字母者,表示该品种对照组和接菌组间差异不显著(P>0.05,Duncan's法)。Figure 2. Changes of SOD activities (based on fresh mass) in leaves of resistant and susceptible tobacco cultivars after inoculation of Ralstonia solanacearum2.1.3 POD活性变化
从图 3可知,抗病品种粤烟97的POD活性在整个测定时期,接菌组与对照组均高于感病品种长脖黄。抗病品种粤烟97在接菌后第3天,酶活性显著高于对照组(P<0.05),其余时间酶活性均显著低于对照组(P<0.05);感病品种长脖黄在接菌后第3、5天,酶活性高于对照组,其中第5天差异显著(P<0.05),其余时间POD活性均低于对照组。
![图 3 不同抗病烟草品种接种青枯雷尔菌后POD活性(以鲜质量计)的变化]() 图 3 不同抗病烟草品种接种青枯雷尔菌后POD活性(以鲜质量计)的变化相同时间同一烟草品种不同柱子上方凡是有一个相同小写字母者,表示该品种对照组和接菌组间差异不显著(P>0.05,Duncan's法)。Figure 3. Changes of POD activities (based on fresh mass) in leaves of resistant and susceptible tobacco cultivars after inoculation of Ralstonia solanacearum
图 3 不同抗病烟草品种接种青枯雷尔菌后POD活性(以鲜质量计)的变化相同时间同一烟草品种不同柱子上方凡是有一个相同小写字母者,表示该品种对照组和接菌组间差异不显著(P>0.05,Duncan's法)。Figure 3. Changes of POD activities (based on fresh mass) in leaves of resistant and susceptible tobacco cultivars after inoculation of Ralstonia solanacearum2.1.4 PPO活性变化
从图 4可知,抗病品种粤烟97对照组、接菌组的PPO活性均高于感病品种长脖黄的。接菌后第3天,粤烟97和长脖黄的PPO活性略高于各自的对照组,其余时间均低于各自的对照组。
![图 4 不同抗病烟草品种接种青枯雷尔菌后PPO活性(以鲜质量计)的变化]() 图 4 不同抗病烟草品种接种青枯雷尔菌后PPO活性(以鲜质量计)的变化相同时间同一烟草品种不同柱子上方凡是有一个相同小写字母者,表示该品种对照组和接菌组间差异不显著(P>0.05,Duncan's法)。Figure 4. Changes of PPO activities (based on fresh mass) in leaves of resistant and susceptible tobacco cultivars after inoculation of Ralstonia solanacearum
图 4 不同抗病烟草品种接种青枯雷尔菌后PPO活性(以鲜质量计)的变化相同时间同一烟草品种不同柱子上方凡是有一个相同小写字母者,表示该品种对照组和接菌组间差异不显著(P>0.05,Duncan's法)。Figure 4. Changes of PPO activities (based on fresh mass) in leaves of resistant and susceptible tobacco cultivars after inoculation of Ralstonia solanacearum2.2 接菌处理对不同品种烟草叶片中POD和PPO同工酶表达的影响
2.2.1 对POD同工酶的影响
从图 5A可看出,抗病品种粤烟97接菌后,整个测定期间POD同工酶谱带未增加,但凝胶电泳谱带宽度与色度均发生变化。接菌后第3天,接菌组的同工酶谱带较对照组谱带宽,且颜色较深;接菌后第1、11天,接菌组POD同工酶谱带色度比对照组弱;接菌后第5天,接菌组POD同工酶谱带条数比对照组少,且部分条带颜色较浅;接菌后第7、9天,接菌组与对照组POD同工酶谱带无明显差异。从图 5B可看出,感病品种长脖黄在接菌后第1、5、7、9天,接菌组POD同工酶谱带颜色均比对照组颜色浅;接菌后第3、11天,同工酶谱带宽度和颜色与对照基本无差别。对比图 5 A、图 5 B可知,抗病品种粤烟97的POD同工酶谱带为6条,感病品种长脖黄的POD同工酶谱带为2条,说明抗病品种粤烟97的POD同工酶类型多于感病品种长脖黄的POD同工酶类型。
![图 5 不同抗病烟草品种接种青枯雷尔菌后叶片POD同工酶谱带]() 图 5 不同抗病烟草品种接种青枯雷尔菌后叶片POD同工酶谱带CK:对照组,TR:接菌组;A:抗病品种粤烟97,B:感病品种长脖黄。Figure 5. Electrophoretic isoenzyme patterns of POD in leaves of resistant and susceptible tobacco cultivars after inoculation of Ralstonia solanacearum
图 5 不同抗病烟草品种接种青枯雷尔菌后叶片POD同工酶谱带CK:对照组,TR:接菌组;A:抗病品种粤烟97,B:感病品种长脖黄。Figure 5. Electrophoretic isoenzyme patterns of POD in leaves of resistant and susceptible tobacco cultivars after inoculation of Ralstonia solanacearum2.2.2 对PPO同工酶的影响
从图 6A可以看出,抗病品种粤烟97接菌后第1、7天,接菌组与对照组的PPO同工酶谱带无明显差异;接菌后第3天,PPO同工酶谱带宽度与色度均高于对照组;接菌后第5天,PPO同工酶谱条带数与色度均低于对照组;接菌后第9、11天,部分PPO同工酶谱带色度略低于对照组。从图 6B可以看出,感病品种长脖黄接菌后第1、9、11天,PPO同工酶谱带条数、色度与对照组无明显差异;接菌后第3天,PPO同工酶谱带色度均低于对照组;接菌后第5和7天,PPO同工酶谱带色度略高于对照组。对比图 6 A、图 6 B可知,抗病品种粤烟97的PPO同工酶谱带最多为6条,最少为4条,感病品种长脖黄的POD同工酶谱带为2条,说明抗病品种粤烟97的PPO同工酶类型多于感病品种长脖黄的PPO同工酶类型。
![图 6 不同抗病烟草品种接种青枯雷尔菌后叶片PPO同工酶谱带]() 图 6 不同抗病烟草品种接种青枯雷尔菌后叶片PPO同工酶谱带CK:对照组, TR:接菌组;A:抗病品种粤烟97, B:感病品种长脖黄。Figure 6. Electrophoretic isoenzyme patterns of PPO in leaves of resistant and susceptible tobacco cultivars after inoculation of Ralstonia solanacearum
图 6 不同抗病烟草品种接种青枯雷尔菌后叶片PPO同工酶谱带CK:对照组, TR:接菌组;A:抗病品种粤烟97, B:感病品种长脖黄。Figure 6. Electrophoretic isoenzyme patterns of PPO in leaves of resistant and susceptible tobacco cultivars after inoculation of Ralstonia solanacearum2.3 代谢组分差异分析
对不同抗病品种接菌处理后叶片提取物进行GC-MS测定, 得到总离子流图(图 7)。从图 7可以直观地看出不同抗病品种间存在明显差异,接菌组与对照组间,峰的数量和强度也都存在差异。
![图 7 不同抗病烟草品种接种青枯雷尔菌后代谢组分的GS-MS总离子流图]() 图 7 不同抗病烟草品种接种青枯雷尔菌后代谢组分的GS-MS总离子流图1:感病品种长脖黄对照组;2:感病品种长脖黄接菌组;3:抗病品种粤烟97对照组;4:抗病品种粤烟97接菌组。Figure 7. Total ion chromatograms of metabolites of resistant and susceptible tobacco cultivars after inoculation of Ralstonia solanacearum
图 7 不同抗病烟草品种接种青枯雷尔菌后代谢组分的GS-MS总离子流图1:感病品种长脖黄对照组;2:感病品种长脖黄接菌组;3:抗病品种粤烟97对照组;4:抗病品种粤烟97接菌组。Figure 7. Total ion chromatograms of metabolites of resistant and susceptible tobacco cultivars after inoculation of Ralstonia solanacearum通过岛津质谱工作站对总离子流图的处理解析, 峰面积归一化法测量了各组分的百分含量。通过检索和解析初步分离最多确定了70个化合物。鉴定的各组分及相对质量分数见表 1。
表 1 不同抗病烟草品种接种青枯雷尔菌后品种间代谢物差异比较1)Table 1. Comparison of metabolites from resistant and susceptible tobacco cultivars after inoculation of Ralstonia solanacearum
从表 1可以看出烟株被青枯雷尔菌侵染后代谢物差异较为明显,粤烟97接菌后烟叶样本中代谢物相对质量分数高于其对照组,且长脖黄接菌后相对质量分数低于其对照组的是乳酸、甘油酸、肌肉肌醇、L-苏糖酸、苹果酸、L-吡咯烷酮-5-羧酸、烟碱、丝氨酸、L-苏氨酸、D-来苏糖、核糖醇、β-D-果糖、D-(+)-塔罗糖、棕榈酸、硬脂酸,说明烟草抗青枯病能力的强弱可能与这些物质有关,或者与参与形成上述代谢物的酶活性有关;2个不同抗病品种接菌后共同产生或者相对质量分数比对照增高的物质分别为:L-缬氨酸、草酸、L-异亮氨酸、丙二酸、磷酸、L-天冬氨酸、赤藓糖醇、L-苯丙氨酸、十四烷醇、L-阿拉伯糖、D-木糖,不同抗病品种烟草在受到青枯雷尔菌侵袭时均会刺激体内代谢物含量的改变,其含量的增高说明上述代谢物可能参与抗病相关物质的形成。
3. 讨论与结论
与植物抗病相关的PAL、SOD、PPO和POD均属于植物防卫系统相关酶,因此可作为植物抗病的生化指标。在植物体内,PAL是苯丙烷类代谢途径的关键酶和限速酶,催化L-苯丙氨酸解氨生成反式肉桂酸(合成各种酚类及木质素的前体物质),因而调控与植物抗病有关的酚类物质和木质素在植物体中的合成和积累[18]。研究表明,PAL活性与植物抗病性呈正相关[15, 19]。本研究中,在整个测定期间,抗病品种粤烟97的PAL活性均高于感病品种长脖黄,说明烟草品种抗病强弱,与体内PAL活性呈正相关。SOD作为植物体内活性氧清除系统重要保护酶之一,能清除超氧化物阴离子自由基,提高植物抗逆性[20]。本研究表明,抗病品种粤烟97在接菌后第3、5、7、9天,SOD活性均大于对照,感病品种长脖黄在接菌后第1、3、5天,SOD活性均高于对照,抗病品种SOD活性在接菌组与对照组中均高于感病品种,说明在青枯雷尔菌侵染后抗病品种可高效地清除活性氧的伤害,感病品种由于本身SOD活性低,接菌后酶活性增长幅度较小而使其受到伤害。POD是一种应激酶,其活性与植物抗病性有着密切的关系,在活性氧的清除和维持植物体内活性氧的正常水平中起作用;POD还能催化木质素合成,木质化的细胞壁机械性能加强,不透水气,阻止营养物质、水分、色素等的扩散,使病原菌无法获得营养而死亡[21-22]。研究表明,POD活性与植物的抗病性具有正相关关系[23-26]。PPO是一类广泛分布于植物体内的质体金属酶,能直接以O2为氧化底物将酚氧化成醌,从而抑制病虫害的侵袭。PPO不仅参与酚类物质的氧化,同时也参与木质素的形成[27]。本研究发现抗病品种粤烟97的POD活性高于感病品种长脖黄,且POD同工酶谱分析表明,抗病品种粤烟97的谱带条数均要多于感病品种长脖黄。接菌后抗病品种的POD活性迅速升高,而感病品种则较弱,2品种的POD活性随着时间延长均降低,说明青枯雷尔菌侵染烟草后均能刺激体内POD活性的提高,且抗病品种总体强于感病品种。抗病品种粤烟97和感病品种长脖黄接菌后第3天,PPO活性略高于对照组,随着时间延长酶活性均降低。抗病品种粤烟97中PPO活性高于感病品种长脖黄,可能与其抗病性强有关。
烟草植株受到病菌侵染时,引起相关酶活性的改变,继而引起植株体内代谢物质的改变,形成的代谢物本身具有抗病作用或者进一步通过转化形成与抗病有关的次级代谢物。例如PAL酶活性高低会影响L-苯丙氨酸含量[28],POD酶活性影响某些糖类、氨基酸类等与木质素合成有关物质的含量[29]。本研究发现,粤烟97与长脖黄在接菌处理3 d时,SOD、POD、PPO活性均比对照高,而PAL活性均低于对照组,因此植株体内相关酶催化形成的物质应该有所提高,或者酶促反应的底物应该有所降低。代谢物检测表明,2个抗病性不同的品种接菌后共同产生或者相对质量分数相比对照增高的物质分别为:L-缬氨酸、草酸、L-异亮氨酸、丙二酸、磷酸、L-天冬氨酸、赤藓糖醇、L-苯丙氨酸、十四烷醇、L-阿拉伯糖、D-木糖,说明此类代谢物含量的变化可能与防御性酶活性有关;粤烟97接菌后烟叶样本中代谢物相对质量分数高于其对照组,但长脖黄接菌后相对质量分数低于其对照组的物质分别为:乳酸、甘油酸、肌肉肌醇、L-苏糖酸、苹果酸、L-吡咯烷酮-5-羧酸、烟碱、丝氨酸、L-苏氨酸、D-来苏糖、核糖醇、β-D-果糖、D-(+)-塔罗糖、棕榈酸、硬脂酸,说明此类代谢物含量的变化与防御性酶活性变化无关,可能与其他酶活性变化有关。研究表明,肌醇在植物体内通过自身氧化途径可以代谢为与植物细胞壁合成息息相关的多糖,从而提高植物抗病性[30]。烟碱作为一种防御性物质,在烟草受到病虫害侵袭时会刺激体内产生大量烟碱[31],此外,相关研究表明,在细胞质中苹果酸酶催化苹果酸脱羧形成的丙酮酸参与莽草酸途径,进一步形成丙酮酸族氨基酸和一些与防御反应有关的次生代谢物质[32]。L-苏糖酸作为L-抗坏血酸的一种降解产物,广泛存在于植物体内,其含量的增加,说明抗坏血酸积极参与抗氧化作用,保护机体免于自由基的威胁[33]。不同烟草品种抗青枯病的机理,不仅与其体内防御性酶活性有关,可能还与某些代谢物或与其相关的酶活性变化有关。
-
![]()
图 1 垄作灌溉与不同时期减施氮肥下稻田土壤总有机酸含量的变化
TF:分蘖肥,BF:孕穗肥
Figure 1. Changes of total organic acid content in paddy soil under ridge irrigation and nitrogen reduction at different growth stages
TF: Tillering fertilizer, BF: Booting fertilizer
![]()
图 2 垄作灌溉和不同时期减施氮肥下稻田土壤甲基辅酶M还原酶编码基因mcrA表达量
TF:分蘖肥,BF:孕穗肥
Figure 2. Methyl coenzyme M reductase encoding gene mcrA expression level in paddy soil under ridge irrigation and nitrogen reduction at different growth stages
TF: Tillering fertilizer, BF: Booting fertilizer
![]()
图 3 垄作灌溉与不同时期减施氮肥下土壤CH4单加氧酶编码基因sMMO表达量
TF:分蘖肥,BF:孕穗肥
Figure 3. CH4 monooxygenase encoding gene sMMO expression level in paddy soil under ridge irrigation and nitrogen reduction at different growth stages
TF: Tillering fertilizer, BF: Booting fertilizer
![]()
图 4 垄作灌溉和不同时期减施氮肥下稻田CH4通量的变化情况
SF:苗肥,TF:分蘖肥,BF:孕穗肥
Figure 4. Changes of CH4 flux in paddy field under ridge irrigation and nitrogen reduction at different growth stages
SF: Seedling fertilizer; TF: Tillering fertilizer, BF: Booting fertilizer
表 1 垄作灌溉和不同时期减施氮肥下稻田CH4累积排放量(E)1)
Table 1 Cumulative CH4 emission (E) in paddy field under ridge irrigation and nitrogen reduction at different growth stages
项目
Item处理
Treatment返青期
Regreening
stage分蘖前期
Early tillering
stage分蘖后期
Late tillering
stage拔节孕穗期
Jointing-booting
stage抽穗期
Heading
stage成熟期
Maturing
stage合计
TotalE/(kg·hm−2) YNC 128.2±0.5a 240.6±32.1b 144.9±7.9b 40.2±9.6g 77.3±13c 68.0±16.5d 699.1±2.9b L1NC 57.1±6.5d 114.5±12.8f 89.8±26.3g 68.4±9.2b 176.3±50.2a 47.3±15.4g 553.4±8.2e L2NC 57.9±4.1d 173.6±10.1d 135.1±11d 36.9±3.4h 44.4±50.3e 36.7±3.8h 484.6±2.6f YNS 100.8±3.9b 239.3±30.5b 103.9±19.4f 45.7±20.3e 67.1±11.5d 83.1±7.5b 640.0±5.6c L1NS 78.7±1.1c 111.3±13.5 53.9±7.0i 18.2±3.6i 44.7±5.6e 50.3±12.8f 357.1±1.5h L2NS 66.8±3.3cd 216.4±19c 139.0±9.9c 42.0±12.5f 28.9±15.2f 78.2±1.6c 571.4±2.6d YNB 107.7±6.6b 253.6±30.5a 153.0±25.5a 83.1±13.8a 76.1±42.8c 128.1±53.5a 801.6±9.0a L1NB 68.2±1.6cd 130.4±21.7e 76.9±3.2h 51.8±6.4c 67.5±25.8d 54.7±9.0e 449.5±3.2g L2NB 58.5±7.5d 174.6±9.9d 114.4±22.7e 48.4±9.0d 98.7±7.0b 46.4±8.9g 541.1±8.7e P IM 0.443 <0.001 <0.001 0.720 0.530 <0.05 <0.05 NT <0.05 0.784 0.926 0.106 0.243 0.446 0.181 IM×NT 0.667 <0.001 <0.001 0.243 <0.001 <0.05 <0.05 1)同列数据后不同小写字母表示在P<0.05水平差异显著(Duncan’s法);Y:淹水灌溉,L1:垄灌80,L2:垄灌100;NC:常规施氮,NS:返青期减氮,NB:孕穗期减氮;IM:灌溉模式,NT:施氮处理
1) Different lowercase letters in the same column indicate significant differences at P<0.05 level (Duncan’s method); Y: Flooding irrigation, L1: Ridge irrigation with ridge width of 80 cm, L2: Ridge irrigation with ridge width of 100 cm; NC: Conventional nitrogen application, NS: Nitrogen reduction at seedling stage, NB: Nitrogen reduction at booting stage; IM: Irrigation mode, NT: Nitrogen treatment 下载: 导出CSV
下载: 导出CSV
表 2 稻田CH4通量与土壤总有机酸含量和酶编码基因表达量的相互关系1)
Table 2 Correlation among CH4 flux from paddy field, total organic acid content and encoding gene expression level in paddy soil
指标
IndexmcrA表达量
mcrA expression levelsMMO表达量
sMMO expression level总有机酸含量
Total organic acid contentCH4通量 CH4 flux 0.644** −0.111 −0.348** mcrA表达量 mcrA expression level −0.150 −0.240* sMMO表达量 sMMO expression level 0.197* 1)“*”“**”分别表示在P<0.05、P<0.01水平显著相关(Pearson法)
1) “*” and “**” indicate significant correlations at P<0.05 and P<0.01 levels respectively (Pearson method)
下载: 导出CSV
-
[1] IPCC. Climate change 2014: Synthesis report[R]//PACHAURI R K, MEYER L A. Contribution of working groups I, II and III to the fifth assessment report of the intergovernmental panel on climate change. Geneva, Switzerland, 2014: 87.
[2] SMITH P, MARTINO D, CAI Z, et al. Policy and technological constraints to implementation of greenhouse gas mitigation options in agriculture[J]. Agriculture Ecosystems and Environment, 2007, 118(1/2/3/4): 6-28.
[3] 刘英烈. 不同氮肥水平集约化栽培模式双季稻生态系统温室气体收支的田间观测[D]. 南京: 南京农业大学, 2016. [4] 张自常, 李鸿伟, 陈婷婷, 等. 畦沟灌溉和干湿交替灌溉对水稻产量与品质的影响[J]. 中国农业科学, 2011, 44(24): 4988-4998. doi: 10.3864/j.issn.0578-1752.2011.24.003 [5] 马微微, 陈灿, 黄璜, 等. 垄作稻鱼鸡共生对稻田土壤养分含量及水稻产量的影响[J]. 河南农业科学, 2021, 50(8): 9-17. [6] ZENG Y, LI F S. Ridge irrigation reduced greenhouse gas emission in double-cropping rice field[J]. Archives of Agronomy and Soil Science, 2021, 67(8): 1003-1016. doi: 10.1080/03650340.2020.1771698
[7] 曹开勋, 赵坤, 金王飞飞, 等. 水氮互作对稻田温室气体排放的影响[J]. 土壤学报, 2022, 59(5): 1386-1396. [8] 胡敏杰, 仝川, 邹芳芳. 氮输入对土壤甲烷产生、氧化和传输过程的影响及其机制[J]. 草业学报, 2015, 24(6): 204-212. doi: 10.11686/cyxb2014313 [9] 姜珊珊. 氮肥减量及不同品种肥料配施对稻麦农田CH4和N2O(直接和间接)排放的影响[D]. 南京: 南京农业大学, 2017. [10] BODELIER P L, ROSLEV P, HENCKEL T, et al. Stimulation by ammonium-based fertilizers of methane oxidation in soil around rice roots[J]. Nature, 2000, 403(6768): 421-424. doi: 10.1038/35000193
[11] 江瑜, 朱相成, 钱浩宇, 等. 水稻丰产与稻田甲烷减排协同的研究展望[J]. 南京农业大学学报, 2022, 45(5): 839-847. [12] 董艳, 杨智仙, 董坤, 等. 供氮水平对小麦生长特性及根系分泌有机酸的影响[C]//中国土壤学会. 面向未来的土壤科学(上册): 中国土壤学会第十二次全国会员代表大会暨第九届海峡两岸土壤肥料学术交流研讨会论文集. 成都: 电子科技大学出版社, 2012: 481-485. [13] 蒋娜, 陈紫娟, 曹轶, 等. 低温湿地甲烷古菌及其介导的甲烷产生途径[J]. 微生物学通报, 2013, 40(1): 137-145. [14] 余锋, 李思宇, 邱园园, 等. 稻田甲烷排放的微生物学机理及节水栽培对甲烷排放的影响[J]. 中国水稻科学, 2022, 36(1): 1-12. [15] AWASTHI M K, WAINAINA S, MAHBOUBI A, et al. Methanogen and nitrifying genes dynamics in immersed membrane bioreactors during anaerobic co-digestion of different organic loading rates food waste[J]. Bioresource Technology, 2021, 342: 125920. doi: 10.1016/j.biortech.2021.125920.
[16] JODTS R J, ROSS M O, KOO C W, et al. Coordination of the copper centers in particulate methane monooxygenase: Comparison between methanotrophs and characterization of the CuC site by EPR and ENDOR spectroscopies[J]. Journal of the American Chemical Society, 2021, 143(37): 15358-15368. doi: 10.1021/jacs.1c07018
[17] YUAN J, YUAN Y, ZHU Y, et al. Effects of different fertilizers on methane emissions and methanogenic community structures in paddy rhizosphere soil[J]. Science of the Total Environment, 2018, 627: 770-781. doi: 10.1016/j.scitotenv.2018.01.233
[18] 鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社, 2000. [19] QIAN L, CHEN L, JOSEPH S, et al. Biochar compound fertilizer as an option to reach high productivity but low carbon intensity in rice agriculture of China[J]. Carbon Management, 2014, 5(2): 145-154. doi: 10.1080/17583004.2014.912866
[20] TAN S D, CHEN B, ZHENG H B, et al. Effects of cultivation techniques on CH4 emissions, net ecosystem production, and rice yield in a paddy ecosystem[J]. Atmospheric Pollution Research, 2018, 10(1): 274-282.
[21] 马艳婷, 赵志远, 冯天宇, 等. 有机肥配施对苹果园温室气体排放的影响[J]. 农业环境科学学报, 2021, 40(9): 2039-2048. [22] SCHERER R, RYBKA A C P, BALLUS C A, et al. Validation of a HPLC method for simultaneous determination of main organic acids in fruits and juices[J]. Food Chemistry, 2012, 135(1): 150-154. doi: 10.1016/j.foodchem.2012.03.111
[23] 马倩倩, 吴翠云, 蒲小秋, 等. 高效液相色谱法同时测定枣果实中的有机酸和VC含量[J]. 食品科学, 2016, 37(14): 149-153. doi: 10.7506/spkx1002-6630-201614026 [24] HAN J I, SEMRAU J D. Quantification of gene expression in methanotrophs by competitive reverse transcription-polymerase chain reaction[J]. Environmental Microbiology, 2004, 6(4): 388-399. doi: 10.1111/j.1462-2920.2004.00572.x
[25] ZHU Y, LI Y, ZHENG N, et al. Similar but not identical resuscitation trajectories of the soil microbial community based on either DNA or RNA after flooding[J]. Agronomy, 2020, 10(4): 502. doi: 10.3390/agronomy10040502.
[26] HENRY A, DOUCETTE W, NORTON J, et al. Changes in crested wheatgrass root exudation caused by flood, drought, and nutrient stress[J]. Journal of Environmental Quality, 2007, 36(3): 904-912. doi: 10.2134/jeq2006.0425sc
[27] 徐国伟, 孙会忠, 陆大克, 等. 不同水氮条件下水稻根系超微结构及根系活力差异[J]. 植物营养与肥料学报, 2017, 23(3): 811-820. [28] 戢林, 李廷轩, 张锡洲, 等. 水稻氮高效基因型根系分泌物中有机酸和氨基酸的变化特征[J]. 植物营养与肥料学报, 2012, 18(5): 1046-1055. [29] 王钧美, 张莉, 徐桃元, 等. 不同灌溉稻田产甲烷菌与甲烷产生率的变化规律[J]. 灌溉排水学报, 2014, 33(Z1): 360-363. [30] 王智平, 胡春胜, 杨居荣. 无机氮对土壤甲烷氧化作用的影响[J]. 应用生态学报, 2003(2): 305-309. doi: 10.3321/j.issn:1001-9332.2003.02.033 [31] 熊浩, 张保成, 李建柱, 等. 灌水量对冬小麦农田土壤N2O与CO2排放的影响[J]. 灌溉排水学报, 2020, 39(9): 41-50. [32] 马艳芹, 钱晨晨, 孙丹平, 等. 施氮水平对稻田土壤温室气体排放的影响[J]. 农业工程学报, 2016, 32(S2): 128-134. [33] 范玉婧, 姚惠娇, 高志岭, 等. 氮源对甲烷氧化混合菌群甲烷氧化和氧化亚氮排放的影响[J]. 河北农业大学学报, 2021, 44(3): 54-61. [34] 丁维新, 蔡祖聪. 温度对甲烷产生和氧化的影响[J]. 应用生态学报, 2003(4): 604-608. [35] SINGH A, SINGH R S, UPADHYAY S N, et al. Community structure of methanogenic archaea and methane production associated with compost-treated tropical rice-field soil[J]. FEMS Microbiology Ecology, 2012, 82(1): 118-134. doi: 10.1111/j.1574-6941.2012.01411.x
[36] LIU P, KLOSE M, CONRAD R. Temperature effects on structure and function of the methanogenic microbial communities in two paddy soils and one desert soil[J]. Soil Biology and Biochemistry, 2018, 124: 236-244. doi: 10.1016/j.soilbio.2018.06.024
-
期刊类型引用(6)
1. 梁丽丽, 钟璐瑶, 张辽, 侯俊, 束良佐. 生物炭及其替代泥炭比例对樱桃萝卜生长的影响. 中国瓜菜. 2025(06)  百度学术
百度学术
2. 侯赛赛,仝姗姗,王鹏企,谢冰雪,张瑞芳,王鑫鑫. 生物炭和秸秆对不同作物生长性状和养分吸收的影响. 中国农业科技导报(中英文). 2025(04): 179-191 . 百度学术
3. 吉光鹏,张栋海,牛蛉磊,吴玉蓉,常学艳. 密植苹果根系空间分布特征研究. 农业科技通讯. 2024(03): 111-114 . 百度学术
4. 杨凌,陈泽涛,陈嘉显,陈祝锋,曾曙才,何茜,苏艳,邱权. 减氮配施生物炭对闽楠生长和光合特性的影响. 林草资源研究. 2024(02): 116-123 . 百度学术
5. 兰宇,孟军,韩晓日,陈温福. 生物炭基产品及其对土壤培肥改良效应的研究进展. 植物营养与肥料学报. 2024(07): 1396-1412 . 百度学术
6. 刘丹妮,陈伟盛,黄连喜,刘忠珍,王艳红,魏岚. 生物炭-椰糠栽培基质特性及其对香蕉组培苗生长的影响. 中国农学通报. 2024(28): 112-118 . 百度学术
其他类型引用(7)