
Regulation mechanism and breeding application of rice floret-opening-time
-
摘要:
水稻为严格自花授粉作物。花时是水稻的重要农艺性状,受到内部因素(植物激素、遗传因子等)和外部环境因素(温度、光照、湿度、CO2浓度等)的复杂调控。合适的开花时间是水稻成功繁殖的重要保障。颖花过早开放会导致水稻易受低温、露水重引起的病虫害影响;而过晚开放易使水稻遭受高温胁迫,降低产量。此外,在杂交水稻制种过程中,确保父母本的花时一致是提高杂交水稻制种产量的重要条件之一。本文从控制水稻颖花开放的结构基础、生理基础、遗传基础以及分子机理等方面,系统总结了水稻花时调控的研究进展,提出了今后水稻花时研究应重点解决的问题,并讨论了花时性状的改良在水稻生产和籼粳亚种间杂交稻育种上的应用价值。
Abstract:Rice (Oryza sativa L.) is a strictly autogamous crop. Floret-opening-time (FOT) is an important agronomic trait in rice, which is complicatedly regulated by internal factors (plant hormones, genetic factors, etc.) and external environmental factors (temperature, light, humidity, CO2 concentration, etc.). Floret opening at an appropriate time is very critical for the success of rice reproduction. If the floret opens too early in the morning, rice is vulnerable to pathogen infection caused by low temperature and heavy dew. In contrast, if the floret opens too late, the seed setting of rice would be severely affected by high temperature in the afternoon. Moreover, synchronized FOT between the male and female parents is a key requirement for successful production of hybrid seeds. In this review, we summarize the research progresses on the regulation of FOT in rice, mainly focusing on the aspects including the structural basis, physiological basis, genetic basis and molecular mechanism of controlling rice floret opening. We also present some important topics for future in-depth studies of FOT. Finally, we discuss the value of modulating FOT trait for improving rice production and indica-japonica inter-subspecies hybrid rice breeding.
-
Keywords:
- Oryza sativa L. /
- Floret-opening-time /
- Jasmonic acid /
- Lodicule /
- Hybrid seed production
-
磷是植物生长发育的必需营养元素之一,不仅是生物大分子如核酸,蛋白质和脂质等的重要组成成分,而且参与多种代谢过程,例如核苷酸合成、光合作用、能量传递和信号转导等[1-4]。在土壤中,全磷质量分数一般在0.2~1.1 g·kg−1之间,其形态主要分为无机态磷和有机态磷[5]。其中,能直接被植物直接吸收利用的是无机磷酸根离子(
${{\rm{H}}_2}{\rm{PO}}_4^{ - 1}$ 或${\rm{HPO}}_4^{ - 2}$ ),其含量一般占土壤全磷的0.1%左右,难以满足作物生长的需求[6-8]。当培养介质中有效磷浓度较低时,作物体内各种代谢活动受到显著的影响,从而表现出生长迟缓、植株瘦弱等缺磷症状,最终导致作物减产[9]。据统计,在全世界范围内低磷胁迫导致农作物产量减少30%~40%[7]。在我国的农业生产中,主要通过施用磷肥来缓解土壤中磷有效性低的问题。但是,由于作物对磷肥的当季利用率不到25%和磷肥的过量施用,我国土壤磷素不断累积,土壤有效磷质量浓度从20世纪90年代的17.1 mg·L−1增加到2012年的33.3 mg·L−1,全国平均磷(P2O5)盈余可达59.2 kg·hm−2[10-11]。过量施用磷肥也带来了水体富营养化等一系列环境问题,并加速了自然界磷矿资源的枯竭[12]。因此,优化磷肥资源的田间管理和培育磷肥利用率高的作物品种,并对二者进行有机结合,是提高磷肥利用效率的必由之路,也是农业可持续发展的必然要求[7, 13-14]。在长期进化过程中,植物形成了一系列适应低磷胁迫的机制,包括重建根系的形态构型、促进根系分泌物(如有机酸和酸性磷酸酶)的分泌、形成植物–微生物共生体系等[1-4, 15]。阐明植物适应低磷胁迫的分子和遗传机制,不仅可以解析作物磷高效的机理,而且为培育磷高效的作物品种奠定了理论基础。本文总结了近十年来关于低磷胁迫调控根系分泌物合成和分泌的研究进展,旨在阐明通过控制作物根系分泌物来提高作物磷效率的途径,为培育磷高效作物品种和优化磷肥的田间管理提供思路。
1. 低磷胁迫下植物根系分泌物的功能
根系分泌物是植物在生长过程中,根系以主动或被动方式向培养介质分泌的所有物质的总称,包括有机/无机离子、植物激素、根细胞脱落物及其分解产物等[16-17]。根系分泌物的合成和分泌过程是实现植物与根际环境互作的重要途径,不仅可以改善根际土壤性质,调控根际微生物的群落组成,而且植物可以通过感受根系分泌物及其根际微生态的变化,反馈调节植物的生长[17]。以往的研究表明,低磷胁迫可以调控植物根系紫色酸性磷酸酶(Purple acid phosphatase, PAP)活性以及有机酸的合成和分泌,从而分别影响植物对根际有机磷和难溶性无机磷活化利用[17-19]。
2. 根系分泌紫色酸性磷酸酶的功能及其调控机制
酸性磷酸酶(Acid phosphatase, APase; E.C. 3.1.3.2)是一类能催化水解磷酸单酯或酸酐,释放无机磷酸根离子(Pi)的水解酶类,其酶活性的最适pH一般低于7.0[20]。在植物中,具有酸性磷酸酶酶学性质的蛋白种类比较多,其中一类是紫色酸性磷酸酶。紫色酸性磷酸酶除了具备以上酸性磷酸酶的特性外,还具有其他明显的生物化学特征,即蛋白存在双金属离子的催化中心、在水溶液中呈紫色或粉红色以及L−酒石酸对其活性无明显抑制作用等[19, 21-22]。
2.1 紫色酸性磷酸酶的蛋白结构和酶学特性
随着不同物种基因组测序完成,紫色酸性磷酸酶家族成员的组成、表达模式及其功能在不同植物中都有报道,如拟南芥Arabidopsis thaliana、水稻Oryza sativa、大豆Glycine max和菜豆Phaseolus vulgaris等[19, 23-26]。虽然,不同物种之间紫色酸性磷酸酶的同源性较低,但是其结构上具有高度的保守性,例如催化位点均含有β-α-β-α-β折叠[19, 27-28]。同时,紫色酸性磷酸酶的氨基酸序列均包含5个保守的结构域,即 DXG/G DXX Y/G NH(D/E)/VXX H/G HX H(下划线表示7个保守的氨基酸),其中保守结构域中包含的7个保守氨基酸可以结合Fe3+-X2+(X2+表示Fe2+,Mn2+或Zn2+)等金属离子,形成紫色酸性磷酸酶的催化中心[28-31]。以往的研究表明,其催化中心的三价金属离子具有高度保守性,主要是Fe3+。但是,二价离子的种类会存在差异,例如大豆和菜豆的紫色酸性磷酸酶包含Zn2+、在黄花羽扇豆Lupinus luteus中包含Mn2+、在甘薯Ipomoea batatas中包含Zn2+或Mn2+,表明紫色酸性磷酸酶在酶催化中心结构上的多样性[32-35]。
根据植物紫色酸性磷酸酶单亚基的大小,其成员可分为大分子量和小分子量的紫色酸性磷酸酶[19, 28]。其中,小分子量紫色酸性磷酸酶的相对分子质量约为35 000,常以单体的形式存在于植物体中,如拟南芥的AtPAP17和菜豆的PvPAP3等[25, 36]。大分子量紫色酸性磷酸酶的亚基相对分子质量在45 000~75 000之间[19, 28]。在植物体中,大分子量紫色酸性磷酸酶的亚基通常会通过二硫键或非共价结合的方式形成同质二聚体或异质二聚体,如菜豆的KbPAP和黄羽扇豆的LlPPD1[28, 35, 37]。
一般而言,紫色酸性磷酸酶对含有磷酸单脂键的底物都具有水解活性,尤其是对三磷酸腺苷(ATP)、焦磷酸(PPi)、磷酸化糖类、磷酸化氨基酸或植酸等的水解活性较高。但是,同源性相差较远的紫色酸性磷酸酶的最适底物具有显著的差别。例如,在植物紫色酸性磷酸酶的进化树中(图1),属于Ia亚家族成员的大部分紫色酸性磷酸酶对ATP和磷酸烯醇丙酮酸盐(PEP)具有较高的水解酶活性,包括拟南芥的AtPAP10、AtPAP12、AtPAP25和AtPAP26,大豆的GmSAP,菜豆的KeACP (PvPAP1)和KbPAP (PvPAP2),甘薯的IbPAP1,轮花大戟Euphorbia characias的ELPAP,洋葱Allium cepa的AcPEPP和烟草Nicotiana tabacum的NtPAP12等[38-46]。而且,NtPAP12和AtPAP25还兼有蛋白磷酸酶的酶学性质,分别参与了细胞壁中的α−木糖苷酶和β−葡糖苷酶脱磷酸化过程[43, 46]。与其他紫色酸性磷酸酶不同,Ib-1亚家族的紫色酸性磷酸酶大部分对植酸具有较高的水解活性,包括拟南芥的AtPAP15和AtPAP23、水稻的OsPHY1、烟草的NtPAP、大豆的GmPhy和GmPAP14、玉米Zea mays的ZmPAPhy_b、大麦Hordeum vulgare的HvPAPhy_a和HvPAPhy_b2、小麦Triticum aestivum的TaPAPhy_a1和TaPAPhy_b1、枸橘Poncirus trifoliate的PtPAP3等[47-55]。在IIb亚家族中,部分紫色酸性磷酸酶成员具有核苷酸水解酶的活性。根据金属离子是否激活其活性,将其分为依赖金属离子的双磷酸核苷磷酸酶/磷酸二酯酶和不依赖金属离子的核苷酸焦磷酸酶/磷酸二酯酶[56-58]。其中,黄羽扇豆的LlPPD1是最早发现的依赖金属离子的双磷酸核苷磷酸酶/磷酸二酯酶[35]。紫云英Astragalus sinicus的AsPPD1具有水解磷酸二酯酶活性,也是依赖金属离子的双磷酸核苷磷酸酶/磷酸二酯酶[59]。但是,不依赖金属离子的核苷酸焦磷酸酶/磷酸二酯酶主要催化核苷酸或核苷酸糖类的焦磷酸键/磷酸二酯键的水解[56-57]。例如,在水稻中对UDP−葡萄糖和淀粉合成前体ADP−葡萄糖具有水解活性的OsNPP1和OsNPP6,以及对UDP和ADP具有水解活性的OsNPP2[60]。第III亚家族紫色酸性磷酸酶中,只有拟南芥的AtPAP17 (AtACP5)和菜豆的PvPAP3具有相关的报道[25, 36]。其中,菜豆的PvPAP3对ATP具有较高的水解活性,暗示了PvPAP3参与ATP代谢的生物学功能[25]。虽然,紫色酸性磷酸酶最适底物的研究结果为阐明其生物学功能奠定了基础,但是,由于体外试验难以准确模拟植物细胞内复杂的催化条件,而且植物细胞内存在多种含磷酸单酯键的代谢物,难以逐一进行测试。所以,只有结合生物化学、遗传学和分子生物学等研究手段才能明确紫色酸性磷酸酶的生物学功能。
![]() 图 1 植物紫色酸性磷酸酶进化树分析Ac: 洋葱Allium cepa; As: 紫云英Astragalus sinicus; At: 拟南芥Arabidopsis thaliana; Ec: 轮花大戟Euphorbia characias; Gm: 大豆Glycine max; Hv: 大麦Hordeum vulgare; Ib: 甘薯Ipomoea batatas; La: 白花羽扇豆Lupinus albus; Ll: 黄花羽扇豆Lupinus luteus; Mt: 截形苜蓿Medicago truncatula; Nt: 烟草Nicotiana tabacum; Os: 水稻Oryza sativa; Pt: 枳Poncirus trifoliate; Pv: 菜豆Phaseolus vulgaris; St: 马铃薯Solanum tuberosum; Ta: 小麦Triticum aestivum; Zm: 玉米Zea maysFigure 1. Phylogenetic tree analysis of plant purple acid phosphatase
图 1 植物紫色酸性磷酸酶进化树分析Ac: 洋葱Allium cepa; As: 紫云英Astragalus sinicus; At: 拟南芥Arabidopsis thaliana; Ec: 轮花大戟Euphorbia characias; Gm: 大豆Glycine max; Hv: 大麦Hordeum vulgare; Ib: 甘薯Ipomoea batatas; La: 白花羽扇豆Lupinus albus; Ll: 黄花羽扇豆Lupinus luteus; Mt: 截形苜蓿Medicago truncatula; Nt: 烟草Nicotiana tabacum; Os: 水稻Oryza sativa; Pt: 枳Poncirus trifoliate; Pv: 菜豆Phaseolus vulgaris; St: 马铃薯Solanum tuberosum; Ta: 小麦Triticum aestivum; Zm: 玉米Zea maysFigure 1. Phylogenetic tree analysis of plant purple acid phosphatase2.2 磷有效性对植物紫色酸性磷酸酶表达的调控
低磷胁迫在转录、翻译和蛋白修饰等水平调控紫色酸性磷酸酶的活性,从而促进了植物体内或分泌的酸性磷酸酶的活性[19]。例如:对不同植物紫色酸性磷酸酶家族成员表达模式进行分析,发现低磷胁迫显著提高了拟南芥中29个成员中的9个、大豆中35个成员中的23个、水稻26个成员中的10个、玉米33个成员中的11个紫色酸性磷酸酶基因的表达[19, 61]。同时,也有报道,低磷胁迫增强紫色酸性磷酸酶蛋白积累,比如番茄Lycopersicon esculentum的LeSAP1和LeSAP2、拟南芥的AtPAP10、AtPAP12和AtPAP26,菜豆的PvPAP3,白花羽扇豆Lupinus albus的LaSAP2和大豆GmPAP1-like等[21, 25, 36, 45-46, 62-64]。而且,最近的研究结果表明,在植物的磷信号网络中存在调控紫色磷酸酶表达的重要调控因子。例如:在水稻中,OsPAP10a的表达受转录因子OsMYB2P-1、OsPHR2以及OsSPX-MSF1的正调控,但受到生长素响应因子OsARF12和OsSPX1、OsSPX3和OsSPX5的负调控[13, 65];在拟南芥中,AtPAP10的表达受转录因子AtPHR1和AtPHL2的正调控,但受到AtTHO1和AtTHO3的负调控[66-67];同时,有研究结果显示在拟南芥中乙烯信号途径参与调控了AtPAP10的表达[24]。这些结果说明,低磷胁迫参与调控紫色酸性磷酸酶的表达。但是,由于其组成的复杂性决定了低磷胁迫对其表达调控的途径具有多样性。
2.3 根分泌的紫色酸性磷酸酶活化利用有机磷的机制
根据紫色酸性磷酸酶的酶学特性和低磷促进其酶活性增加的特性,一般认为紫色酸性磷酸酶参与了植物体内和根际有机磷的活化利用[19, 28]。据报道,根系分泌或根系细胞壁定位的紫色酸性磷酸酶能水解根际或质外体空间的有机磷化合物,释放出无机磷酸根离子,从而参与植物对有机磷的活化利用。在菜豆中,Liang等[25]通过分子筛和亲和层析等蛋白纯化的方法,从菜豆根系纯化获得了一个受低磷上调表达的紫色酸性磷酸同工酶PvPAP3,结合其最适底物的酶学性质和超量表达该基因的表型,揭示了PvPAP3参与菜豆根系对外源ATP活化利用的功能。随后,在拟南芥和水稻中也报道了根系分泌紫色酸性磷酸酶具有类似的功能[13, 45, 63, 65]。除了对外源ATP有活化功能外,在大豆、菜豆、柱花草Stylosanthes guianensis和拟南芥中发现根系分泌的紫色酸性磷酸酶具有活化外源dNTP、DNA、ADP和6−磷酸果糖等有机磷的功能,暗示了植物根系分泌的紫色酸性磷酸酶功能的多样性[13, 45, 61, 63-65, 68]。
除了核酸磷、磷酸化糖类和磷酸化氨基酸等有机磷外,土壤还存在着大量的植酸磷。早期通过转基因技术,在植物中超量表达分泌型紫色酸性磷酸酶能显著提高转基因植物根际植酸酶活性,从而增强其对外源植酸磷的活化利用能力。例如,在大豆中超量表达含分泌信号肽的AtPAP15,能显著促进转基因大豆对植酸磷活化利用的能力[69]。此外,在白三叶Trifolium repens中超量表达来源于蒺藜苜蓿Medicago sativa的MtPHY1,在烟草中超量表达来源于水稻OsPHY1或来源于白花羽扇豆的LaSAP3,也发现类似的结果[26, 70-71]。最近,在柱花草根系中克隆了低磷加强表达的紫色酸性磷酸酶SgPAP23基因,其编码的蛋白对植酸磷具有较高的酶活性,而且超量表达该基因显著提高了转基因菜豆毛根和拟南芥根系分泌植酸酶的活性,从而提高对外源植酸磷的活化利用能力,进一步揭示了根系分泌植酸酶活化利用根际植酸磷的机制[54]。
3. 根系有机酸的分泌及其活化利用磷的功能
植物在生长的过程中会通过主动或被动的方式向根际分泌多种有机酸[72-73]。低磷条件下,大多数植物根系有机酸的组成和分泌量均明显改变,这对于促进土壤难溶性无机磷的活化利用,改善植物的根际磷营养具有十分重要的作用[17-18]。
3.1 有机酸对土壤难溶性无机磷的活化作用
土壤中难溶性无机磷只有在水解后产生磷酸根离子,才能被植物直接吸收利用。在难溶性无机磷的水解过程中,根系分泌的有机酸具有重要的作用[2, 17]。在早期的研究中发现,在土壤中添加有机酸能够降低土壤对磷酸根离子的吸持作用,而且三元羧酸对土壤磷酸根离子的解吸附作用最强,二元羧酸次之,一元羧酸最弱[74]。在比较草酸对FePO4、CaHPO4、AlPO4和磷矿粉等难溶性无机磷活化利用的研究中发现,草酸对AlPO4的活化效果较显著[75]。现在,一般认为有机酸参与活化土壤难溶性无机磷的主要机制包括:1)与土壤中的铁、铝和钙等金属离子进行络合反应,提高这些含磷化合物的溶解性;2)有机酸阴离子与磷酸根离子之间竞争土壤颗粒的结合位点,降低土壤对磷酸根的吸附;3)通过络合反应,改变土壤中的铁铝氧化物等吸附剂表面的电荷,从而降低其对磷酸根的吸附固定;4)降低根际pH,促进难溶性无机磷的溶解等[17, 76]。
3.2 低磷对植物根系有机酸合成和分泌的影响
有机酸是植物碳代谢过程中重要的中间产物。低磷胁迫下,植物细胞内与有机酸合成相关的途径会发生改变。同时,有机酸可以通过细胞质膜上的特定转运蛋白或通道分泌到根际。在油菜Brassica napus的研究中发现,低磷条件下其叶片中的磷酸烯醇式丙酮酸羧化酶的活性和柠檬酸含量均显著增加。而且14CO2标记结果显示,叶片合成的柠檬酸大部分转运至根系及根系分泌物中[77]。随后,在番茄、豌豆Pisum sativum和菜豆等植物中也发现低磷胁迫促进了磷酸烯醇式丙酮酸羧化酶的活性,从而增强了根系对有机酸的分泌[78-80]。另外,在白花羽扇豆的研究中发现,低磷胁迫除了加强磷酸烯醇式丙酮酸羧化酶的活性,还增加了柠檬酸合成酶和苹果酸脱氢酶等与有机酸代谢相关酶的活性,从而促进白花羽扇豆排根中有机酸的分泌[81-83]。这些结果说明了植物可以协同调控有机酸的合成和分泌来适应低磷胁迫。
虽然,促进根系有机酸的分泌是植物适应低磷胁迫的普遍机制,但不同植物种类,甚至是同种植物内不同基因型间,其分泌的有机酸种类和数量都存在显著的差异。据报道,低磷胁迫显著促进了玉米、水稻、柱花草和洋白菜Brassica oleraceae等植物根系柠檬酸的分泌[84-87]。在白花羽扇豆和大豆中,低磷胁迫同时促进了柠檬酸和苹果酸的分泌,尤其是白花羽扇豆柠檬酸的分泌量增加了13倍左右[81, 88-89];除了苹果酸和柠檬酸以外,低磷胁迫下植物根系也会分泌其他类型的有机酸包括番石榴酸、草酸、酒石酸和丁二酸等。例如,在低磷条件下木豆Cajanus cajan根系可以分泌大量的番石榴酸[90]。而且在大麦中,对难溶性无机磷活化能力强的基因型分泌的柠檬酸较多,表明了柠檬酸的分泌是影响大麦活化利用难溶性无机磷的重要机制。
3.3 低磷对有机酸合成和分泌的分子调控机制
3.3.1 有机酸合成的分子调控机制
有机酸是植物碳代谢过程中重要的中间产物,所以,碳代谢过程相关酶的活性及其编码基因的表达量会影响有机酸的合成。目前,低磷胁迫调控有机酸合成相关的酶及其基因主要是苹果酸脱氢酶(Malate dehydrogenase, MDH)和柠檬酸合成酶(Citrate synthase, CS)。对多种植物的研究发现,低磷胁迫明显提高了植物MDH基因的表达水平,暗示了其表达量与苹果酸合成和分泌具有较高的相关性[91-94]。随后,在棉花Gossypium hirsutum的研究中发现,超量表达GhmMDH1基因后,其根系苹果酸浓度和分泌量显著增加。而且,在供给Al-P、Fe-P和Ca-P等难溶性无机磷时,超量表达GhmMDH1的转基因株系具有较高的生物量,且磷含量高,进一步说明了GhmMDH1基因可以通过调控苹果酸合成和分泌,影响棉花对难溶性无机磷的活化利用的生物学功能[95]。与该结果相似,在苜蓿中超量表达neMDH基因和在烟草中异源表达菌根真菌的MDH基因也显著提高了苹果酸浓度和分泌量,进一步说明了MDH参与调控苹果酸合成和分泌的功能[96-97]。与MDH基因相似,CS基因的表达水平也影响了植物柠檬酸的合成和分泌,从而参与调控植物活化利用难溶性无机磷的能力。在烟草中超量表达CS基因导致柠檬酸合成酶的活性提高约2倍,柠檬酸的分泌量提高2~4倍[98]。而且,与野生型相比,转基因株系对难溶性无机磷的活化利用能力明显增强[98]。在油菜和拟南芥的研究中也获得了类似的结果,即超量表达CS基因,显著提高了转基因材料内源柠檬酸含量与根系柠檬酸的分泌量,从而分别促进了油菜对Fe-P和拟南芥对Al-P的活化利用能力[99-101]。这些研究结果表明,植物MDH 和CS 可以分别调控植物苹果酸和柠檬酸的合成,从而控制有机酸的分泌和植物对难溶性无机磷活化利用的能力。
3.3.2 低磷调控有机酸转运的分子机制
根系苹果酸的分泌主要通过铝激活型苹果酸转运蛋白(Aluminum-activated malate transporter, ALMT)所介导[102]。植物苹果酸转运子的蛋白结构比较保守,在其N端通常含有5~7个跨膜结构域。但是,其C端的保守性较低,部分成员含有一段较长的跨膜结构域,部分成员则为亲水氨基酸序列[103-106]。虽然,关于ALMT控制植物根系分泌苹果酸的研究较多,但主要是关于苹果酸分泌与植物耐铝毒害的研究。其中,小麦TaALMT1是第1个在植物中克隆到的、编码苹果酸转运蛋白的基因[107]。超量表达该基因显著增加了小麦、大麦和烟草悬浮细胞苹果酸分泌及其耐铝毒害的能力[107-108]。随后,在拟南芥、油菜、苜蓿和大豆等植物研究中,也发现其同源基因具有类似的功能,说明了ALMT参与调控植物根系分泌苹果酸的功能[109-114]。最近,对大麦、橙Citrus sinensis和大豆等植物的研究也表明,苹果酸转运子参与调控了植物对低磷胁迫的响应[92, 115-116]。例如,在大麦中,Delhaize等[115]通过超量表达TaALMT1基因提高酸性土壤中转基因大麦的生物量及磷含量,说明TaALMT1通过调控苹果酸的分泌,促进大麦根系的生长和磷吸收。在大豆根系的研究中发现,34个GmALMT家族成员中仅有4个成员的表达量显著受低磷胁迫加强[116]。而且,超量表达GmALMT5不仅提高了转基因材料根系苹果酸的分泌量,而且促进了转基因拟南芥对难溶性Ca-P的活化和利用,说明ALMT介导的根系苹果酸分泌是植物适应低磷胁迫的重要机制。
与苹果酸的分泌不同,植物根系柠檬酸的分泌主要是由Multidrug and toxic compound extrusion (MATE)家族成员所控制[117]。虽然,在多种植物中已克隆了MATE的同源基因,但目前的研究主要集中在植物MATE调控植物适应缺铁胁迫或铝毒害方面[117],而MATE参与植物适应低磷胁迫的机制鲜有报道。最近Valentinuzzi等[118]在草莓Fragaria×ananassa中发现,2个MATE基因的表达水平受低磷上调,暗示MATE可能介导低磷调控根系柠檬酸分泌的机制。
4. 展望
根系分泌物是根际微生态环境的重要组成部分,也是植物根系–土壤–根际微生物产生互作的主要桥梁。以往的研究表明,低磷胁迫下有机酸和紫色酸性磷酸酶的分泌显著影响了根际磷营养的状况,充分说明了根系分泌物调控植物适应低磷胁迫的机制。而且,随着对根际微生物的深入研究,发现植物根系分泌物显著调控了根际微生物群落的组成和活性[119-120]。因此,低磷胁迫下根际微生物群落组成和活性是否发生改变?这些变化是否与根系分泌物(有机酸和酸性磷酸酶)的组成和含量有关?这些科学问题的解答,将有助于全面解析根系分泌物对植物适应低磷胁迫的生物学意义。
-
![]()
图 1 水稻颖花结构及籼、粳稻花时对比
a:水稻小穗,Bar = 0.5 cm,b:水稻颖花结构图,Bar = 1.0 mm;c:粳稻‘中花11’与籼稻‘天丰B’在09:30的开花对比图,Bar = 1.0 cm,d:粳稻‘中花11’与籼稻‘天丰B’在11:00的开花对比图,Bar = 1.0 cm
Figure 1. The rice floret structure and the comparison of floret-opening-time between indica and japonica
a: Rice spikelet, bar = 0.5 cm; b: The structure of a rice floret, bar = 1.0 mm; c: Comparison of flowering between japonica ‘Zhonghua 11’ and indica ‘Tianfeng B’ at 09:30, bar = 1.0 cm; d: Comparison of flowering between japonica ‘Zhonghua 11’ and indica ‘Tianfeng B’ at 11:00, bar = 1.0 cm
![]()
图 2 水稻开花前后不同时间点浆片吸水膨胀与失水萎缩过程
a:水稻浆片形态,Bar = 1 mm;b:水稻颖花示意图;c:b图红色虚线对应的横切示意图,红色箭头表示浆片膨胀时张力方向
Figure 2. The process of lodicule swelling and shrinking in rice before and after floret-opening
a: Dynamic morphology change of the rice lodicules during anthesis, bar = 1 mm; b: Schematic diagrams of rice florets during anthesis; c: Schematic diagrams of the cross-sections of florets as the red dash lines indicates in Fig. 2b, the red arrow indicates the direction of tension when the lodicule expanding
![]()
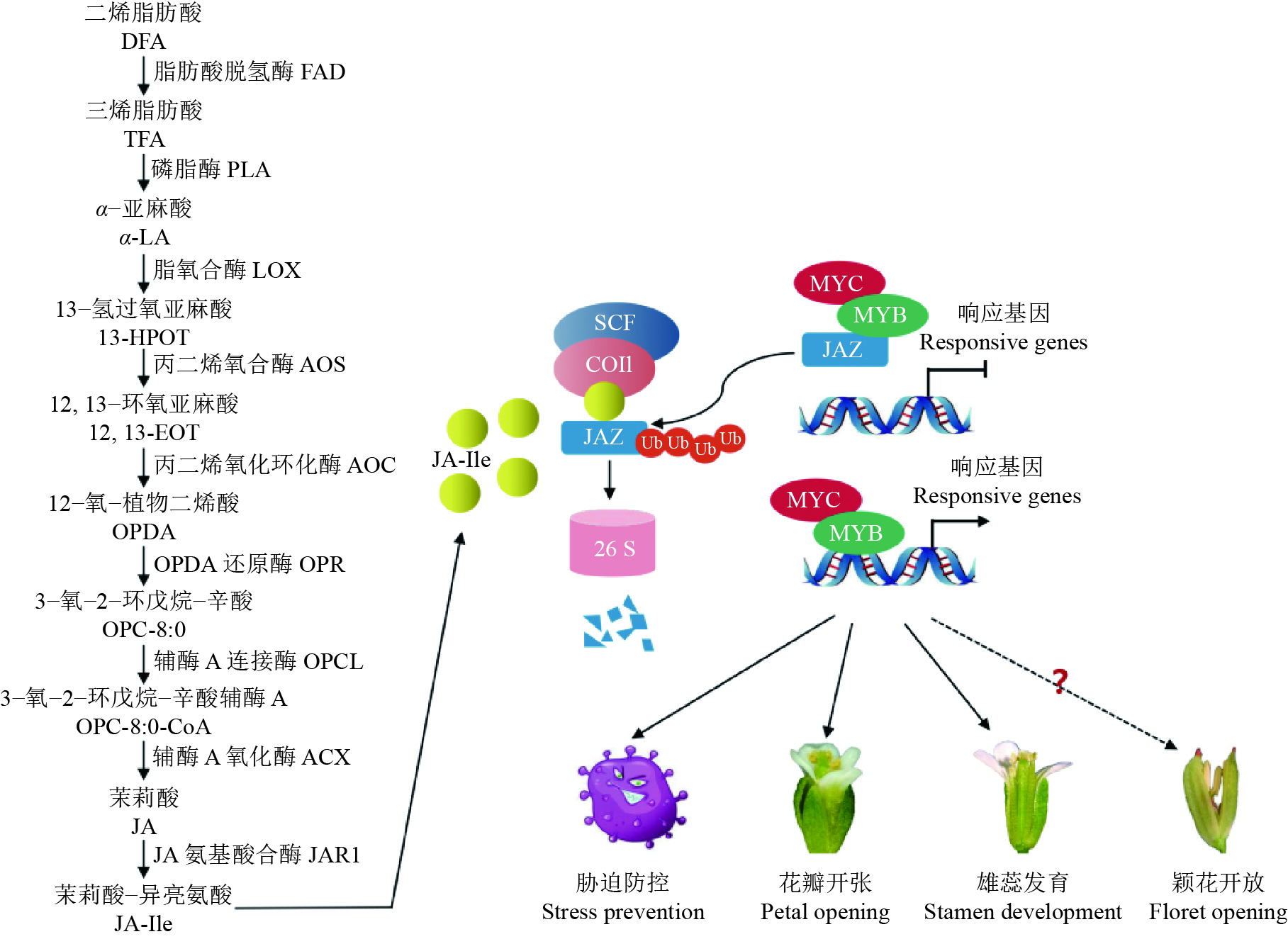
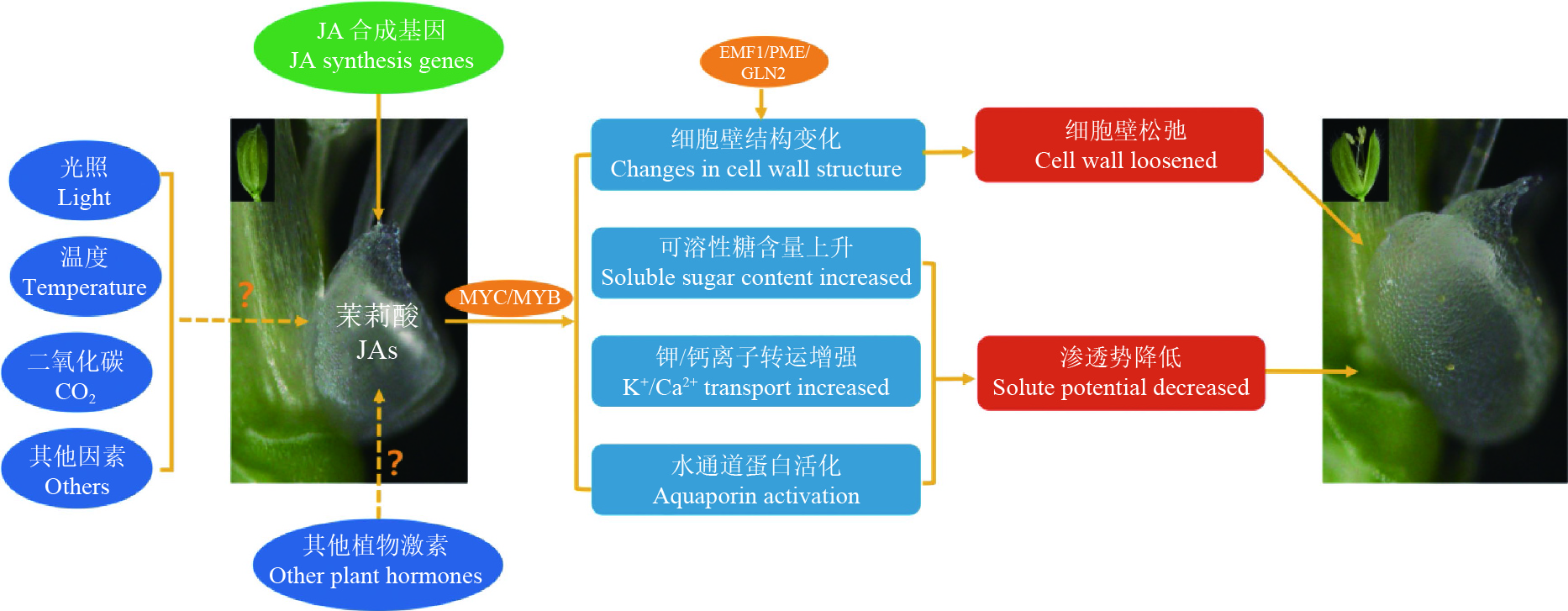
图 3 茉莉酸合成、信号转导及生物学功能[46]
Figure 3. Jasmonic acid synthesis, signal transduction and biological function
表 1 花时QTLs的详细信息
Table 1 The details of floret-opening-time QTLs
数量性状 基因座 QTL 亲本1)Parent 性状 Trait 染色体 Chromosome 定位区间 Mapping location 基因来源 Gene source 贡献率/% Contribution rate 文献 Reference qFBT-12 七山占 × 秋光 始花时 12 PSM420—RM247 秋光 10.33 [7] qVFT-1 七山占 × 秋光 盛花时 1 RM259—RM449 秋光 12.43 [7] qVFT-8 七山占 × 秋光 盛花时 8 RM477—PSM396 秋光 10.57 [7] qVFT-10 七山占 × 秋光 盛花时 10 RM269—RM222 七山占 26.95 [7] qFET-2 七山占 × 秋光 终花时 2 RM240—RM525 秋光 7.08 [7] qFET-10 七山占 × 秋光 终花时 7 RM82—RM420 七山占 10.11 [7] SOTb W630 × 日本晴 始花时 5 RM249—RM440 W630 27.21 [34] SOTb W630 × 日本晴 始花时 10 RM171—RM496 W630 28.55 [34] SOTm W630 × 日本晴 盛花时 4 RM303—RM255 W630 14.66 [34] SOTm W630 × 日本晴 盛花时 5 RM249 W630 20.50 [34] qFT1a WAB368-B-2-H2-HB × 六千辛 盛花时 1 RM580—RM8004 WAB368-B-2-H2-HB 7.50 [35] qFT1b WAB368-B-2-H2-HB × 六千辛 盛花时 1 RM7318—RM5497 WAB368-B-2-H2-HB 5.80 [35] qFT10 WAB368-B-2-H2-HB × 六千辛 盛花时 10 RM184—RM3773 WAB368-B-2-H2-HB 6.20 [35] qFT12 WAB368-B-2-H2-HB × 六千辛 盛花时 12 RM511—RM519 WAB368-B-2-H2-HB 11.30 [35] qEMF3 Oryza officinalis × 南京11 始花时 3 RM14407 O. officinalis 19.70 [36] 1)籼稻:‘七山占’‘南京11’,粳稻:‘秋光’‘日本晴’‘WAB368-B-2-H2-HB’‘六千辛’,野生稻:‘W630’、Oryza officinalis 1) Indica: ‘Qishanzhan’ ‘Nanjing11’; Japonica: ‘Qiuguang’ ‘Ribenqing’ ‘WAB368-B-2-H2-HB’ ‘Liuqianxin’; Wild rice: ‘W630’, Oryza officinalis  下载: 导出CSV
下载: 导出CSV
-
[1] FITZGERALD M A, MCCOUCH S R, HALL R D. Not just a grain of rice: The quest for quality[J]. Trends in Plant Science, 2009, 14(3): 133-139. doi: 10.1016/j.tplants.2008.12.004
[2] PENG S, KHUSH G S, VIRK P, et al. Progress in ideotype breeding to increase rice yield potential[J]. Field Crops Research, 2008, 108(1): 32-38. doi: 10.1016/j.fcr.2008.04.001
[3] CHENG S H, ZHUANG J Y, FAN Y Y, et al. Progress in research and development on hybrid rice: A super-domesticate in China[J]. Annals of Botany, 2007, 100(5): 959-966. doi: 10.1093/aob/mcm121
[4] 孙凌飞, 李绍波, 官杰, 等. 亚洲栽培稻的籼粳分化[J]. 现代农业科技, 2008(22): 157-159. [5] 张桂权. 5G水稻的演变和发展[J]. 华南农业大学学报, 2019, 40(5): 211-216. [6] 张萌, 戴冬青, 李西明, 等. 水稻花时性状研究进展[J]. 核农学报, 2016, 30(2): 267-274. [7] 马作斌, 詹瞻, 徐海, 等. 籼粳稻杂交后代花时性状的QTL分析[J]. 植物生理学报, 2011, 47(8): 799-802. [8] YANG J, FEI K, CHEN J, et al. Jasmonates alleviate spikelet-opening impairment caused by high temperature stress during anthesis of photo-thermo-sensitive genic male sterile rice lines[J]. Food and Energy Security, 2020, 9(4): e233.
[9] 徐乾坤, 任德勇, 李自壮, 等. 水稻小穗颖壳发育的研究进展[J]. 中国水稻科学, 2016, 30(1): 99-105. [10] TANAKA W, TORIBA T, HIRANO H Y. Flower development in rice[J]. Advances in Botanical Research, 2014, 72(8): 221-262.
[11] 黄俊宝, 何永明, 曾晓春, 等. 水稻颖花开放前花器官茉莉酸水平变化及浆片茉莉酸信号基因表达分析[J]. 中国农业科学, 2015, 48(6): 1219-1227. [12] KOBAYASI K, MATSUI T, YOSHIMOTO M, et al. Effects of temperature, solar radiation, and vapor-pressure deficit on flower opening time in rice[J]. Plant Production Science, 2010, 13(1): 21-28. doi: 10.1626/pps.13.21
[13] 田大成. 水稻异交栽培学: 杂交水稻高产制种原理与技术[M]. 成都: 四川科学技术出版社, 1991. [14] 曾晓春. 茉莉酸类对稻、高梁和果园草颖花开放的诱导效应[D]. 南京: 南京农业大学, 2000. [15] 仲维功, 李传国, 苏自强. 不同类型水稻品种花时特性的研究[J]. 江苏农业科学, 1990(1): 16-18. [16] 陆燕雯, 王冬翼, 顾庆华, 等. 粳型光温敏感雄性核不育系开花习性研究[J]. 上海农业学报, 2012, 28(3): 124-126. [17] 曾晓春, 周燮, 吴晓玉. 水稻颖花开放机理研究进展[J]. 中国农业科学, 2004, 37(2): 188-195. [18] 王忠, 顾蕴洁, 高煜珠. 水稻开颖机理的探讨: Ⅲ: 浆片的结构及其在开颖过程中内含物的变化[J]. 作物学报, 1991, 17(2): 96-101. [19] HESLOP-HARRISON Y, HESLOP-HARRISON J S. Lodicule function and filament extension in the grasses: Potassium ion movement and tissue specialization[J]. Annals of Botany, 1996: 573-582.
[20] 王忠, 顾蕴洁. 水稻开闭颖过程及其影响因素[EB/OL]. 中国科技论文在线, [2022-09-03]. http://www.paper.edu.cn. [21] 王忠, 顾蕴洁, 高煜珠. 水稻开颖机理的探讨: Ⅴ: 不育系与可育系浆片和花丝结构的比较[J]. 作物学报, 1994, 20(1): 13-17. [22] 薛欣艳. 关于水稻浆片调节颖花开放机理的分子生物学基础研究[D]. 扬州: 扬州大学, 2010. [23] 刘娟. 一个水稻花时提前突变体的遗传分析与基因定位[D]. 成都: 四川农业大学, 2016. [24] 邹春梅, 何永明, 曾晓春. MeJA诱导水稻颖花开放作用机理的研究[G]. 2007年全国植物生长物质研讨会论文摘要汇编. 南昌: 中国植物生理学会, 2007. [25] QIN Y, YANG J, ZHAO J. Calcium changes and the response to methyl jasmonate in rice lodicules during anthesis[J]. Protoplasma, 2005, 225(1/2): 103-112.
[26] 玉忠, 顾蕴洁, 高煜珠. 水稻开颖机理的探讨: Ⅱ: CO2对水稻开颖的效应[J]. 作物学报, 1989, 15(1): 59-66. [27] FRY S C, SMITH R C, RENWICK K F, et al. Xyloglucan endotransglycosylase, a new wall-loosening enzyme activity from plants[J]. The Biochemical Journal, 1992, 282(Pt 3): 821-828.
[28] CARPITA N C, GIBEAUT D M. Structural models of primary cell walls in flowering plants: Consistency of molecular structure with the physical properties of the walls during growth[J]. The Plant Journal, 1993, 3(1): 1-30. doi: 10.1111/j.1365-313X.1993.tb00007.x
[29] 付永琦, 向妙莲, 蒋海燕, 等. 水稻颖花开放前浆片转录组变化[J]. 中国农业科学, 2016, 49(6): 1017-1033. [30] SAKURAI J, ISHIKAWA F, YAMAGUCHI T, et al. Identification of 33 rice aquaporin genes and analysis of their expression and function[J]. Plant and Cell Physiology, 2005, 46(9): 1568-1577. doi: 10.1093/pcp/pci172
[31] 何永明, 曾晓春, 向妙莲, 等. 水稻花时调控研究进展[J]. 湖北农业科学, 2014, 53(7): 1489-1492. [32] ZENG X C, ZHOU X, ZHANG W, et al. Opening of rice floret in rapid response to methyl jasmonate[J]. Journal of Plant Growth Regulation, 1999, 18(4): 153-158. doi: 10.1007/PL00007063
[33] 李金军, 范国华, 张仁余, 等. 不同水稻品种开花时间的比较试验[J]. 浙江农业科学, 2007(1): 63-66. [34] PHAM T T, PHUONG D T P, ISHIKAWA R, et al. QTL analysis for flowering time using backcross population between Oryza sativa Nipponbare and O. rufipogon[J]. Genes & Genetic Systems, 2010, 85(4): 273-279.
[35] 万国, 冯跃, 张凤娇, 等. 水稻花时性状的QTL定位[J]. 核农学报, 2013, 27(5): 562-567. [36] HIRABAYASHI H, SASAKI K, KAMBE T, et al. qEMF3, a novel QTL for the early-morning flowering trait from wild rice, Oryza officinalis, to mitigate heat stress damage at flowering in rice, O. sativa[J]. Journal of Experimental Botany, 2015, 66(5): 1227-1236. doi: 10.1093/jxb/eru474
[37] SHEEHY J, ELMIDO A, CENTENO G, et al. Searching for new plants for climate change[J]. Journal of Agricultural Meteorology, 2005, 60(5): 463-468. doi: 10.2480/agrmet.463
[38] SHEEHY J E, MABILANGAN A E, DIONORA M J A, et al. Time of day of flowering in wild species of the genus Oryza[J]. International Rice Research Notes, 2007, 32(1): 12-13.
[39] XIAO H, WANG Y, LIU D, et al. Functional analysis of the rice AP3 homologue OsMADS16 by RNA interference[J]. Plant Molecular Biology, 2003, 52(5): 957-966. doi: 10.1023/A:1025401611354
[40] NAGASAWA N, MIYOSHI M, SANO Y, et al. SUPERWOMAN1 and DROOPING LEAF genes control floral organ identity in rice[J]. Development, 2003, 130(4): 705-718. doi: 10.1242/dev.00294
[41] YADAV S R, PRASAD K, VIJAYRAGHAVAN U. Divergent regulatory OsMADS2 functions control size, shape and differentiation of the highly derived rice floret second-whorl organ[J]. Genetics, 2007, 176(1): 283-294. doi: 10.1534/genetics.107.071746
[42] YAO S G, OHMORI S, KIMIZU M, et al. Unequal genetic redundancy of rice PISTILLATA orthologs, OsMADS2 and OsMADS4, in lodicule and stamen development[J]. Plant and Cell Physiology, 2008, 49(5): 853-857. doi: 10.1093/pcp/pcn050
[43] WANG M, ZHU X, PENG G, et al. Methylesterification of cell-wall pectin controls the diurnal flower-opening times in rice[J]. Molecular Plant, 2022, 15(6): 956-972. doi: 10.1016/j.molp.2022.04.004
[44] XU P, WU T, ALI A, et al. EARLY MORNING FLOWERING 1 (EMF1) regulates the floret opening time by mediating lodicule cell wall formation in rice[J]. Plant biotechnology journal, 2022, 20: 1441-1443. doi: 10.1111/pbi.13860
[45] ZHAI Q, ZHANG X, WU F, et al. Transcriptional mechanism of jasmonate receptor COI1-mediated delay of flowering time in Arabidopsis[J]. Plant Cell, 2015, 27(10): 2814-2828.
[46] SONG S, QI T, HUANG H, et al. Regulation of stamen development by coordinated actions of jasmonate, auxin, and gibberellin in Arabidopsis[J]. Molecular Plant, 2013, 6(4): 1065-1073. doi: 10.1093/mp/sst054
[47] WASTERNACK C, STRNAD M. Jasmonates are signals in the biosynthesis of secondary metabolites: Pathways, transcription factors and applied aspects: A brief review[J]. New Biotechnology, 2019, 48: 1-11. doi: 10.1016/j.nbt.2017.09.007
[48] CHINI A, FONSECA S, FERNÁNDEZ G, et al. The JAZ family of repressors is the missing link in jasmonate signaling[J]. Nature, 2007, 448(7154): 666-671. doi: 10.1038/nature06006
[49] THINES B, KATSIR L, MELOTTO M, et al. JAZ repressor proteins are targets of the SCF(COI1) complex during jasmonate signaling[J]. Nature, 2007, 448(7154): 661-665. doi: 10.1038/nature05960
[50] KATSIR L, CHUNG H S, KOO A J, et al. Jasmonate signaling: A conserved mechanism of hormone sensing[J]. Current Opinion in Plant Biology, 2008, 11(4): 428-435. doi: 10.1016/j.pbi.2008.05.004
[51] GEERINCK J, PAUWELS L, DE JAEGER G, et al. Dissection of the one-MegaDalton JAZ1 protein complex[J]. Plant Signaling & Behavior, 2014, 5(8): 1039-1041.
[52] WANG C, LIU Y, LI S S, et al. Insights into the origin and evolution of the plant hormone signaling machinery[J]. Plant Physiology, 2015, 167(3): 872-886. doi: 10.1104/pp.114.247403
[53] 宋平, 夏凯, 吴传万, 等. 雄性不育和可育水稻开颖对茉莉酸甲酯响应的差异[J]. 植物学报, 2001, 43(5): 480-485. [54] 闫芝芬, 周燮, 马春红, 等. 冠毒素和茉莉酸甲酯对诱导小麦、黑麦和高羊茅草颖花开放的效应[J]. 中国农业科学, 2001, 34(3): 334-337. [55] 何永明, 林拥军, 曾晓春. 水稻颖花自然开放过程中茉莉酸(JA)生物合成的变化[J]. 作物学报, 2012, 38(10): 1891-1899. [56] XIAO Y, CHEN Y, CHARNIKHOVA T, et al. OsJAR1 is required for JA-regulated floret opening and anther dehiscence in rice[J]. Plant Molecular Biology, 2014, 86(1/2): 19-33.
[57] LI X, WANG Y, DUAN E, et al. OPEN GLUME1: A key enzyme reducing the precursor of JA, participates in carbohydrate transport of lodicules during anthesis in rice[J]. Plant Cell Reports, 2018, 37(2): 329-346. doi: 10.1007/s00299-017-2232-y
[58] HIBARA K I, ISONO M, MIMURA M, et al. Jasmonate regulates juvenile-adult phase transition in rice[J]. Development, 2016, 143(18): 3407-3416.
[59] 闫志强, 徐海, 马作斌, 等. 籼稻与粳稻花时对茉莉酸甲酯(MeJA)响应的敏感性差异[J]. 中国农业科学, 2014, 47(13): 2529-2540. [60] 黄友明, 曾晓春. 环境因子和颖花构造对水稻颖花关闭的效应[J]. 江苏农业科学, 2021, 49(19): 94-100. [61] LIU L, ZOU Z, QIAN K, et al. Jasmonic acid deficiency leads to scattered floret opening time in cytoplasmic male sterile rice Zhenshan 97A[J]. Journal of Experimental Botany, 2017, 68(16): 4613-4625. doi: 10.1093/jxb/erx251
[62] 杨天玲. 提高杂交水稻繁殖制种异交结实率的途经[J]. 种子科技, 2007(3): 51-53. [63] 夏原野, 杜志敏, 杨宇尘, 等. 喷施表油菜素内酯对籼稻和粳稻花时的影响[J]. 作物杂志, 2019(4): 139-147. [64] 闫志强, 徐海, 宫彦龙, 等. 籼粳稻的花时对乙烯利调控的响应及其敏感性差异[J]. 沈阳农业大学学报, 2015, 46(6): 641-647. [65] 丁超尘, 郑立平. 激素改变杂交水稻制种父本花时的试验初报[J]. 安徽农业科学, 1994, 22(4): 295-299. [66] HUANG Y, ZENG X, CAO H. Hormonal regulation of floret closure of rice (Oryza sativa)[J]. PLoS One, 2018, 13(6): e198828.
[67] ZHAO Z, ZHANG Y, LIU X, et al. A role for a dioxygenase in auxin metabolism and reproductive development in rice[J]. Developmental Cell, 2013, 27(1): 113-122. doi: 10.1016/j.devcel.2013.09.005
[68] ZHAO Z, WANG C, YU X, et al. Auxin regulates source-sink carbohydrate partitioning and reproductive organ development in rice[J]. Proceedings of the National Academy of Sciences of the United States of America, 2022, 119(36): e2121671119. doi: 10.1073/pnas.2121671119
[69] JAGADISH S V K, BAHUGUNA R N, DJANAGUIRAMAN M, et al. Implications of high temperature and elevated CO2 on flowering time in plants[J]. Frontiers in Plant Science, 2016, 7: 913. doi: 10.3389/fpls.2016.00913.
[70] 蒯建敏, 莫惠栋, 惠大丰. 水稻花时与气象因子的关系[J]. 中国水稻科学, 1994, 8(2): 79-84. [71] 王忠, 卢从明, 顾蕴洁, 等. 水稻开颖机理的探讨: Ⅰ: 温度对水稻开颖及花粉生活力的影响[J]. 作物学报, 1988, 14(1): 14-21. [72] JAGADISH S, CRAUFURD P, WHEELER T. High temperature stress and spikelet fertility in rice (Oryza sativa L.)[J]. Journal of Experimental Botany, 2007, 58(7): 1627-1635. doi: 10.1093/jxb/erm003
[73] BISWAS K K O C, NEUMANN R, HAGA K, et al. Photomorphogenesis of rice seedlings: A mutant impaired in phytochrome-mediated inhibition of coleoptile growth[J]. Plant and Cell Physiology, 2003, 44(3): 242-254. doi: 10.1093/pcp/pcg040
[74] NISHIYAMA I, BLANCO L. Artificial control of flower opening time during the day in rice plant: I: Preliminary experiments[J]. Japanese Journal of Crop Science, 1981, 50(1): 59-66. doi: 10.1626/jcs.50.59
[75] 顾蕴洁, 王忠, 高煜珠. 环境因素对水稻颖花开闭影响的机理[J]. 植物生理学报, 1993, 19(4): 345-352. [76] 王忠, 何循宏. CO2促进小麦开花的效应[J]. 江苏农学院学报, 1991, 12(1): 46. [77] 许祥明, 王忠. 某些酸类物质对水稻开颖的效应[J]. 植物生理学报, 1998, 24(2): 124-130. [78] KOBAYASI K, SAKAI H, TOKIDA T, et al. Effects of free-air CO2 enrichment on flower opening time in rice[J]. Plant Production Science, 2019, 22(3): 367-373. doi: 10.1080/1343943X.2019.1569472
[79] 王忠, 顾蕴洁, 高煜珠. CO2诱导水稻开花技术的应用[J]. 植物生理学报, 1993, 29(4): 282-287. [80] ISHIMARU T, HIRABAYASHI H, IDA M, et al. A genetic resource for early-morning flowering trait of wild rice Oryza officinalis to mitigate high temperature-induced spikelet sterility at anthesis[J]. Annals of Botany, 2010, 106(3): 515-520. doi: 10.1093/aob/mcq124
[81] 徐伟东, 蔡金洋, 杨尧城. 水稻籼粳亚种间杂种优势利用研究现状与展望[J]. 中国稻米, 2016, 22(2): 1-7. doi: 10.3969/j.issn.1006-8082.2016.02.001 -
期刊类型引用(30)
1. 张捷,崔健,舒军,吴年隆,李梦晴,赵雄伟. 低磷胁迫下不同耐受型谷子的代谢组学差异. 植物营养与肥料学报. 2025(01): 144-155 .  百度学术
百度学术
2. 王潇,王艺雄,季杭翔,施曼,王会来,宋新章,李全. 毛竹根际土壤磷组分对氮和生物炭添加的响应. 生态学杂志. 2025(04): 1135-1143 . 百度学术
3. 杜艺,周波,袁娜娜,姚佳妮,代金霞. 荒漠植物根际解磷细菌的筛选及抑菌和促生特性研究. 农业科学研究. 2024(01): 20-25 . 百度学术
4. 宋惠洁,吴艳,胡丹丹,胡志华,柳开楼,徐小林,张九兰. 不同磷肥用量下红壤区早稻季产量与田面水磷素动态变化. 中国土壤与肥料. 2024(02): 51-57 . 百度学术
5. 王敏,彭大榕,曾吉兴,王诗语,朱林星,郭世伟. 植物矿质营养与病害研究进展及展望. 植物营养与肥料学报. 2024(07): 1339-1353 . 百度学术
6. 李蒙,熊婷婷,朱思远,袁童瑶,张健,龚守富. 不同钾肥用量对甜瓜幼苗生长及基质特性的影响. 分子植物育种. 2024(18): 6120-6128 . 百度学术
7. 黄雨轩,游欣,张林平,吴斐,张扬,黄绍华. 根系分泌物提高土壤磷有效性研究概述. 林业科学研究. 2024(04): 193-203 . 百度学术
8. 安海涛,孙彩彩,董全民,杨晓霞,刘文亭,赵新全. 根系分泌物与植物—土壤间互作机制的研究进展. 青海畜牧兽医杂志. 2024(05): 50-59 . 百度学术
9. 薛迎斌,宋佳,李枭艺,李小豪,陈经烨,伍萍珍,朱胜男,刘颖. 大豆GmMADS4基因克隆、亚细胞定位及功能分析. 华南农业大学学报. 2023(03): 420-429 . 本站查看
10. 孙志伟,徐月梅,许荣越,朱宽宇,杨建昌. 水稻低磷胁迫响应及其调控机制的研究进展. 核农学报. 2023(08): 1562-1570 . 百度学术
11. 刘瑀,陆超,焦点,刘宇馨,王国光. 低磷胁迫下翅碱蓬生长特性及脂肪酸含量的变化机制. 海洋科学. 2023(04): 117-125 . 百度学术
12. 吕铭滔,龚海光,黄永芳,龚勇军. 磷对铝胁迫油茶芽苗初生根保护酶的影响. 经济林研究. 2023(03): 271-277 . 百度学术
13. 陈浩婷,张玉静,刘洁,代泽敏,刘伟,石玉,张毅,李天来. 低磷胁迫下番茄转录因子WRKY6功能分析. 生物技术通报. 2023(10): 136-147 . 百度学术
14. 夏雪,蔡康锋,刘磊,宋秀娟,汪军妹,岳文浩. 大麦根形态和分子水平对低磷胁迫响应研究进展. 作物杂志. 2023(06): 11-16 . 百度学术
15. 田磊. 不同林龄泡桐根系分泌物对土壤酶活性和微生物的影响. 林业调查规划. 2022(01): 28-33 . 百度学术
16. 莫维维,韦建圩,梁嘉玲,刘易,谢伟东,任哲. 桂西北喀斯特地区漾濞泡核桃林地土壤与叶片养分化学计量特征. 经济林研究. 2022(01): 26-35 . 百度学术
17. 赵宽,万昕,邢德科,胡睿鑫,周葆华,袁可升. 低分子量有机酸对土壤有效磷及重金属释放影响的研究进展. 土壤通报. 2022(05): 1228-1236 . 百度学术
18. 蔡银美,赵庆霞,张成富. 低磷下植物根系分泌物对土壤磷转化的影响研究进展. 东北农业大学学报. 2021(02): 79-86 . 百度学术
19. 马庆华,贺淑霞,王伟南,曹晗,李贺. 磷供应水平对盆花月季‘仙境’生长和品质的影响. 北方园艺. 2021(11): 69-75 . 百度学术
20. 周梦岩,何冬梅,李亚超,刘静,马祥庆,李明. 紫色酸性磷酸酶在植物响应低磷胁迫中的作用研究进展. 分子植物育种. 2021(11): 3763-3770 . 百度学术
21. 惠乾龙,叶文彬,郭晋隆,袁照年,许莉萍. 植物磷匮乏下的根系、代谢和分子响应研究进展. 中国糖料. 2021(03): 34-42 . 百度学术
22. 靳琇,陈浩婷,石玉,白龙强,侯雷平,张毅. 柠檬酸浸种引发对低磷胁迫下番茄幼苗生长及生理特性的影响. 中国生态农业学报(中英文). 2021(07): 1159-1170 . 百度学术
23. 沈启维,李艳春,张健. 提高植物磷高效利用能力方法的研究进展. 绿色科技. 2021(11): 157-160 . 百度学术
24. 肖如武,黄楚龙,宗钊辉,王维,赵伟才,王军. 低磷胁迫对烤烟根系有机酸含量及土壤磷酸酶活性的影响. 广东农业科学. 2021(08): 74-82 . 百度学术
25. 忙顺兰,罗晓蔓,丁贵杰. 马尾松幼苗根系分泌物对土壤酶活性和养分的影响. 中南林业科技大学学报. 2021(12): 53-59 . 百度学术
26. 丁艳,邢媛,任蒙莲. 低磷水稻根表铁膜形成对养分吸收的影响. 安徽科技学院学报. 2021(05): 47-52 . 百度学术
27. 武海燕,李喜焕,李文龙,孔佑宾,杜汇,张彩英. 大豆耐低磷性状鉴定及优异种质筛选. 河南农业科学. 2020(01): 61-67 . 百度学术
28. 张弛,孟琳钦,苏丹,仝则乾,胡祖庆. 麦蚜及植物病毒胁迫下小麦体内保护酶和解毒酶活性的变化. 麦类作物学报. 2020(03): 328-333 . 百度学术
29. 赵雄伟,吴年隆,乔佳辉,李旭凯,韩渊怀,邢国芳. 谷子酸性磷酸酶ACP家族基因鉴定与SiACP1耐低磷单倍型分析. 华北农学报. 2020(04): 35-45 . 百度学术
30. 王宇蕴,汤利. 多样性种植体系提高酸性红壤磷有效性的研究进展. 磷肥与复肥. 2020(09): 47-49 . 百度学术
其他类型引用(35)





计量
- 文章访问数: 214
- HTML全文浏览量: 40
- PDF下载量: 479
- 被引次数: 65
