
Pathogen isolation, identification and pathomorphological observation of ulcerative syndrome of largemouth bass in winter and spring
-
摘要:目的
近年来,养殖大口黑鲈Micropterus salmoides在冬春季流行一种体表形成明显溃疡的疫病,本研究旨在明确其病因。
方法从眉山东坡区和洪雅县采集病料,采用病原学、病理学和分子生物学相结合的方法对患病鱼进行研究。
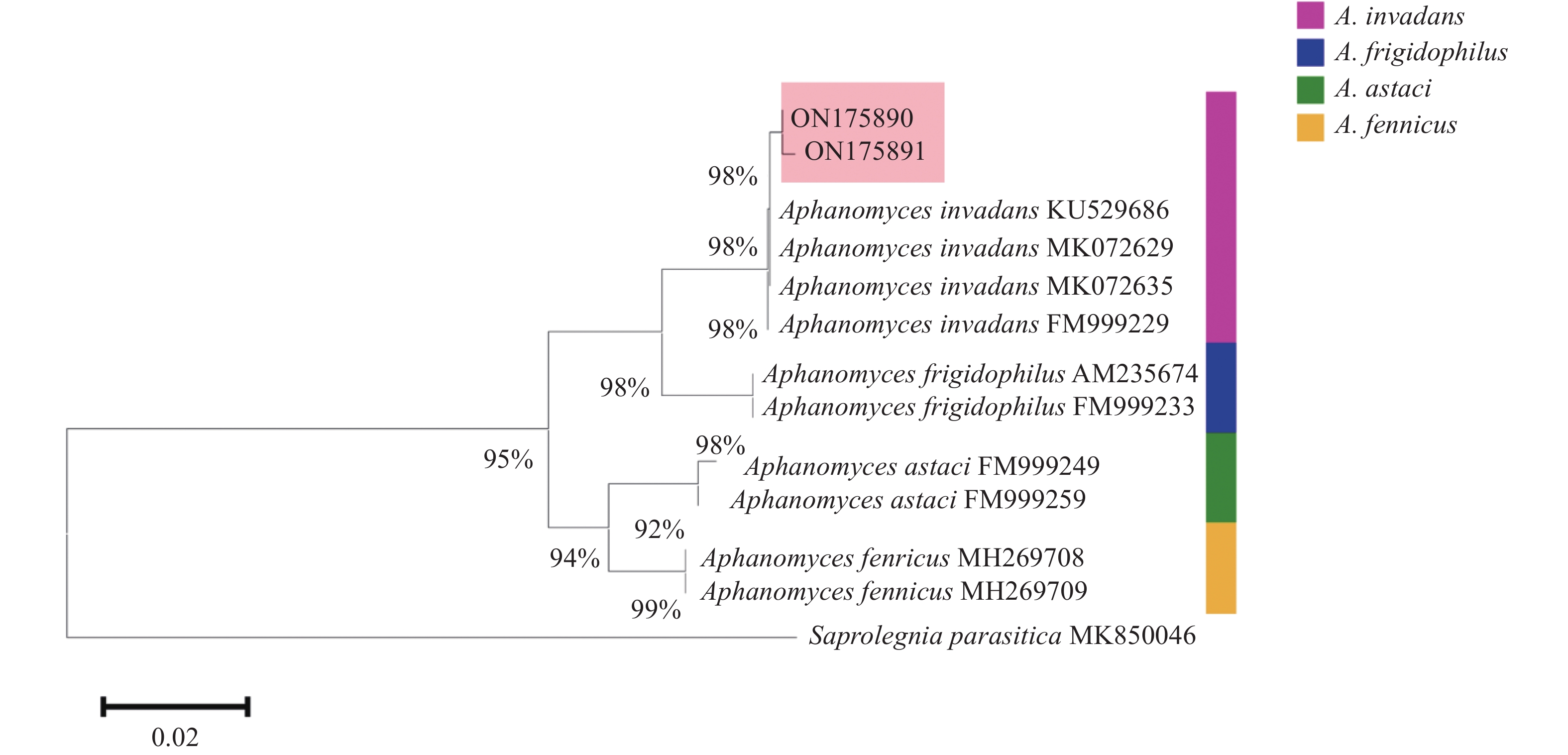
结果溃疡组织压片观察到大量直径10~30 μm、长丝状且分支较少的菌丝体;组织病理学上,肌纤坏死、溶解,炎性细胞浸润,病变组织中分布大量由上皮样细胞、纤维细胞及位于中央的菌丝体组成的霉菌性肉芽肿;六胺银染色(Periodic acid-silver metheramine,PASM)发现,霉菌性肉芽肿中央的菌丝体呈黑色。病变肌肉组织基于侵入丝囊霉菌Aphanomyces invadans ITS基因的特异性PCR检测为阳性。以葡糖糖/蛋白胨(Glucose and peotone,GP)琼脂分离培养得到的霉菌纯培养物(BW1、BW3)在灭菌池塘水过夜培养12 h后,形成原孢子囊,符合侵入丝囊霉菌属特征;两株分离霉菌ITS基因序列分析发现,其与侵入丝囊霉菌ITS基因的序列相似性分别为99.64%与99.46%,在系统发育树上与侵入丝囊霉菌聚类为一支。
结论确定大口黑鲈冬春季溃疡病是侵入丝囊霉菌感染所致。
Abstract:ObjectiveIn recent years, outbreak of an epidemic disease with significant ulcers on the pond-farmed largemouth bass (Micropterus salmoides) happen in winter and spring. This study aims to identify the underlying etiology.
MethodTo clarify its etiology, disease fish samples from Dongpo and Hongya District, Meishan were collected and analyzed using pathogenesis, pathology and molecular biology methods.
ResultA large number of long filamentous and less-branched mycelia with 10−30 μm diameter were observed in the ulcerated tissues. Pathologically, we observed necrosis or lysed muscle fibers, inflammatory cell infiltration in the muscle tissue, and distribution of numerous fungal granulomas consisting of epithelioid cells, fibroblasts, and central located mycelia in the lesion tissues. By using periodic acid-silver metheramine (PASM) staining, the mycelia in the fungal granulomas center displayed black colour. We confirmed the infection of Aphanomyces invadans in the lesion muscle tissues by PCR using species-specific primers that target the ITS gene of A. invadans. The A. invadans (BW1 and BW3) pure cultures were isolated by glucose and peotone (GP) agar and then incubated in sterilized pond water. Prosporangium formation was observed in the culture after 12 h incubation period, which was consistent with the characteristics of Aphanomyces. Based on ITS gene sequencing of BW1 and BW3, their ITS genes were found to have 99.64% and 99.46% sequence similarities with A. invadans, and the two strains clustered with A. invadans in the phylogenetic tree.
ConclusionThe ulcer disease of largemouth bass in winter and spring has been caused by A. invadans.
-
苹果作为一种常见的水果,其质量问题直接关系到消费者的健康和生产者的经济利益。我国是苹果种植大国,苹果采摘、运送、储藏与加工处理是不可忽视的关键环节[1]。然而,由于人工质检的主观性和效率低下,传统的苹果质检方式已经无法满足现代农产品质量的需求[2]。基于机器学习的苹果识别技术应运而生,该方法可以对苹果进行自动识别和分类。李大华等[3]针对自然复杂环境下的苹果重叠问题,利用谱聚类算法进行分割,然后使用随机霍夫变换实现果实的识别和定位,尽管改进后的谱聚类算法在计算量上有所优化,但仍然涉及多步骤的处理,包括均值漂移预分割、稀疏矩阵构建、K-means分类和随机霍夫变换,整体流程较为复杂,对硬件性能要求较高,不适合实时性要求较高的应用场景。王迎超等[4]为了准确快速实现多特征融合的苹果分级,提出了一种基于K-means聚类和改进多层感知器(Multilayer perception, MLP)的苹果分级方法,但特征权重的确定存在主观性,且在不同的应用场景或苹果品种中特征的重要性会有所不同,导致分级准确性下降。宋怡焕等[5]提出基于最小二乘支持向量机(LS-SVM)检测苹果果梗/花萼缺陷,试验中仅使用了 180 幅苹果图像,而训练数据的不足会导致模型的泛化能力受限,对不同环境或苹果品种的误判率较高。上述传统的机器学习算法试验设计过于繁琐,对复杂的特征提取和分类能力不佳,易受外界因素的干扰,难以获得较好的检测结果。随着深度学习技术突飞猛进,以深度学习为基础的目标检测技术在苹果果实检测[6]、病虫害监测[7]、果实成熟度[8]和农业自动化[9-10]等领域发挥着重要的作用。目前,基于深度学习的苹果检测算法依据检测的阶段划分主要有2大类:一类是要先用特征提取器生成一系列可能包含待检物体的预选框,然后利用算法对生成的候选区域进行更精细的检测和分类的两阶段算法,主要有Faster-RCNN[11]和Mask-RCNN[12]算法;另一类为直接预测物体类别和位置的一阶段检测算法,主要有RetinaNet[13]、SSD[14]和YOLO系列[15-17]。两阶段算法计算复杂度高、检测速度较慢,一阶段算法在精度上有所欠缺。两阶段算法由于包含多个阶段和较多的候选框处理,推理速度通常比单阶段算法慢,不适合实时应用。为了进一步提升模型的检测速度和准确度,Tian等[18]提出了一种名为VMF-SSD(基于V空间的多尺度特征融合SSD)的新型苹果叶片病害检测方法;Wang等[19]通过迁移学习构建YOLOv5s检测模型的同时,采用通道修剪算法对模型进行修剪及微调,以实现对苹果果实的快速准确检测。
目前,国内外目标检测领域对苹果的研究主要集中在自然环境下的苹果识别、苹果采摘机器人以及叶片病虫害识别方面。虽然已有部分研究针对缺陷苹果的检测[20-21],但仍存在上下文信息和多尺度特征融合不充分的问题。针对缺陷苹果识别,本文提出一种基于BiFPN[22]和Triplet注意力机制的YOLOv5s (BTF-YOLOv5s)缺陷苹果识别算法,通过BiFPN结合特征的上采样和下采样路径,联合跨尺度的特征交互,能够更好地传递和融合来自不同尺度的特征信息,提升目标检测性能;同时,学习动态的特征融合权重,根据不同目标的重要性进行灵活调整,增强模型的鲁棒性;在Neck层应用Triplet注意力机制,模型能更好地捕捉图像中的细节和上下文信息,提升目标检测的精度;采用Focal-CIoU调整正负样本之间的损失权重,让模型对难以分类的样本给予更多关注,提升模型的分类性能。
1. 基于BiFPN和Triplet注意力机制的YOLOv5s算法
本研究以YOLOv5s (版本6.2,包含4种不同大小的模型)为基础,网络结构由输入端、骨干网络、颈部网络和输出端4个部分组成。在输入端引入了Mosaic数据增强方法,随机选取4张图像,对它们进行随机放大、缩小等操作后,拼接成一张新的图像,使模型可以在更小的范围内识别目标;骨干网络主要使用了Focus和CSP1_X结构;颈部网络采用FPN+PAN和CSP2_X结构进一步进行特征融合和上采样操作,以提供更高级的语义信息和适应不同尺度图片的能力;输出端采用CIoU_Loss(Complete intersection over union loss)作为损失函数,并使用了非极大值抑制(Non-maximum suppression,NMS)进行后处理。本文提出了一种改进的YOLOv5s算法(BTF-YOLOv5s)用于缺陷苹果识别,首先,加权双向特征金字塔网络(Bidirectional feature pyramid network,BiFPN)替代YOLOv5中的FPN+PAN结构,通过引入可学习的权重学习不同输入特征的重要性;然后,将Triplet注意力机制应用到模型的最后一层,确保在最终输出前对特征进行充分的处理和优化;最后,采用Focal-CIoU损失函数调整损失权重给予缺陷苹果更多的关注,BTF-YOLOv5s算法结构整体框图如图1所示。
1.1 BiFPN
在目标检测任务中,有效地获取并处理不同尺度的特征信息是一个主要的挑战。传统的特征金字塔网络(FPN)[23]通过自上而下的方式聚合多尺度特征,如图2a所示,但容易受到单向信息流的限制;路径聚合网络(PANet)[24]在此基础上额外添加了一个自下向上的路径聚合网络,如图2b所示。在YOLOv5的Neck结构中,借鉴PANet的思想,高层的特征信息通过FPN+PAN结构进行传递融合。FPN+PAN结构虽然提高了特征传递的效率,但也增加了计算复杂度,尤其是在处理高分辨率输入时,可能导致较高的计算成本和较差的实时性;此外,固定结构导致对不同任务和数据缺乏自适应的能力。
为了解决上述信息流动单一、精度高但参数较多、计算量较大以及简单拼接导致的信息丢失和冗余等问题,本文提出了采用BiFPN替代YOLOv5s中的FPN+PAN结构。BiFPN在PANet和NAS-FPN(图2c)基础上优化了多尺度特征融合方式,结构如图2d所示。
传统方法在融合具有不同分辨率的特征时,平等地对待所有输入特征,并将它们简单相加。但不同分辨率的输入特征对输出特征的贡献通常是不等的。为了解决这一问题,BiFPN为每个输入特征增加了一个额外的权重,并让网络学习每个输入特征的重要性,具体见式(1)和式(2):
$$ P_6^{{\mathrm{td}}} = {\mathrm{Conv}}\left[ {\frac{{{w_1}P_6^{{\mathrm{in}}} + {w_2}{\mathrm{Reseize}}\left( {P_7^{{\mathrm{in}}}} \right)}}{{{w_1} + {w_1} + \varepsilon }}} \right], $$ (1) $$ P_6^{{\mathrm{out}}} = {\mathrm{Conv}}\left[ {\frac{{w_1^{'}P_6^{{\mathrm{in}}} + w_2^{'}P_6^{{\mathrm{td}}} + w_3^{'}{\mathrm{Reseize}}\left( {P_5^{{\mathrm{out}}}} \right)}}{{w_1^{'} + w_2^{'} + w_3^{'} + \varepsilon }}} \right], $$ (2) 式中,
$ P_6^{{\mathrm{td}}}$ 为自顶向下路径第6层的中间特征;$ P_6^{{\mathrm{in}}}$ 为第6层的输入特征; Resize用于分辨率匹配的上采样或下采样运算;$ P_6^{{\mathrm{out}}}$ 为自底向上路径第6层的输出特征。这一改进使得网络能够自适应地调整每个特征图的重要性,从而实现更有效的特征融合。引入BiFPN的YOLOv5s模型通过多尺度特征融合和简化的计算过程,有效地提升了对缺陷苹果检测的准确率;BiFPN通过引入可学习的融合权重优化了不同分辨率特征的重要性,增强了特征复用和多级特征融合路径,使得YOLOv5能够有效地检测出不同大小和形状的苹果缺陷。
1.2 Triplet注意力机制
在缺陷苹果检测任务中,模型需要准确地识别和定位图像中的目标(苹果)。图像中存在大量的背景信息以及不同大小的苹果会分散模型的注意力,使得模型难以准确地区分缺陷苹果和背景,引入注意力机制旨在提升模型对关键目标的感知能力。SENet通过简单的操作(全局平均池化和全连接层)自适应地调整特征图中每个通道的重要性,使网络更加集中地关注重要的特征信息,在低计算成本的条件下通过通道注意力机制显著提升网络的表示能力[25];CBAM将通道注意力和空间注意力相结合,能够同时关注重要的通道特征和空间位置;CBAM中的通道注意力方法虽然提供了一定的性能改进,但通道注意力和空间注意力是相互分离和计算的[26]。
Triplet 注意力机制(简称“Triplet”)以一种有效的方式解释了CBAM未考虑到的跨维度的相互作用[27]。如图3所示,Triplet由3个平行的分支构成,其中2个分支分别用来捕获通道
$C$ 维度和空间维度$ W/H $ 之间的跨通道交互,最后的一个分支用于捕获$ H $ 和$W$ 的空间依赖关系。在第1个分支中,输入张量C×H×W(通道数$C$ 、高度$H$ 和宽度$W$ )沿$H$ 轴逆时针旋转90°,形状变为$W$ ×$H$ ×$C$ ,输入特征先后经过$ Z - {\mathrm{Pool}} $ 、$ K \times K $ 的标准卷积层、批量归一化层、通过Sigmod激活函数生成空间注意力权重后、再沿H轴顺时针旋转90°保持与输入的形状一致。其中$ Z - {\mathrm{Pool}} $ 表示为:$$ Z - {\mathrm{Pool}}(\chi ) = \left[{\mathrm{Max}}{{\mathrm{Pool}}_{0{\mathrm{d}}}}(\chi ),{\mathrm{Avg}}{{\mathrm{Pool}}_{0{\mathrm{d}}}}(\chi )\right]{\text{,}} $$ (3) 式中,
$ 0{\mathrm{d}}$ 表示发生最大池化和平均池化操作的第0维度。第2、第3个分支同理,最后对3个分支输出特征进行平均后聚合在一起,最终输出的张量:
$$\begin{split} y =& \frac{1}{3}\Biggr\{ \overline {\mathop {{\chi _1}}\limits^ \wedge \sigma \left[ {{\psi _1}\left( {\mathop {\chi _1^*}\limits^ \wedge } \right)} \right]} + \overline {{{\mathop \chi \limits^ \wedge }_2}\sigma \left[ {{\psi _2}\left( {\mathop {\chi _2^*}\limits^ \wedge } \right)} \right]} + \Biggr.\\&\Biggr.\chi \sigma \left[ {{\psi _3}\left( {\mathop {{\chi _3}}\limits^ \wedge } \right)} \right] \Biggr\}{\text{,}} \end{split} $$ (4) 式中,
$ \sigma $ 代表Sigmod激活函数;$ \psi _1$ 、$ \psi _2$ 和$ \psi _3$ 表示由核大小$K$ 定义的标准二维卷积层。Triplet通过旋转操作构建各维度间的相关性,再对其进行残差变换,并以极小的计算成本对信息进行编码,在不需要太多可学习参数的前提下建立通道之间的相互依赖关系,有效地捕捉苹果图像中的关键特征。
1.3 Focal-CIoU损失函数
在机器学习任务中,常常面临数据集中不同类别的样本数量差异较大的问题,传统的交叉熵损失函数在处理样本不均衡问题时表现不佳,因为它将所有样本的重要性视为相等,继而导致模型在训练过程中更容易偏向于数量多的类别,公式如下:
$$ {\mathrm{CE}}(p,y) = {\mathrm{CE}}({P_t}) = - \ln ({P_t}) {\text{,}}$$ (5) 式中,CE是交叉熵(Cross entropy),
$p$ 代表模型预测样本属于类别1的概率(取值范围0~1),$y$ 表示样本的标签(取值为−1和1)。定义$$P_t=\left\{ \begin{array}{ll} {p}, &{{\text{当}} \; y=1{\text{时}}}\\ {1-p},& {{\text{其他}}} \end{array}\right. {\text{。}}$$ (6) 数据集中正常苹果和缺陷苹果图片数量均为1 600张左右,但标注数量略有差异,分别为4 624和2 039张。为了进一步加强模型对缺陷苹果的检测能力,引入Focal-CIoU损失函数。Focal Loss(FL)在交叉熵损失的基础上进行了改进,引入了平衡因子和聚焦因子,平衡因子用于调整正负样本之间的权重,聚焦因子用于调整难易样本间的损失权重;这样,既能调整正负样本的权重,又能控制难易分类样本的权重。FL表达式[28]为:
$$ {\mathrm{FL}}({P_t}) = - {\alpha _t}{(1 - {P_t})^\gamma }\ln ({P_t}), $$ (7) 式中,
${\alpha _t}$ 是超参数(取0.25);$ {(1 - {P_t})^\gamma } $ 是调节因子,$\gamma $ 是$ \geqslant $ 0的可调节聚焦参数(取2)。CIoU损失函数(LossCIoU)考虑边界框宽高比的尺度信息,在DIoU的基础上进行优化,计算公式[29]如下:
$$ {\mathrm{DI}}{\text{o}}{\mathrm{U}} = {\mathrm{IoU}} - \frac{{{d^2}}}{{{c^2}}}{\text{,}} $$ (8) $$ {\mathrm{Loss}}_{\mathrm{CIoU}} = {\mathrm{IoU}} - {\frac{{{\rho ^2}\left( {b,{b^{gt}}} \right)}}{{{c^2}}}^{}} - \alpha V{\text{,}} $$ (9) 式中,IoU为交并比(Intersection over Union),即“预测的边框”和“真实的边框”的交集和并集的比值;
$ d$ 为预测框与真实框中心点的长度,$ c$ 为最小外接矩形的对角线距离,$\rho $ 代表的是2个中心点之间的欧式距离,$ b、{b^{gt}} $ 分别代表了预测框和真实框的中心点,$\alpha $ 是权重函数,$V$ 用来衡量长宽比的相似度。Focal-CIoU损失函数(
${\mathrm{L}}{\text{os}}{{\text{s}}_{{\mathrm{Focal}} - {\mathrm{CIoU}}}} $ )为:$$ {\mathrm{L}}{\text{os}}{{\text{s}}_{{\mathrm{Focal}} - {\mathrm{CIoU}}}} = {\mathrm{Io}}{{\mathrm{U}}^\gamma } \times {\mathrm{Los}}{{\mathrm{s}}_{{\mathrm{CIoU}}}} {\text{。}}$$ (10) Focal-CIoU通过调整正负样本的权重和提升边界框回归精度,减少了训练过程中梯度的不稳定性,使模型更关注缺陷苹果,提高了检测性能。
2. 试验与结果分析
2.1 数据集
本研究所用数据集从百度网站和飞桨AI Studio网站上获取,这2个网站提供了丰富的图像资源,包含虫蛀、腐烂、机械损伤和褶皱共4类苹果缺陷图像(图4),数量分别为414、750、148和280张,以确保对于苹果目标的全面覆盖。数据集共
3213 张(正常苹果1621 张、缺陷苹果1592 张),训练集和验证集按照9∶1的比例划分,其中,训练集图片2 891张、验证集图片322张。为了增加数据的多样性,通过Python程序对部分数据集进行水平、垂直方向的翻转以及旋转45º操作。构建和处理数据集,确保模型在训练和验证过程中能够充分学习和适应苹果目标的多样性特征,为后续的目标检测提供数据保障。2.2 模型训练与评价指标
本试验平台基于Ubuntu 18.04.6的64位操作系统,显卡为GPU(NVIDIA GeForce RTX2080Ti),显存为12 G,使用PyTorch框架构建模型,编程语言为Python,Torch版本为1.10.1。在训练过程中,设置初始学习率为0.01、周期学习率为0.20、动量为0.937、权重衰减系数为0.000 5、批量大小为16。模型评估指标包括准确率(Precision,P)、召回率(Recall,R)、平均精确率均值(Mean average precision,mAP)和F1。TP (True positive)表示模型正确地检测出缺陷苹果的数量;TN (True negative)表示模型正确地排除正常苹果的数量;FP (False positive)表示模型错误地将正常苹果判定为缺陷苹果的数量; R指模型正确检测出的缺陷苹果(TP)占所有真正的缺陷苹果(TP + FN)的比例,表示模型对于真实缺陷苹果的检测能力;AP(Average precision)是衡量目标检测模型在不同召回率下准确性的指标,mAP是所有类别AP的平均,用来评估模型整体性能; F1综合了P和R这2个指标,能够更全面地评估模型的性能。
训练过程中损失值参数的动态变化如图5所示,随着训练轮次递增,损失值经历了急剧下降的阶段,最终趋向于0.025左右的平稳水平。训练损失和验证损失逐渐趋于拟合,意味着模型所学习的训练数据中的模式能够有效泛化到验证数据,表明该模型在面对新数据时具有优异的泛化性能。
2.3 Triplet的不同添加位置
Triplet插入YOLOv5s结构中的位置(A表示YOLOv5s-BiFPN-Triplet)主要有以下4种方式:1)在网络结构SPPF前面添加,即第9层[A+CIoU+Focal (9)];2)在网络结构最后一层添加,即第24层[A+ CIoU+Focal(24)];3)在网络结构SPPF前面和最后一层添加,即A+CIoU+Focal(9+25);4)替换Backbone中的C3模块,即A+CIoU+Focal。不同插入位置对比试验结果见表1。由表1可见,A+CIoU+Focal(24)比其他4种模型在各个方面均有不错的提升,准确率比YOLOv5s的提高了5.7个百分点,综合分析表明A+ CIoU+Focal(24)是最优的模型。
表 1 Triplet注意力机制不同插入位置的模型试验结果对比Table 1. Result comparison of Triplet attention mechanism with different insertion position% 模型1)Model P R mAP F1 YOLOv5s 75.4 83.6 86.5 79.29 A+ CIoU+Focal(9) 77.3 81 85.6 79.11 A+CIoU+Focal(9+25) 80.8 79.9 86.2 80.35 A+CIoU+Focal 75.9 85.2 87.1 80.28 A+CIoU+Focal(24) 81.1 85.8 90.0 83.38 1) A:YOLOv5s-BiFPN-Triplet,括号内数字表示Triplet插入位置。
1) The number in parentheses indicated the insertion position of Triplet.2.4 BiFPN添加不同注意力机制
以YOLOv5s-BiFPN为基础,逐步添加SE,CBAM、CA和Triplet注意力机制。SE仅考虑通道间的信息,忽略了位置信息;CBAM通过学习的方式自动获取每个特征通道和特征空间的重要程度;而CA不仅考虑通道间的关系,而且考虑方向相关的位置信息。试验结果见表2,由表2可见,BiFPN+CA在准确率和mAP方面优于BiFPN+SE和BiFPN+CBAM;Triplet注意力机制利用三分支结构能够捕获更多的关键信息,建立通道之间的相互依赖关系,试验结果也验证了BiFPN+Triplet是最优模型。
表 2 BiFPN添加不同注意力机制的模型试验结果对比Table 2. Result comparison of BiFPN with different attention mechanism% 模型Model P R mAP BiFPN+SE 76.1 83.1 87.0 BiFPN+CBAM 77.5 81.5 87.6 BiFPN+CA 78.6 81.3 88.1 BiFPN+Triplet 79.3 83.2 88.4 2.5 消融试验
为了验证各个模块的作用,以YOLOv5s为基础,进行逐步添加和替换,结果见表3。从表3中可以看出,与初始的YOLOv5s模型相比,单独添加BiFPN、Triplet或Focal-CIoU模块的模型准确率、召回率和mAP均有一定的提升。同时添加2种模块时,取得的效果更佳,添加BiFPN+Triplet的模型mPA比单独添加BiFPN的提高了0.4个百分点、准确率比单独添加Triplet的提高2.9个百分点;添加Triplet+Focal-CIoU的模型准确率和mAP比单独添加Triplet的分别提高1.1和0.3个百分点;添加BiFPN+Focal-CIoU的模型准确率和mAP比单独添加Focal-CIoU的分别提高1.7和1.0个百分点。当三者同时添加时,提高了模型对缺陷苹果的感知能力,与YOLOv5s模型相比,准确率、召回率和mAP分别提高5.7、2.2和3.5个百分点。
表 3 消融试验结果Table 3. The result of ablation test% BiFPN Triplet Focal-CIoU P R mAP 75.4 83.6 86.5 √ 80.3 84.3 88.0 √ 76.4 86.1 89.3 √ 78.4 83.0 89.8 √ √ 79.3 83.2 88.4 √ √ 77.5 84.5 89.6 √ √ 80.1 84.0 88.9 √ √ √ 81.1 85.8 90.0 为了验证Focal-CIoU损失函数的优越性,本文设计了YOLOv5s-BiFPN-Triplet+损失函数的消融试验,结果见表4。由表4可知,A+ CIoU+Focal与次优模型A+ SIoU相比,准确率提高了1个百分点、召回率和mAP提高0.2个百分点、F1提高0.62个百分点;虽然A+ WIoU的准确率与A+ CIoU+Focal相同,但召回率、mAP和F1均低于A+ CIoU+Focal。综合准确率、召回率、mAP和F1等指标, A+ CIoU+Focal是最优模型。
表 4 Focal-CIoU与其他损失函数对比Table 4. Comparison of Focal-CIoU with other loss functions% 模型1)Model P R mAP F1 A 79.3 83.2 88.4 81.20 A+ DIoU 79.1 84.8 89.2 81.85 A+ DIoU+Focal 79.4 84.3 89.6 81.78 A+ SIoU 80.1 85.6 89.8 82.76 A+ SIoU+Focal 77.9 85.3 88.5 81.43 A+ EIoU 78.7 84.3 88.7 81.40 A+ EIoU+Focal 80.7 80.0 88.1 80.35 A+ WIoU 81.1 79.4 88.2 80.24 A+CIoU+Focal 81.1 85.8 90.0 83.38 1) A:YOLOv5s-BiFPN-Triplet. 2.6 模型有效性验证
为了证明本文提出模型(BTF-YOLOv5s)的有效性,与当前主流的目标检测模型进行对比(表5)。由表5可见,与SSD相比,BTF-YOLOv5s的准确率提高了4.8个百分点、召回率和mAP分别提高了4.1和5.7个百分点,模型大小大幅度减小;与YOLOv3相比,BTF-YOLOv5s的准确率提高近10个百分点,模型小了近8倍;与YOLOv4相比,BTF-YOLOv5s虽然准确率低了3.1个百分点,但召回率和mAP分别提升近20和13个百分点,模型大小减少了17倍;与YOLOv5s、YOLOv7和YOLOv8s相比,BTF-YOLOv5s在准确率、召回率和mAP方面均有提升;YOLOv8n和YOLOv9的准确率相较于BTF-YOLOv5s各提高0.6和4.9个百分点,但二者召回率过低; F1显示BTF-YOLOv5s是最优的。综合分析,BTF-YOLOv5s显著优于大多数目标检测模型,更具有优势。图6直观地展示了SSD、YOLOv3、YOLOv5s等模型与BTF-YOLOv5s的mAP比较,在训练轮次逐步增加的过程中,BTF-YOLOv5s的mAP最终稳定在90%左右,明显优于其他模型。
表 5 BTF-YOLOv5与其他模型的对比Table 5. Comparison of BTF-YOLOv5 with other models模型
ModelP/% R/% mAP/% 模型大小/MB
Model sizeF1/% 参数
ParameterGFLOPs1) SSD 76.3 81.7 84.3 95.5 78.91 2.49×107 31.4 YOLOv3 71.8 83.8 86.5 123.5 77.34 3.30×107 78.1 YOLOv4 84.2 65.6 76.6 256.3 73.75 6.50×107 142.3 YOLOv5s 75.4 83.6 86.5 14.4 79.29 7.03×106 16.0 YOLOv7 79.6 83.1 87.1 142.1 81.31 3.72×107 105.1 YOLOv8n 81.7 76.6 87.4 6.2 79.07 3.01×106 8.2 YOLOv8s 73.6 83.0 87.2 22.5 78.02 1.11×107 28.6 YOLOv9 86.0 78.8 89.7 102.8 82.24 5.10×107 238.9 BTF-YOLOv5s 81.1 85.8 90.0 14.7 83.38 7.17×106 16.7 1) GFLOPs:每秒10亿次的浮点运算数。
1) GFLOPs: Giga floating-point operations per second.2.7 应用场景试验结果
本模型的应用场景之一即苹果采摘机器人在采摘过程中的分拣,故选择缺陷苹果所处的自然环境下的场景以及摆放不规整的场景。如图7所示,其中,第1行图像为原始图像、第2行图像为YOLOv5s检测图、第3行图像为BTF-YOLOv5s检测图;在图7的第1列中,BTF-YOLOv5s能够有效检测出正常苹果与缺陷苹果,准确率比YOLOv5s有显著提升;在第2列中对于像素更少的瑕疵斑点,YOLOv5s模型不仅出现了误检,而且准确率也低于BTF-YOLOv5s;在第3列中,BTF-YOLOv5s模型的检测效果明显优于YOLOv5s模型。
3. 结论
为了实现对瑕疵苹果的快速准确检测,本文提出基于BiFPN和Triplet注意力机制的YOLOv5s缺陷苹果识别算法。通过加权双向特征金字塔网络(BiFPN)来学习不同的输入特征;采用Triplet注意力机增强模型对目标之间的关联和上下文信息的表示能力;采用Focal-CIoU损失函数,在计算损失时调整损失权重,给予目标物体更多的关注。注意力机制的应用使模型更加关注目标,在4种位置的插入对比中,YOLOv5s网络结构最后一层的插入位置表现最为优越,与在网络结构SPPF前面和最后一层添加相比,准确率、召回率和mAP分别提高了0.3、5.9和3.8个百分点;BiFPN与SE、CBAM、CA、Triplet注意力机制两两组合,验证了BiFPN+Triplet的优越性;相较于BiFPN+CA,准确率、召回率和mAP分别提升了0.7、1.9和0.3个百分点。消融试验表明,以YOLOv5s为基础,同时添加3种模块的准确率、召回率和mAP分别提高了5.7、2.2和3.5个百分点,提升效果显著。以YOLOv5s-BiFPN-Triplet为基础,Focal-CIoU的准确率比次优损失函数SIoU高1个百分点;准确率、召回率和mAP比主流的YOLOv7算法提高了1.5、2.7和2.9个百分点,同时,模型大小从142.1 MB减小到14.7 MB,显著降低了内存占用,在计算资源受限的环境中,为部署目标检测系统提供了有力支持。YOLOv8n和YOLOv9的准确率虽略高于BTF-YOLOv5s,但均面临召回率过低的问题,而YOLOv8s的准确率只有73.6%。在应用场景的对比试验中,BTF-YOLOv5s也表现出较好的优越性。
-
![]()
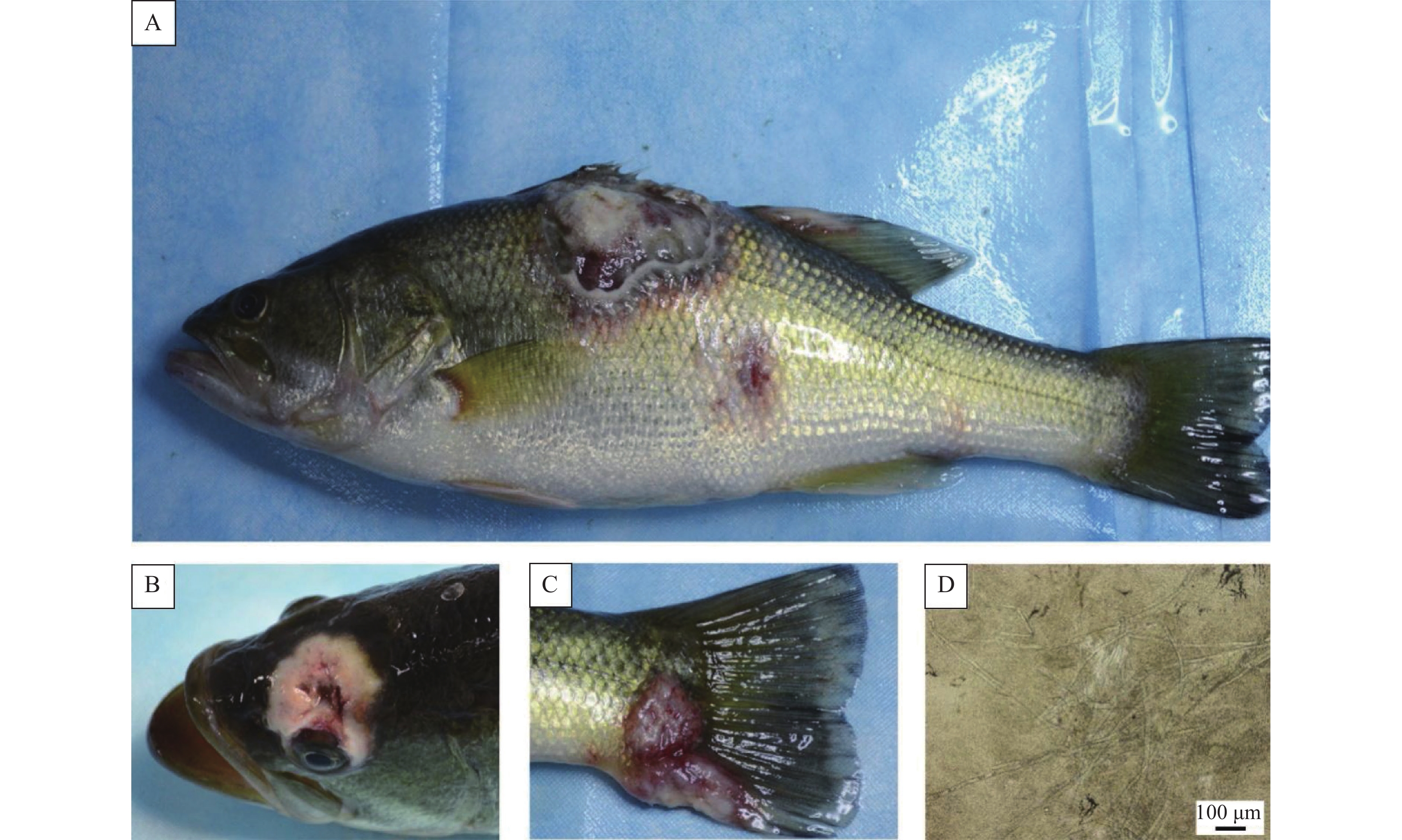
图 1 自然感染大口黑鲈临床症状及压片观察
A:体表溃疡与红斑;B:头部溃疡伴随颅骨破损;C:尾柄基部溃疡;D:溃疡压片组织中的菌丝体
Figure 1. Clinical symptoms of naturally infected Micropterus salmoides and squashing tissue observation
A: Ulceration and red spot on the body surface; B: Head ulcer with skull breakage; C: Ulceration at the base of the tail fin; D: Mycelia in the squashing tissue
![]()
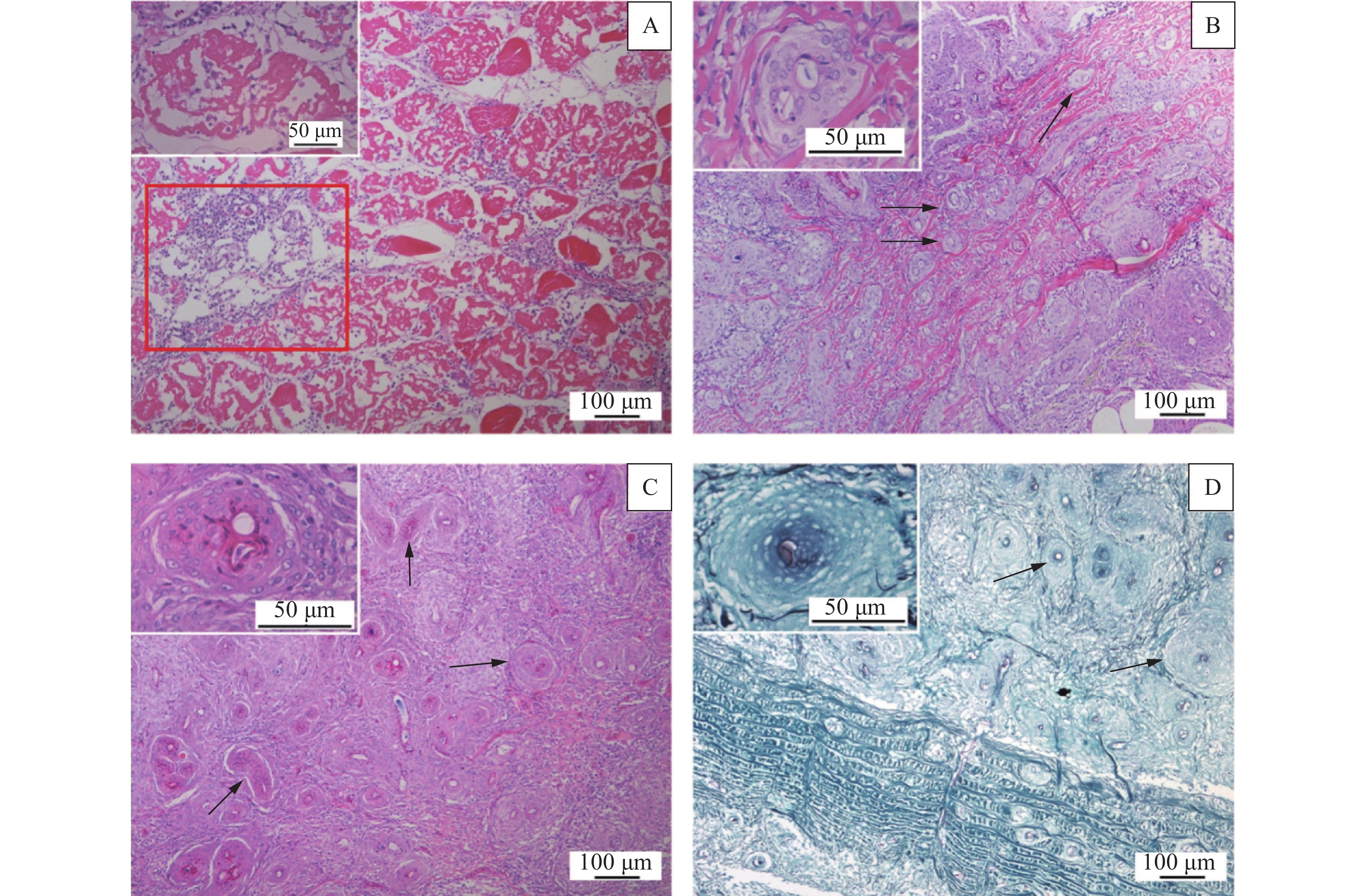
图 2 大口黑鲈组织病理学变化
A:肌肉坏死,红框表示肌纤维内大量炎症细胞浸润,左上角插入图为坏死肌纤维高倍数图,HE染色;B:皮肤真皮结缔组织中大量霉菌性肉芽肿(箭头标记),HE染色;C:肌肉组织中大量霉菌性肉芽肿(箭头标记),HE染色;D:真皮与肌肉中的霉菌性肉芽肿,中心为黑色菌丝体(箭头标记),PASM染色;图B、C、D左上角插入图为肉芽肿高倍数图
Figure 2. Histopathological lesions of Micropterus salmoides
A: Necrosis of muscle fibers, the red box indicates massive infiltration of inflammatory cells in the muscle fibres, insert at the upper left corner shows high magnification of necrotic muscle fibres, HE stained; B: Massive fungal granulomas in dermal connective tissue (marked by arrow), HE stained; C: Massive fungal granulomas in muscle tissue (marked by arrow), HE stained; D: Fungal granulomas in dermis and muscle, with black mycelia in the center (marked by arrow), PASM stained; Inserts at the upper left corners of figure B, C and D show enlarged granulomas
![]()
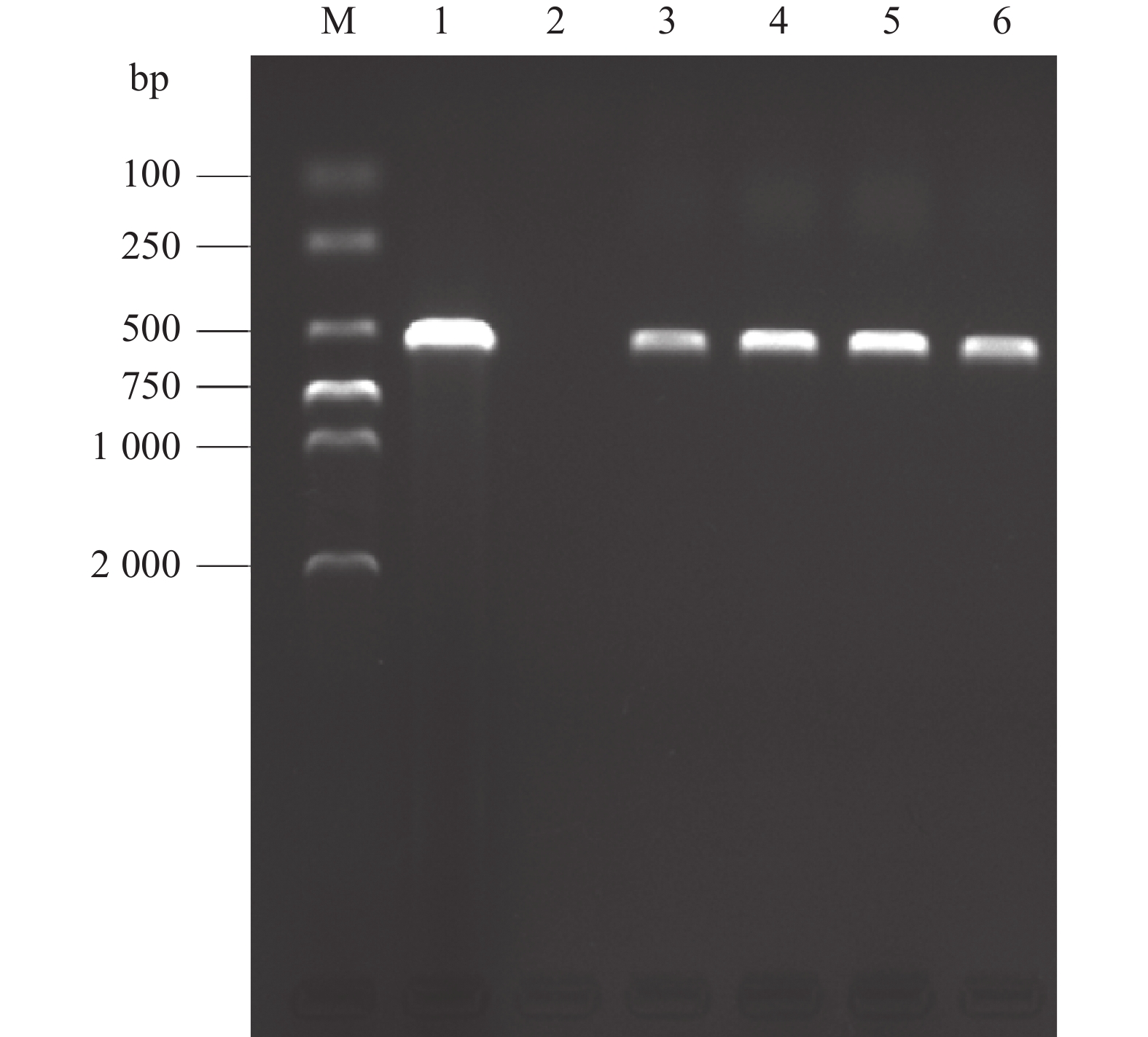
图 3 侵入丝囊霉菌ITS基因PCR扩增电泳图
M:2 000 bp marker;1:阳性对照;2:阴性对照;3,4:两养殖场患病鱼溃疡组织;5,6:分离霉菌BW1和BW3
Figure 3. Electropherogram of the PCR amplification product of Aphanomyces invadans ITS gene
M: 2 000 bp marker; 1: Positive control; 2: Negative control; 3 and 4: Ulcer organization of diseased fish in two farms; 5 and 6: Isolated fungi BW1 and BW3
![]()
图 4 侵入丝囊霉菌形态观察
A:琼脂块压片;B:纯培养物(箭头标记处为原孢子囊)
Figure 4. Morphology of Aphanomyces invadans
A: Squashing agar block; B: Pure culture (prosporangium is marked by arrow)
-
[1] SIBANDA S, PFUKENYI D M, BARSON M, et al. Emergence of infection with Aphanomyces invadans in fish in some main aquatic ecosystems in Zimbabwe: A threat to national fisheries production[J]. Transboundary and Emerging Diseases, 2018, 65(6): 1648-1656. doi: 10.1111/tbed.12922
[2] PARIA A, DEV A K, PRADHAN P K, et al. Evaluation of therapeutic potential of selected antifungal chemicals and drugs against Aphanomyces invadans[J]. Aquaculture, 2020, 529: 735643. doi: 10.1016/j.aquaculture.2020.735643
[3] SOSA E R, LANDSBERG J H, KIRYU Y, et al. Pathogenicity studies with the fungi Aphanomyces invadans, Achlya bisexualis, and Phialemonium dimorphosporum: Induction of skin ulcers in striped mullet[J]. Journal of Aquatic Animal Health, 2007, 19(1): 41-48. doi: 10.1577/H06-013.1
[4] IBERAHIM N A, TRUSCH F, VAN WEST P. Aphanomyces invadans, the causal agent of epizootic ulcerative syndrome, is a global threat to wild and farmed fish[J]. Fungal Biology Reviews, 2018, 32(3): 118-130. doi: 10.1016/j.fbr.2018.05.002
[5] KAR D, SINGHA R, DAS B K, et al. Preliminary epidemiological studies on epizootic ulcerative syndrome (EUS) in freshwater fishes of assam[J]. International Journal of Scientific Research in Science, Engineering and Technology, 2015, 1: 256-263.
[6] IBERAHIM N A, SOOD N, PRADHAN P K, et al. The chaperone Lhs1 contributes to the virulence of the fish-pathogenic oomycete Aphanomyces invadans[J]. Fungal Biology, 2020, 124(12): 1024-1031. doi: 10.1016/j.funbio.2020.09.003
[7] 常藕琴, 石存斌, 王亚军, 等. 鳢流行性溃疡综合征病原分离鉴定与病理形态学观察[J]. 中国水产科学, 2019, 26(6): 1213-1220. [8] 全国水产技术推广总站. 2019水产新品种推广指南[M]. 北京: 中国农业出版社, 2019: 3-15. [9] 夏焱春, 曹铮, 蔺凌云, 等. 大口黑鲈主要病害研究进展[J]. 中国动物检疫, 2018, 35(9): 72-76. doi: 10.3969/j.issn.1005-944X.2018.09.019 [10] GRIZZLE J M, ALTINOK I, FRASER W A, et al. First isolation of largemouth bass virus[J]. Diseases of Aquatic Organisms, 2002, 50(3): 233-235.
[11] OIE. Manual of diagnostic tests for aquatic animals[M]. Paris: Office International des Épizooties, 2010: 188-199.
[12] 陈绮梨, 常藕琴, 张德峰, 等. 加州鲈源弗氏柠檬酸杆菌的分离鉴定及耐药性分析[J]. 南方农业学报, 2021, 52(2): 465-474. doi: 10.3969/j.issn.2095-1191.2021.02.023 [13] 赵武义. 加州鲈源维氏气单胞菌的鉴定与分析[D]. 雅安: 四川农业大学, 2018. [14] ZHU Q, WANG Y, FENG J. Rapid diagnosis of largemouth bass ranavirus in fish samples using the loop-mediated isothermal amplification method[J]. Molecular and Cellular Probes, 2020, 52: 101569. doi: 10.1016/j.mcp.2020.101569
[15] SONGE M M, HANG OMBE M B, PHIRI H, et al. Field observations of fish species susceptible to epizootic ulcerative syndrome in the Zambezi River Basin in Sesheke District of Zambia[J]. Tropical Animal Health and Production, 2012, 44(1): 179-183.
[16] YADAV M K, PRADHAN P K, SOOD N, et al. Innate immune response of Indian major carp, Labeo rohita infected with oomycete pathogen Aphanomyces invadans[J]. Fish & Shellfish Immunology, 2014, 39(2): 524-531.
[17] DAS M K, DAS R K. A review of the fish disease epizootic ulcerative syndrome in India[J]. Environment and Ecology, 1993, 11(1): 134-145.
[18] PAGRUT N, GANGULY S, JAISWAL V, et al. An overview on epizootic ulcerative syndrome of fishes in India: A comprehensive report[J]. Journal of Entomology and Zoology Studies, 2017, 5: 1941-1943.
[19] LEI X, ZHAO R, GENG Y, et al. Nocardia seriolae: A serious threat to the largemouth bass Micropterus salmoides industry in Southwest China[J]. Diseases of Aquatic Organisms, 2020, 142: 13-21. doi: 10.3354/dao03517
[20] LILLEY J H, CALLINAN R B, CHINABUT S, et al. Epizootic ulcerative syndrome (EUS) technical handbook[J]. Bangkok: The Aquatic Animal Health Research Institute, 1998.
[21] VERMA D K, PERUZZA L, TRUSCH F, et al. Transcriptome analysis reveals immune pathways underlying resistance in the common carp Cyprinus carpio against the oomycete Aphanomyces invadans[J]. Genomics, 2021, 113(1): 944-956.
[22] KANBUTRA P, HANJAVANIT C, HORIUCHI E, et al. The in vitro antifungal effects of chlorine dioxide on water molds[J]. Aquaculture Science, 2010, 58(2): 219-224.
[23] CHOONGO K, HANG’OMBE B, SAMUI K L, et al. Environmental and climatic factors associated with epizootic ulcerative syndrome (EUS) in fish from the Zambezi floodplains, Zambia[J]. Bulletin of Environmental Contamination and Toxicology, 2009, 83(4): 474-478. doi: 10.1007/s00128-009-9799-0
[24] BORISUTPETH M, KANBUTRA P, WEERAKHUN S, et al. In vitro antifungal activity of Cassia fistula L. against selected pathogenic water molds[J]. International Journal of Phytomedicine, 2014, 6(2): 237.
[25] SHANTHI MARI L S, JAGRUTHI C, ANBAZAHAN S M, et al. Protective effect of chitin and chitosan enriched diets on immunity and disease resistance in Cirrhina mrigala against Aphanomyces invadans[J]. Fish & Shellfish Immunology, 2014, 39(2): 378-385.

 下载:
下载:





















































计量
- 文章访问数: 114
- HTML全文浏览量: 5
- PDF下载量: 12
