Effects of duck Tembutsu virus E protein and its domains of I, II, III on DEF cell cycle and apoptosis
-
摘要:目的
探究鸭坦布苏病毒(Duck Tambusu virus,DTMUV) E蛋白全长及其结构域I、II和III (DI、DII和DIII)对鸭胚成纤维细胞(Duck embryo fibroblast,DEF)的细胞周期与凋亡的影响。
方法本研究设计、合成DTMUV E蛋白全长及其DI、DII和DIII真核表达质粒,并转染至DEF,用流式细胞仪检测不同蛋白引起的DEF细胞凋亡和细胞周期的变化。
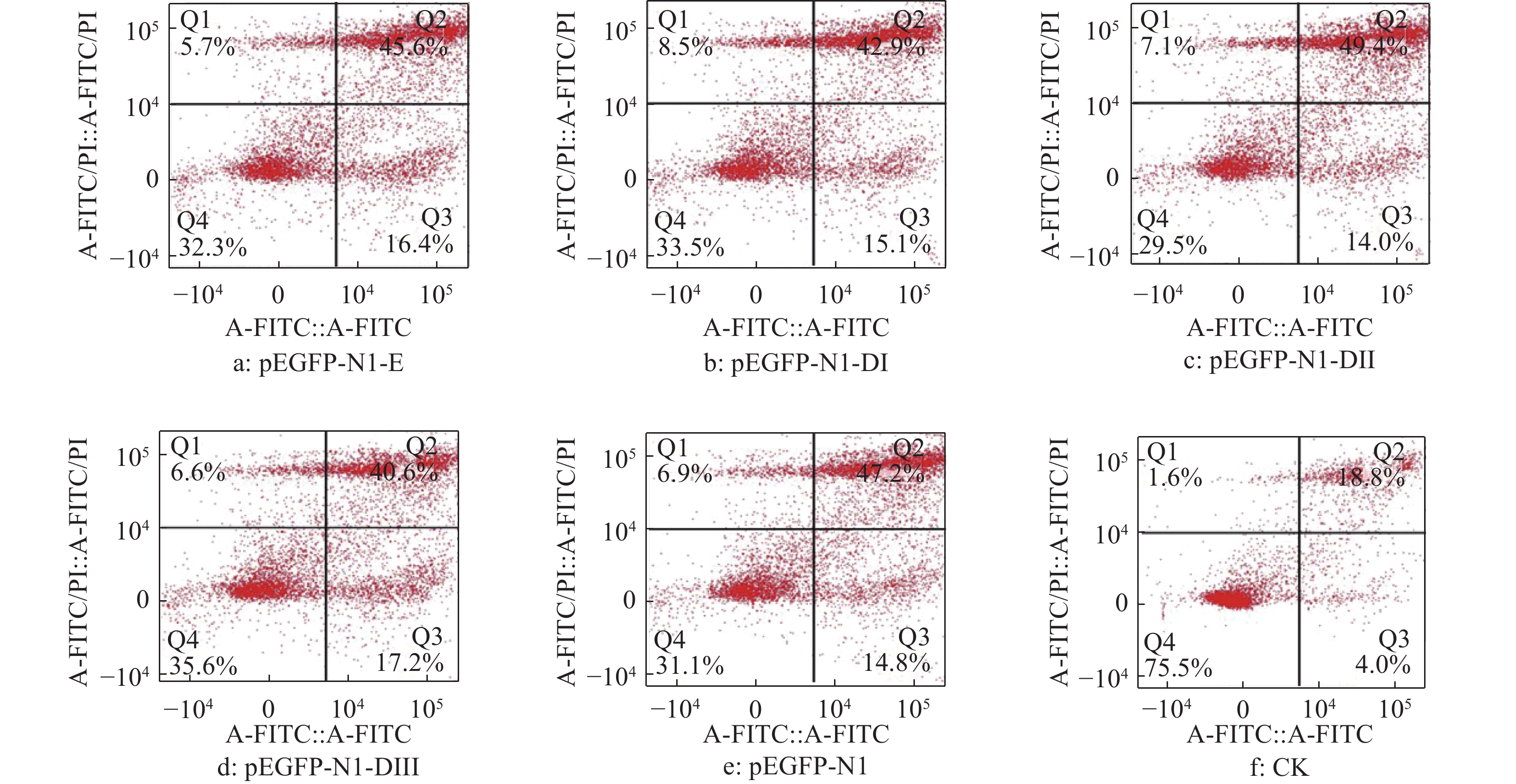
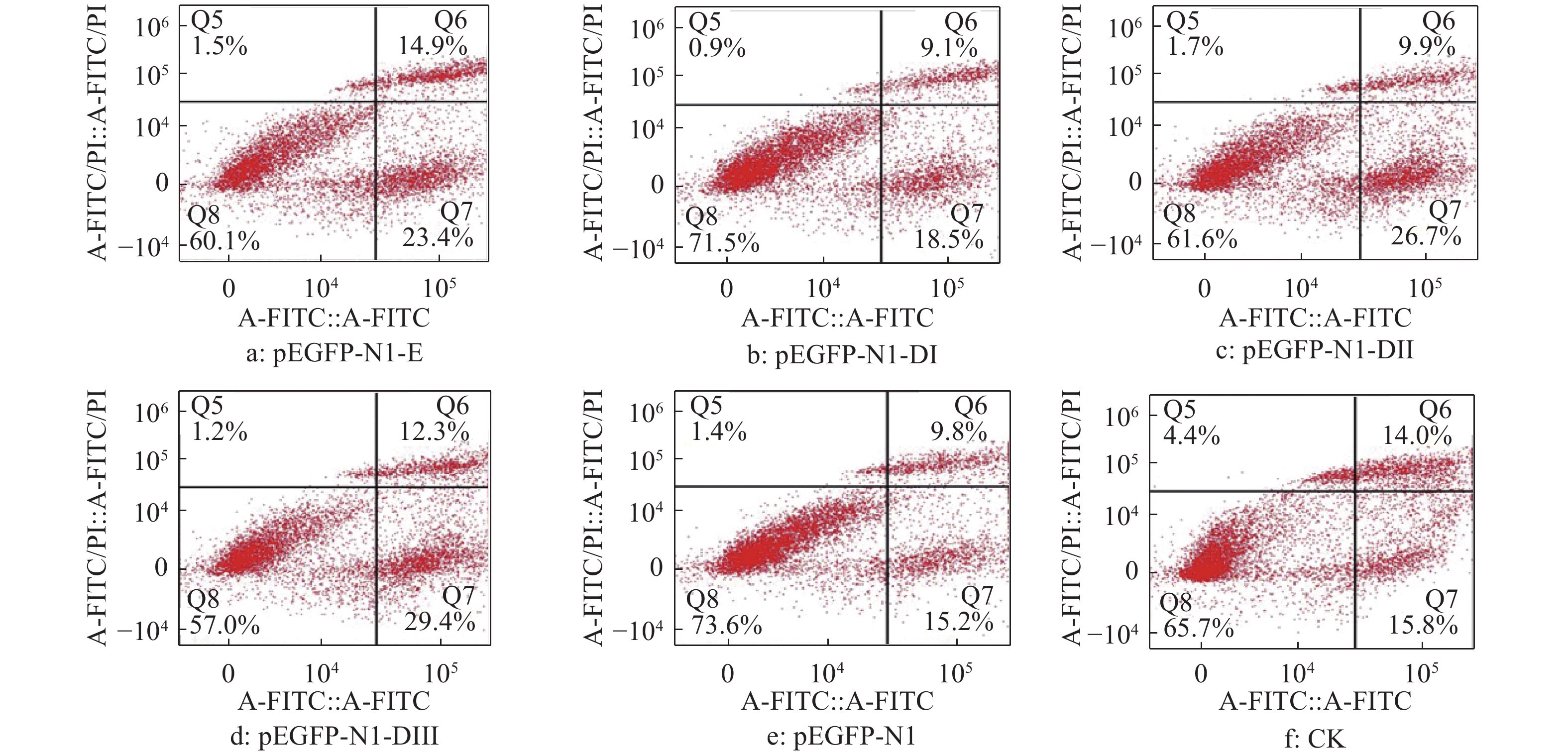
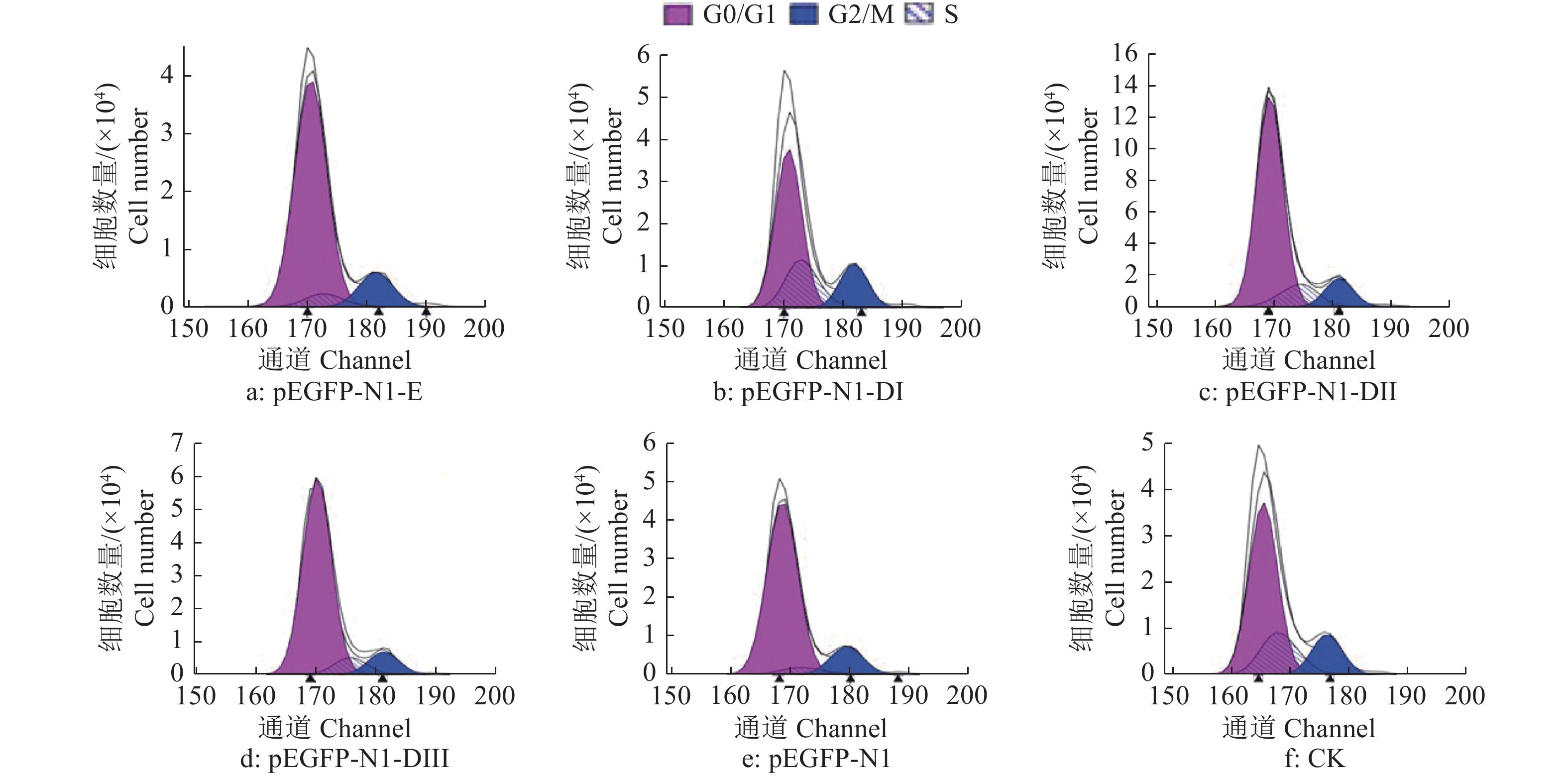
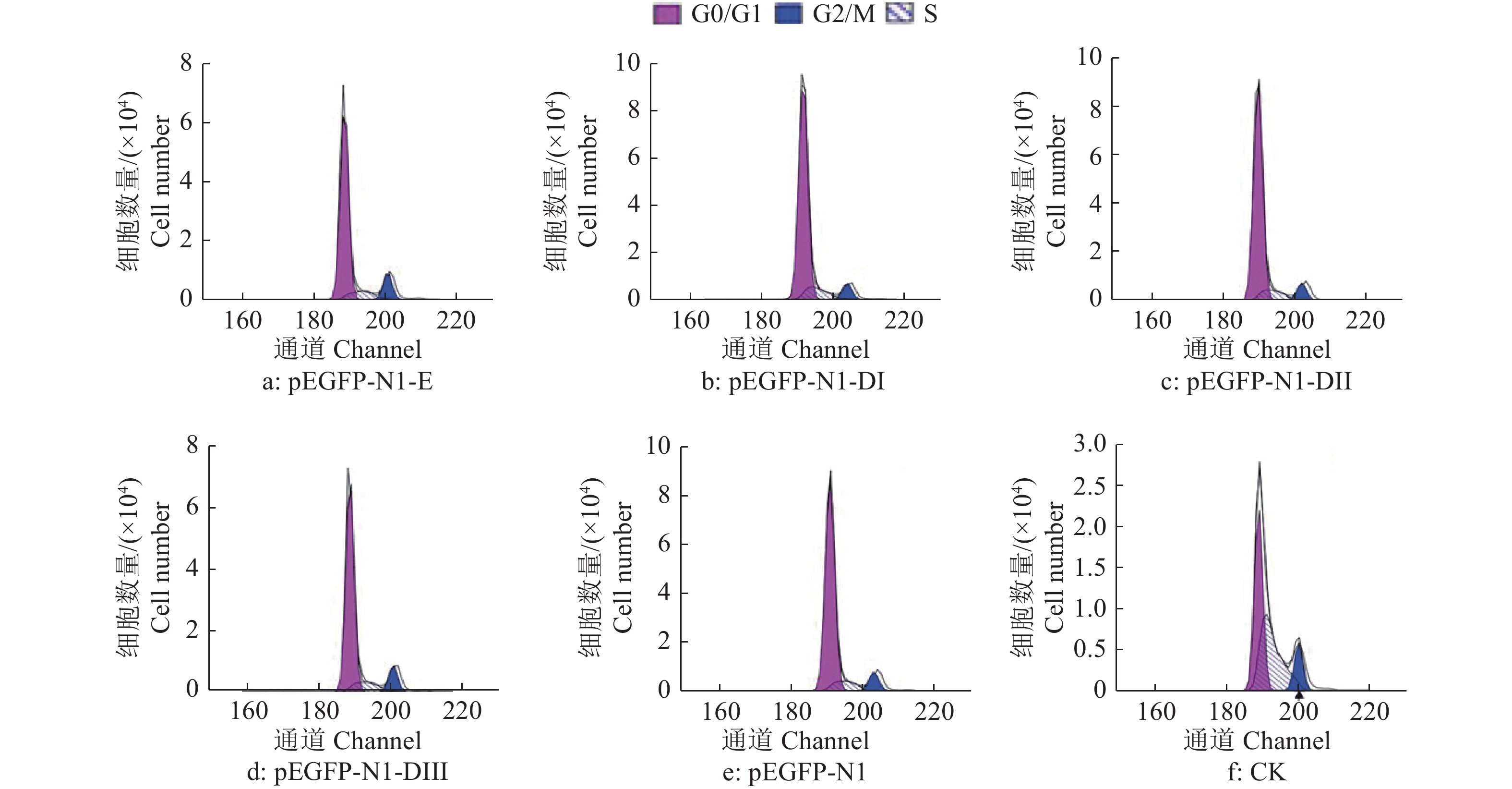
结果细胞凋亡结果显示:质粒转染细胞24 h后,E蛋白全长及DI、DII、DIII诱导的DEF早期凋亡率分别是16.4%、15.1%、14.0%和17.2%;质粒转染细胞36 h后,E蛋白全长及DI、DII、DIII诱导的DEF早期凋亡率分别是23.4%、18.5%、26.7%和29.4%。细胞周期检测结果显示:E蛋白全长及DI、DII、DIII的质粒转染细胞24、36 h 后,DNA合成期(S期)细胞比例都明显高于pEGFP-N1空载体转染组。质粒转染24 h后,E蛋白及DI、DII、DIII的S期细胞比例分别为5.43%、22.58%、12.75%和12.80%;质粒转染细胞36 h后,E蛋白及DI、DII、DIII的S期细胞比例分别为9.98%、11.44%、10.44%和11.00%。
结论DTMUV E蛋白全长及其3个结构域均能诱导早期细胞凋亡,引起细胞S期的停滞,但是各蛋白片段诱导细胞凋亡和引起细胞周期变化的能力存在一定差异。
Abstract:ObjectiveTo investigate the effects of the whole length of Duck Tambusu virus (DTMUV) E protein and its domains of I, II and III (DI, DII and DIII) on the cell cycle and apoptosis of duck embryo fibroblast (DEF) cells.
MethodThe full-length of DTMUV E protein, DI, DII and DIII eukaryotic expression plasmids were designed and synthesized, and transfected into DEF. The changes of DEF apoptosis and cell cycle caused by different proteins were detected by flow cytometry.
ResultThe results of apoptosis showed that after 24 h of plasmid transfection, the apoptosis rates of DEF induced by the full length, DI, DII and DIII were 16.4%, 15.1%, 14.0% and 17.2% respectively; after 36 h of plasmid transfection, the apoptosis rates of DEF induced by E protein full length, DI, DII and DIII were 23.4%, 18.5%, 26.7% and 29.4%. The results of cell cycle detection showed that the percentage of DNA synthesis phase (S-phase) cells transfected with E protein full length, DI, DII and DIII plasmid was obviously higher than those of cells transfected with pEGFP-N1 empty vector 24 h and 36 h later. After 24 h of plasmid transfection, the percentage of S-phase cells in E protein, DI, DII and DIII groups were 5.43%, 22.58%, 12.75% and 12.80%, respectively; After 36 h of plasmid transfection, the percentage of S-phase cells in E protein, DI, DII and DIII groups were 9.98%, 11.44%, 10.44% and 11.00%, respectively.
ConclusionThe full length of DTMUV E protein and its three domains can induce early cell apoptosis and arrest the S-phase cells, but the ability of each protein fragment to induce cell apoptosis and change the cell cycle is different.
-
Keywords:
- Duck Tembusu virus /
- E protein /
- Domain /
- Cell cycle /
- Apoptosis
-
广泛分布在自然界的虫生真菌是害虫生物防治的重要资源[1]。据不完全统计,全球已记载的虫生真菌约100属1000多种,一些有代表性的种类已被开发成环境友好型真菌杀虫剂[2],如球孢白僵菌Beauveria bassiana、金龟子绿僵菌 Metarhizium anisopliae、玫烟色棒束孢Isaria fumosorosea(原名玫色拟青霉Paecilomyces fumosoroseus)、蜡蚧轮枝菌Lecanicillium lecanii等[3]。作为一种重要的昆虫病原真菌属,现已发现的棒束孢属真菌有100多种,在生物防治和生产生活中发挥着重要作用。蝉棒束孢I. cicadae和细脚棒束孢I. tenuipes是一种药食两用真菌,在东亚地区被用于制作传统中草药和保健品[4],具有降血糖、抗肿瘤、抗菌、抗抑郁、抗氧化、抗衰老、降血脂、免疫调节等作用[5]。粉棒束孢分布范围广、寄主多,已被应用于防治温带农林害虫和线虫等[6]。玫烟色棒束孢已应用于对粉虱、蚜虫等刺吸式口器害虫的生物防治,并有多种商品试剂开发问世[7]。环链棒束孢I. cateinannulata对螨虫和鳞翅目害虫有较好的防效,在森林害虫的调控中起着重要作用[8]。爪哇棒束孢I. javanica制成的颗粒剂能有效控制稻飞虱的种群数量。我国地理环境复杂多样,气候复杂多变,为各类虫生真菌的形成提供了有利条件。白僵菌属与绿僵菌属的相关研究较多,也更成熟,而棒束孢属的报道相对较少。
黄曲条跳甲Phyllotreta striolata是十字花科蔬菜的主要害虫,幼虫和成虫均能为害植株,幼虫生活在地下,啃食植物根系,导致植株萎蔫,成虫为害植物叶片和嫩稍,造成芽株干枯甚至死亡,发生严重时可对苗期蔬菜造成毁灭性的危害[9-10]。本研究利用天然农药与化学生物学教育部重点实验室于2013至2019年采集到的国内部分地区土壤,对这些土壤中的棒束孢属菌进行分离、鉴定,并从中挑选9种棒束孢属真菌菌株对黄曲条跳甲进行生物活性测定,以期了解国内地区棒束孢属菌的多样性及分布规律,为虫生真菌资源的保护和农林害虫的生防产品合理开发提供新的菌种资源。
1. 材料与方法
1.1 土壤的采集
采集福建、广东、广西、云南、贵州、湖南、湖北、河南、河北等地区的土壤。记录采集地点、生境、经纬度等信息,每个采集地点都分3处位置采集地表下15~20 cm的土样各100 g,土样用自封袋封装,于4 ℃条件下保存。
1.2 菌株的分离纯化
将采集回来的土壤样品用孔径0.45 mm的筛子筛出细土,每份土壤样品准确称量3份10.0 g细土,用100 mL 0.1%(φ)的吐温−80溶液制成土壤悬浮液,漩涡震荡摇匀,静置15 min。在超净工作台中,将制备好的选择培养基(马铃薯200 g/L、葡萄糖20 g/L、琼脂20 g/L、放线菌酮4 mL/L、氯霉素4 mL/L、孟加拉红0.013 g/L)倒平板,待培养基静置凝固后用移液枪吸取100 μL土壤悬浮液至选择培养基,用玻璃刮铲涂抹均匀,晾干。在(28±1)℃恒温培养箱中培养,待选择培养基上长出单菌落后,用接种环挑取,接种至PDA(马铃薯200 g/L、葡萄糖20 g/L、琼脂20 g/L)培养基上进行纯培养,不断重复,直至菌株被纯化。成功纯化后,将菌株接种到试管斜面培养基上,待菌落长满斜面后于4 ℃冰箱中保存。
1.3 菌种鉴定
1.3.1 形态学鉴定
将纯化后的菌株接种至PDA培养基上进行活化,(28±1) ℃条件下培养3~5 d,待菌株开始产孢时,挑取菌丝到载玻片上,盖上盖玻片,置于光学显微镜下观察其分生孢子形态、产孢结构,并拍照记录。继续培养到12 d,观察记录菌落的大小、表面形态特征,并拍照记录。
1.3.2 分子生物学鉴定
菌种种属鉴定利用rDNA-ITS基因序列进行真菌ITS1-5.8S-ITS4 rDNA同源性分析,PCR通用引物为:ITS1(5′-TCCGTAGGTGAACCTGCGG-3′)和ITS4(5′-TCCTCCGCTTATTGATATGC-3′)。反应扩增条件为:94 ℃预变性3 min;94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸1 min,共33个循环;72 ℃延伸10 min。PCR产物送公司测序。收集真菌rDNA-ITS基因序列(长度为550~580 bp),将扩增所得到的ITS序列进行BLAST相似性比对分析,记录序列相似性最高的真菌种属,用MEGA-X软件构建ML系统发育树,最终确定真菌种属。
1.4 对黄曲条跳甲的活性测定
供试黄曲条跳甲采集自华南农业大学试验农场,置于养虫笼内用新鲜菜心饲养,保持适宜的光照、湿度和温度,3~5 d后选择生长期一致的成虫供生物活性测定。
将分离纯化后的菌株接种到PDA培养基上进行活化,(28±1) ℃条件下培养7~14 d,待菌株充分产孢后,刮取菌落表面分生孢子,用0.05%(φ)吐温−80溶液配制成1.0×108 mL−1孢子悬浮液备用。
参考标准[11]测定孢子悬浮液对黄曲条跳甲的生物学活性。将大小基本一致、健康的黄曲条跳甲成虫用CO2麻醉后转移到1.5 mL离心管,加入1 mL制备好的孢子悬浮液,迅速盖好并上下颠倒30次后,将黄曲条跳甲取出转移到铺有滤纸的一次性塑料碗中,置于(25±1) ℃、光周期为14 h光∶10 h暗,相对湿度(95±5)%的人工气候箱中饲养。浸泡后的黄曲条跳甲用白萝卜块饲养,每2 d更换1次,每天记录1次试虫的死亡数,并将死虫转移到铺有滤纸的培养皿上(己灭菌,加适量无菌水保湿),置于相同条件人工气候箱中培养,根据虫体体表长出的特征菌丝确定是否为真菌侵染致死。以高温灭菌后的0.05% (φ)吐温−80溶液处理为对照,每个处理包括10头黄曲条跳甲成虫,每个处理重复3次。
1.5 数据分析
应用Excel 2010计算生物多样性指数,包括多样性指数(Shannon-Wiener指数,H)、均匀度指数 (Pielou指数,J)和优势度指数 (Berger-Parker指数,d)。计算公式如下:
$$ \begin{array}{c} H = - \displaystyle\sum\limits_{i = 1}^S {{{{P}}_i}} {\ln {{{P}}_i}} ,\\ J = H/{H_{\max }},\\ {{{H}}_{\max }} = \ln S,\\ d = {n_{\max }}/N, \end{array} $$ 式中,S为物种数,ni为物种i的个体数,Pi为物种i的个体数占群落总个体数的比例 (即物种i的多度),Hmax为H的最大理论值,N为群落物种个体总数,nmax为群落中最大物种的个体数。
致病力用校正死亡率来进行判断,计算公式如下所示:
死亡率=死虫数/试虫数×100%,
校正死亡率=(处理死亡率−对照死亡率)/(1−对照死亡率)×100%。
2. 结果与分析
2.1 土壤中棒束孢属的多样性
虫生真菌的分离鉴定结果表明,这些菌株分属于棒束孢属的4个种,分别是玫烟色棒束孢Isaria fumosorosea、粉棒束孢I. farinosa、爪哇棒束孢I. javanica和环链棒束孢I. cateniannulate。菌株的产孢结构及孢子形态如图1所示。
![图 1 4种棒束孢菌株的产孢结构及孢子形态]() 图 1 4种棒束孢菌株的产孢结构及孢子形态a:玫烟色棒束孢;b:环链棒束孢;c:爪哇棒束孢;d:粉棒束孢Figure 1. Sporulation structure and spore morphology of four Isaria strainsa: Isaria fumosorosea; b: Isaria cateniannulate; c: Isaria javanica; d: Isaria farinosa
图 1 4种棒束孢菌株的产孢结构及孢子形态a:玫烟色棒束孢;b:环链棒束孢;c:爪哇棒束孢;d:粉棒束孢Figure 1. Sporulation structure and spore morphology of four Isaria strainsa: Isaria fumosorosea; b: Isaria cateniannulate; c: Isaria javanica; d: Isaria farinosa玫烟色棒束孢菌株的菌落正面呈现杏色粉质状,背面呈现淡杏色,环链棒束孢菌株的菌落形态与玫烟色棒束孢相似,正面呈现淡杏色,比玫烟色棒束孢菌株颜色浅,而粉棒束孢菌株的菌落背面呈现白色,爪哇棒束孢菌株的菌落正面和背面均呈现白色,棒束孢属的孢子形状均为椭圆形。
对41株棒束孢菌株构建的系统发育树如图2所示,结果显示分离纯化得到的菌株中含爪哇棒束孢34株,玫烟色棒束孢5株,环链棒束孢和粉棒束孢各1株。
![图 2 基于棒束孢菌株ITS序列构建的系统发育树]() 图 2 基于棒束孢菌株ITS序列构建的系统发育树Figure 2. Phylogenetic tree based on ITS sequence of Isaria strains
图 2 基于棒束孢菌株ITS序列构建的系统发育树Figure 2. Phylogenetic tree based on ITS sequence of Isaria strains2.2 棒束孢菌多样性指数分析
从土壤中共分离得到棒束孢属菌41株,其中爪哇棒束孢数量最多(34株),玫烟色棒束孢次之(5株),粉棒束孢和环链棒束孢最少,均为1株。结果表明爪哇棒束孢为所分离棒束孢菌中的优势菌株,占总数量的82.93%,玫烟色棒束孢次之,占总数量的12.2%,粉棒束孢和环链棒束孢各占2.44%。多样性指数(H)为0.593,均匀度指数(J)为0.160。
5种不同生境条件下,优势种均为爪哇棒束孢,休耕地、耕地、果园、草地和林地的优势度指数分别为0.889、1.000、1.000、0.889和0.545。休耕地、耕地、果园、草地和林地的多样性指数(H)分别为0.349、0、0、0.349和1.121,林地的H最大,说明林地的物种丰富度最高。休耕地、耕地、果园、草地和林地的均匀度指数分别为0.159、0、0、0.159和0.468,林地的均匀度指数最大,说明林地的棒束孢菌种间分配最均匀。
2.3 棒束孢菌的生境分布特征
对棒束孢属菌的生长环境进行分析,结果表明,在东经101.14°~119.65°,北纬20.42°~31.030°分离得到这些棒束孢属菌株,福建、广东、广西地区分离出来的棒束孢属菌株数量高于其他省份地区(表1),而在这些地区中,可以看出草地、休耕地和林地等环境更有利于棒束孢菌的生长(图3)。
表 1 棒束孢菌株的具体信息Table 1. Specific information of Isaria strains菌株编号
Strain No.菌种
Species采集地点
Location植被环境
Habitat经、纬度/(°)
Latitude and longitudeIjFJ01 爪哇棒束孢 Isaria javanica 福建南平市 耕地 Farmland E118.18,N26.64 IjFJ02 爪哇棒束孢 I. javanica 福建龙岩市连城县 休耕地 Fallow land E116.78,N25.71 IjFJ04 爪哇棒束孢 I. javanica 福建宁德市福安市 休耕地 Fallow land E119.65,N27.07 IjFJ05 爪哇棒束孢 I. javanica 福建三明市沙县 耕地 Farmland E117.97,N26.50 IjFJ06 爪哇棒束孢 I. javanica 福建南平市武夷山市 耕地 Farmland E117.99,N27.61 IjFJ17 爪哇棒束孢 I. javanica 福建三明市永安市 休耕地 Fallow land E117.32,N25.96 IjGD02 爪哇棒束孢 I. javanica 广东潮州市饶平县 耕地 Farmland E116.80,N24.13 IjGD07 爪哇棒束孢 I. javanica 广东湛江市徐闻县 耕地 Farmland E110.25,N20.42 IjGD12 爪哇棒束孢 I. javanica 广东梅州市梅县 休耕地 Fallow land E116.40,N24.65 IjGD14 爪哇棒束孢 I. javanica 广东江门市恩平县 休耕地 Fallow land E112.39,N22.30 IjGD15 爪哇棒束孢 I. javanica 广东惠州市香炉峰 休耕地 Fallow land E114.03, N23.60 IjGD17 爪哇棒束孢 I. javanica 广东惠州市博罗县 耕地 Farmland E114.28,N23.35 IjGD18 爪哇棒束孢 I. javanica 广东惠州市龙门县 休耕地 Fallow land E113.98,N23.53 IjGD20 爪哇棒束孢 I. javanica 广东惠州市龙门县 果园 Orchard E114.13,N23.66 IjGD21 爪哇棒束孢 I. javanica 广东中山市三乡镇 耕地 Farmland E113.44,N22.36 IjYN7003 爪哇棒束孢 I. javanica 云南普洱市宁洱县 耕地 Farmland E101.14,N23.19 IfGZ3703 玫烟色棒束孢 I. fumosorosea 贵州黔南州福泉市 草地 Grassland E107.53,N26.7 IfGZ4206 玫烟色棒束孢 I. fumosorosea 贵州毕节市七星关区 休耕地 Fallow land E105.31,N27.25 IfGZ4304 玫烟色棒束孢 I. fumosorosea 贵州毕节市大方县 林地 Woodland E105.82,N27.27 IjGZ4601 爪哇棒束孢 I. javanica 贵州遵义市播州区 草地 Grassland E106.66,N27.59 IjGX20A02 爪哇棒束孢 I. javanica 广西南宁市武鸣区 林地 Woodland E108.27,N23.01 IjGX20J02 爪哇棒束孢 I. javanica 广西南宁市武鸣区 果园 Orchard E108.27,N23.01 IjGX21G01 爪哇棒束孢 I. javanica 广西南宁市青秀区 果园 Orchard E108.77,N22.83 IjGX21L01 爪哇棒束孢 I. javanica 广西南宁市青秀区 果园 Orchard E108.77,N22.83 IjGX2303 爪哇棒束孢 I. javanica 广西贵港市港南区 草地 Grassland E109.72,N23 IjGX26S01 爪哇棒束孢 I. javanica 广西柳州市柳江区 草地 Grassland E109.28,N24.25 IjGX2701 爪哇棒束孢 I. javanica 广西柳州市柳城县 林地 Woodland E109.28,N24.65 IjGX2802 爪哇棒束孢 I. javanica 广西柳州市融水苗族自治县 草地 Grassland E109.28,N25.06 IjGX2906 爪哇棒束孢 I. javanica 广西柳州市柳城县 草地 Grassland E108.95,N24.51 IjGX30A01 爪哇棒束孢 I. javanica 广西河池市宜州区 林地 Woodland E108.31,N24.68 IjGX30S01 爪哇棒束孢 I. javanica 广西河池市宜州区 林地 Woodland E108.31,N24.68 IjGX32A03 爪哇棒束孢 I. javanica 广西河池市金城江区 林地 Woodland E107.77,N24.84 IcGX32S01 环链棒束孢 I. cateniannulate 广西河池市金城江区 林地 Woodland E107.77,N24.84 IjGX33H05 爪哇棒束孢 I. javanica 广西河池市南丹县 休耕地 Fallow land E107.56,N25.04 IfGX6701 粉棒束孢 I. farinosa 广西桂林市兴安县 林地 Woodland E110.77,N25.71 IfGX70A01 玫烟色棒束孢 I. fumosorosea 广西柳州市鹿寨县 林地 Woodland E109.84,N24.51 IjGX7704 爪哇棒束孢 I. javanica 广西贺州市平桂区 草地 Grassland E111.58,N24.37 IjH0904 爪哇棒束孢 I. javanica 湖北省孝感市孝南区 草地 Grassland E113.938,N31.030 IfH6102 玫烟色棒束孢 I. fumosorosea 湖南省怀化市塘湾镇 林地 Woodland E110.420,N27.216 IjH2702 爪哇棒束孢 I. javanica 湖北省孝感市应城市 草地 Grassland E113.576,N30.868 2.4 棒束孢属真菌对黄曲条跳甲成虫的致病力
从土壤样品分离出来的所有棒束孢属真菌菌株中,随机选取了9株不同的菌株对黄曲条跳甲成虫进行生物活性测定。结果如表2所示,其中玫烟色棒束孢IfH6102菌株在处理黄曲条跳甲成虫11 d后,校正死亡率最高,达60.87%。
表 2 棒束孢菌株对黄曲条跳甲的致病力Table 2. Pathogenicity of Isaria strain to Phyllotreta striolata菌株名称
Strain菌株编号
Strain No.校正死亡率1)/% Corrected mortality 7 d 9 d 11 d 爪哇棒束孢 Isaria javanica IjGX20A02 6.67±0.09c 6.67±0.09b 6.67±0.09c 爪哇棒束孢 I. javanica IjGX21G01 0d 0c 0d 环链棒束孢 I. cateniannulate IcGX32S01 6.67±0.09c 7.14±0.09b 21.43±0.09bc 玫烟色棒束孢 I. fumosorosea IfGX21L01 13.33±0.09bc 7.14±0.09b 7.14±0.09c 爪哇棒束孢 I. javanica IjGX33H05 0d 7.14±0.09b 21.43±0.25bc 玫烟色棒束孢 I. fumosorosea IfGX70A01 6.67±0.09c 13.33±0.09b 13.33±0.09c 爪哇棒束孢 I. javanica IjGX7704 6.67±0.05c 6.67±0.05b 16.67±0.05c 玫烟色棒束孢 I. fumosorosea IfGZ4304 6.67±0.09c 13.33±0.09b 13.33±0.09c 玫烟色棒束孢 I. fumosorosea IfH6102 26.92±9.43a 44.00±9.43a 60.87±8.16a 1)表中数据为平均值±标准误;同列数据后不同小写字母表示差异显著(P<0.05,Duncan’s法)
1) Data in the table were mean value ± standard error; Different lowercase letters in the same column indicated significant differences (P<0.05,Duncan’s test)玫烟色棒束孢IfH6102菌株处理黄曲条跳甲出现死虫后,将死虫转移到铺有滤纸的培养皿上,在光学显微镜下,可观察到白色菌丝首先从黄曲条跳甲尸体的触角、足关节、胸部、胸足等刚毛浓密部位和节间膜上伸出体外,随着时间推移,整个虫体被白色菌丝包裹,如图4所示。
![图 4 菌株IfH6102侵染黄曲条跳甲不同时间的形态]() 图 4 菌株IfH6102侵染黄曲条跳甲不同时间的形态a:死亡后3 d(背部);b:死亡后7 d(背部);c:死亡后11 d(腹部)Figure 4. Characteristics of Phyllotreta striolata after infected by strain IfH6102 for different timea: Three days after death (back); b: Seven days after death (back); c:11 days after death (abdomen)
图 4 菌株IfH6102侵染黄曲条跳甲不同时间的形态a:死亡后3 d(背部);b:死亡后7 d(背部);c:死亡后11 d(腹部)Figure 4. Characteristics of Phyllotreta striolata after infected by strain IfH6102 for different timea: Three days after death (back); b: Seven days after death (back); c:11 days after death (abdomen)3. 讨论与结论
随着社会经济的快速发展,人类活动对自然的影响越来越大,加之气候不断变化,许多物种生境发生了较大变化,环境保护、绿色健康发展变得越来越重要,生物防治也受到广泛关注。作为一种重要的生防资源,昆虫病原真菌也是生态系统的重要组成部分,了解和保护昆虫病原真菌资源有利于人与自然的和谐相处。爪哇棒束孢能有效防控褐飞虱Nilaparvata lugens、美国白蛾Hyphantria cunea、蓟马等农业害虫的种群数量[12]。玫烟色棒束孢对粉虱(烟粉虱Bemisia tabaci、温室白粉虱Trialeurodes vaporariorum等)、蚜虫(梨黄粉蚜Aphanostigma iaksuiense、桃粉蚜Hyalopterus arundimis等)、象甲(茶芽粗腿象甲 Anthonarainae quari maculate、茶大灰象甲Sympiezomias citri等)、柑橘木虱Diaphorina citri等多种害虫均有较高的控制潜力[13-17]。粉棒束孢又名虫草棒束孢,可入侵包括蜱螨目、鞘翅目、同翅目、鳞翅目等在内的100多种昆虫[18]。环链棒束孢对白蜘蛛Tetranychus cinnabarinus、斜纹夜蛾Spodoptera litura、小菜蛾Plutella xylostella等农业害虫具有较好防效[19-20]。从现有报道来看,棒束孢属真菌更易入侵鳞翅目昆虫,对鞘翅目昆虫防效相对较弱,可能是因为鳞翅目昆虫结构更利于真菌孢子黏附,而鞘翅目昆虫有鞘翅,真菌孢子吸附入侵相对困难。目前鲜见棒束孢属菌对黄曲条跳甲的致病力研究相关报道。华南地区十字花科蔬菜连作指数高,黄曲条跳甲的危害非常严重,已经成为广东近年来防治次数最多的主要害虫之一,给广东农业发展带来了巨大的经济损失[21]。黄曲条跳甲的防治方法主要有物理防治、农业防治、生物防治和化学防治,物理防治主要利用黄板和黑光灯进行诱集杀灭;农业防治主要从耕作方式入手,利用十字花科蔬菜与非十字花科蔬菜轮作,减轻黄曲条跳甲为害;生物防治手段目前仍不够成熟,并没有实现大量的田间应用,如黄曲条跳甲的寄生天敌茧蜂,在施放后并未得到该蜂建立种群的有效依据,有报道称白僵菌、绿僵菌能侵染黄曲条跳甲的幼虫和成虫,但研究仍停留在实验室阶段[22-23];化学防治目前依然是黄曲条跳甲的主要防治手段,主要的化学药剂有噻虫嗪、噻虫胺、阿维菌素、哒螨灵、啶虫脒和氯虫苯甲酰胺等,但由于农户长期大量地不合理施药,黄曲条跳甲对大部分化学农药均产生了较高的抗性[24]。对于黄曲条跳甲的防治,不应局限于某一种单一的防治手段,而应从实际出发,多种防治相结合,从而实现十字花科蔬菜的绿色安全生产。
本研究从我国多个省区不同环境的土壤中分离棒束孢属真菌的菌株,共得到4个种,其中爪哇棒束孢菌株数量最多、分布最广,粉棒束孢、玫烟色棒束孢和环链棒束孢菌株数量较少,说明爪哇棒束孢菌株是优势菌种。从系统发育树来看,从我国不同地区分离得到的棒束孢菌相似性较高,说明我国棒束孢菌种间的遗传差异并不明显,可能是因为采集样品区域棒束孢菌并没有形成地理隔离,其种间仍存在基因交流,也可能由于棒束孢菌对生境依赖性较高,种群稳定性较好,遗传变异发生较少。另有研究表明,棒束孢属对单一的生境有较强的依赖性[25]。从选取的9株菌株对黄曲条跳甲的致病力结果来看,大部分供试菌株对黄曲条跳甲都有一定的致病力,由于黄曲条跳甲幼虫阶段是生活在土壤中为害植株,而化学农药则对土壤中的黄曲条跳甲幼虫防效较差,可以考虑使用虫生真菌来防治土壤中的幼虫,化学农药防治地面上的成虫,化学农药与生防真菌相互结合以控制害虫为害,减少化学农药施用量,进一步减少化学农药对环境的污染,延缓害虫抗药性的发展。
在自然环境中, 虫生真菌的生长、分布与气候条件、土壤性质、动植被丰富度、海拔高度、人类活动以及历史环境变迁等多种因素有关[26]。气候条件丰富、人类活动少的地区更有利于虫生真菌的生长。本研究对菌株的鉴定是从菌落形态、分生孢子结构及ITS序列方面着手的,还需要通过其他方式进一步准确地鉴定菌株的种类,进而了解其生物多样性。
-
![]()
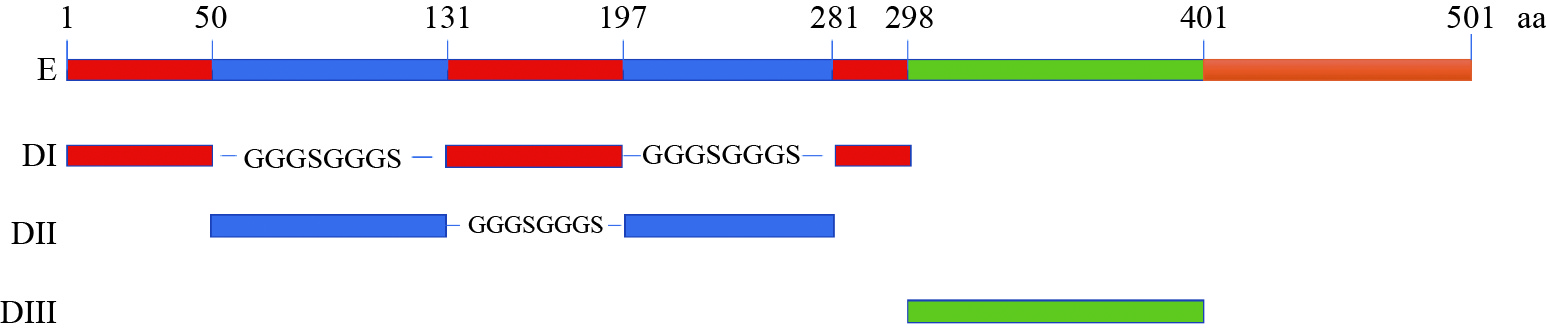
图 1 DTMUV E蛋白及DI、DII、DIII分布
Figure 1. Distribution of DTMUV E protein and DI, DII, DIII
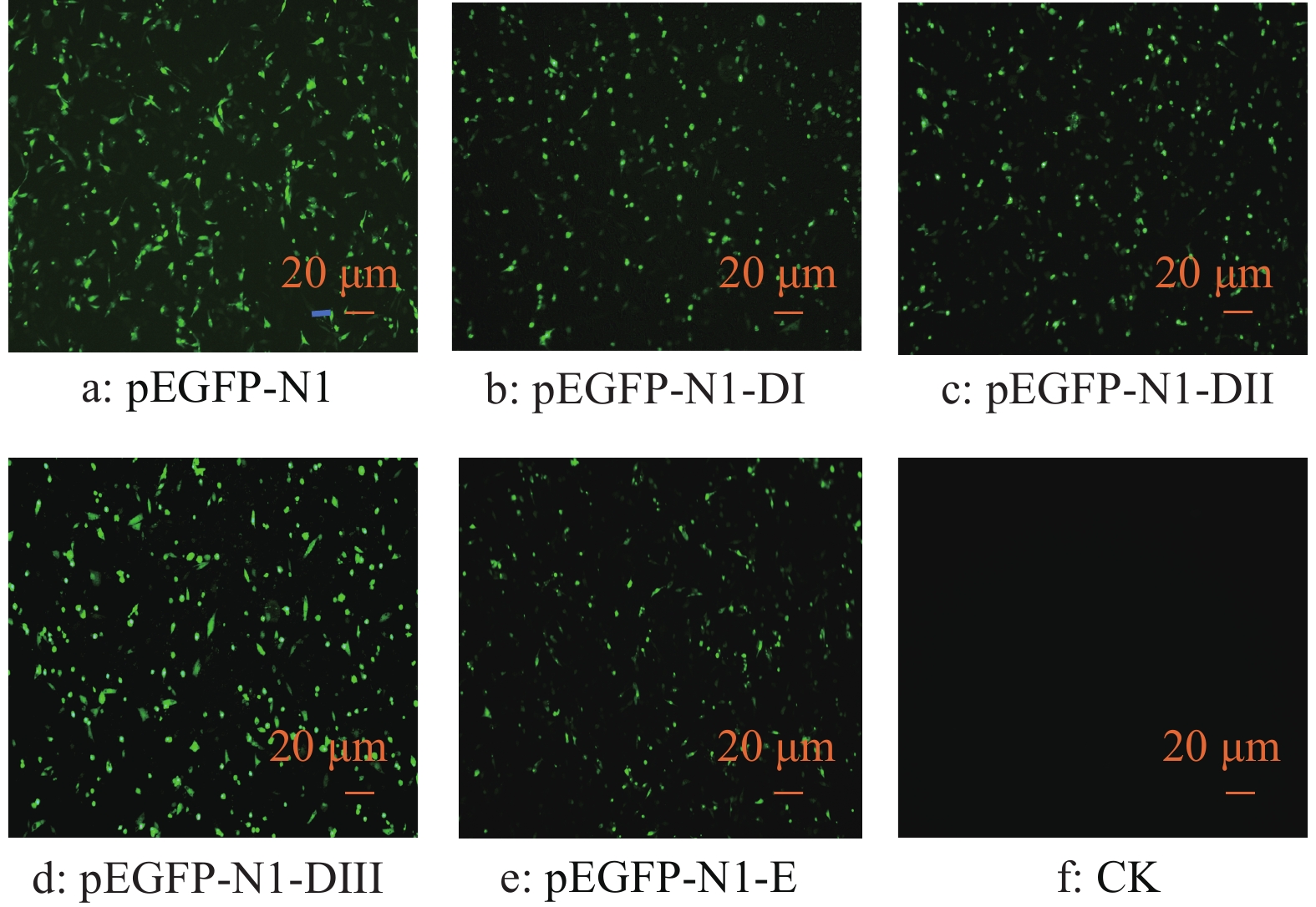
![]()
图 2 不同质粒转染DEF后GFP的表达
Figure 2. GFP expression in DEF transfected by different plasmids
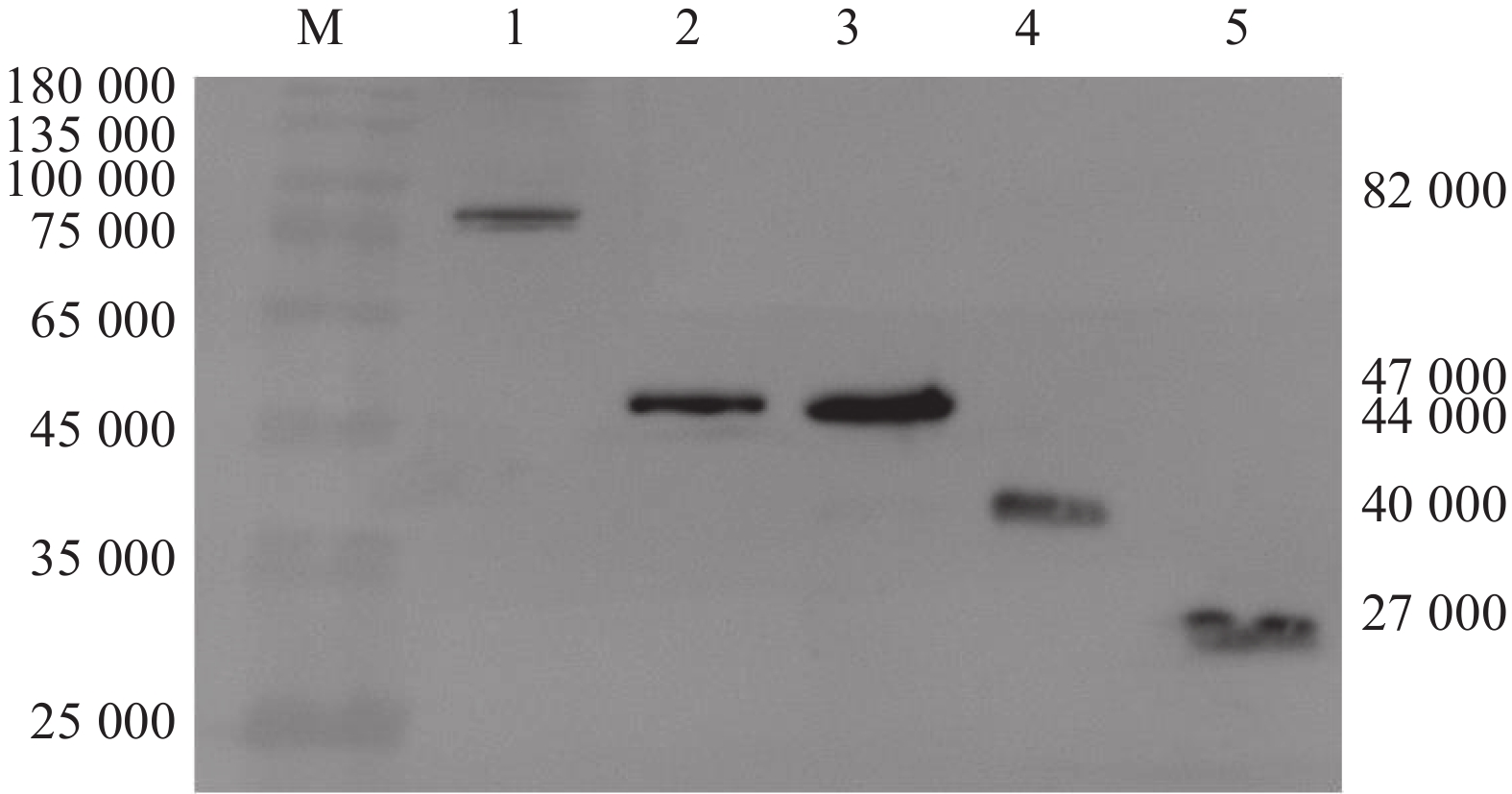
![]()
图 3 Western blot检测E蛋白及其DI、DII、DIII在DEF中的表达
Figure 3. Expression detection of E protein and its DI, DII, DIII in DEF by Western blot
M: Marker, 1: pEGFP-N1-E, 2: pEGFP-N1-DI, 3: pEGFP-N1-DII, 4: pEGFP-N1-DIII, 5: pEGFP-N1
![]()
图 4 质粒转染细胞24 h后DTMUV E蛋白及其DI、DII、DIII真核表达产物对DEF凋亡率的影响
Figure 4. Effect of E protein of DTMUV and its DI, DII, DIII eukaryotic expression products on DEF apoptosis rate after plasmid transfection for 24 h
![]()
图 5 质粒转染细胞36 h后DTMUV E蛋白及其DI、DII、DIII真核表达产物对DEF凋亡率的影响
Figure 5. Effect of E protein of DTMUV and its DI, DII, DIII eukaryotic expression products on DEF apoptosis rate after plasmid transfection for 36 h
![]()
图 6 质粒转染细胞24 h后DTMUV E蛋白及其DI、DII、DIII真核表达产物对DEF细胞周期的影响
G0/G1:细胞停止分裂期与DNA合成前期;G2/M:DNA合成后期与细胞分裂期;S:DNA合成期
Figure 6. Effect of E protein of DTMUV and its DI, DII, DIII eukaryotic expression products on DEF cell cycle after plasmid transfection for 24 h
G0/G1: Cell nondivision phase and DNA presynthetic phase; G2/M: DNA postsynthetic phase and cell division phase; S: DNA synthesis phase
![]()
图 7 质粒转染细胞36 h后DTMUV E蛋白及其DI、DII、DIII真核表达产物对DEF细胞周期的影响
G0/G1:细胞停止分裂期与DNA合成前期;G2/M:DNA合成后期与细胞分裂期;S:DNA合成期
Figure 7. Effect of E protein of DTMUV and its DI, DII, DIII eukaryotic expression products on DEF cell cycle after plasmid transfection for 36 h
G0/G1: Cell nondivision phase and DNA presynthetic phase; G2/M: DNA postsynthetic phase and cell division phase; S: DNA synthesis phase
表 1 DTMUV E蛋白DI、DII、DIII序列
Table 1 Sequence of DI, DII, DIII of DTMUV E protein
基因名称 Gene name 序列设计 Sequence design 长度/bp Length pEGFP-N1-DI CTCGAGATG+50aa+GGGSGGGS+66aa+GGGSGGGS+17aa+CGGGATCC 447 pEGFP-N1-DII CTCGAGATG+81aa+GGGSGGGS+84aa+CGGGATCC 519 pEGFP-N1-DIII CTCGAGATG+103aa+CGGGATCC 312  下载: 导出CSV
下载: 导出CSV
表 2 DTMUV E蛋白及其DI、DII和DIII真核表达产物对DEF细胞周期的影响
Table 2 Effect of DTMUV E protein and its DI, DII, DIII eukaryotic expression products on DEF cell cycle
% t/h 处理 Treatment 细胞比例1) Cell percentage G0/G1 G2/M S 24 pEGFP-N1-E 81.43 13.14 5.43 pEGFP-N1-DI 57.90 17.64 22.58 pEGFP-N1-DII 72.90 14.35 12.75 pEGFP-N1-DIII 72.25 14.95 12.80 pEGFP-N1 81.54 14.08 4.38 CK 63.83 15.89 20.28 36 pEGFP-N1-E 78.52 11.50 9.98 pEGFP-N1-DI 82.33 6.22 11.44 pEGFP-N1-DII 82.47 7.09 10.44 pEGFP-N1-DIII 78.72 10.28 11.00 pEGFP-N1 82.51 7.80 9.69 CK 45.18 11.97 42.84 1) G0/G1:细胞停止分裂期与DNA合成前期;G2/M:DNA合成后期与细胞分裂期;S:DNA合成期 1) G0/G1: Cell nondivision phase and DNA presynthetic phase; G2/M: DNA postsynthetic phase and cell division phase; S: DNA synthesis phase
下载: 导出CSV
-
[1] 张帆帆, 曾艳兵, 方绍培, 等. 鸭坦布苏病毒病的研究进展[J]. 畜牧兽医学报, 2021, 52(6): 1489-1497. [2] 陈仕龙, 陈少莺, 王劭, 等. 一种引起蛋鸡产蛋下降的新型黄病毒的分离与初步鉴定[J]. 福建农业学报, 2011, 26(2): 170-174. [3] 章丽娇. 坦布苏病毒细胞适应毒免疫原性及致弱分子基础研究[D]. 北京: 中国农业大学, 2017. [4] 姜胜男. 鸭DDX3X基因的克隆表达及其抗鸭坦布苏病毒活性研究[D]. 泰安: 山东农业大学, 2020. [5] 董嘉文, 李林林, 孙敏华, 等. 鸭坦布苏病毒JM株E蛋白的截断表达及间接ELISA方法的建立[J]. 中国动物传染病学报, 2017, 25(1): 12-18. [6] LIU M, LIU C, LI G, et al. Complete genomic sequence of duck flavivirus from China[J]. Journal of Virology, 2012, 86(6): 3398-3399. doi: 10.1128/JVI.07086-11
[7] YU K, SHENG Z Z, HUANG B, et al. Structural, antigenic, and evolutionary characterizations of the envelope protein of newly emerging Duck Tembusu Virus[J]. PLoS One, 2013, 8(8): e71319.
[8] ZHANG X, JIA R, SHEN H, et al. Structures and functions of the envelope glycoprotein in flavivirus infections[J]. Viruses, 2017, 9(11): 338. doi: 10.3390/v9110338.
[9] ZHAO D, HUANG X, LIU Y, et al. Domain I and II from newly emerging goose tembusu virus envelope protein functions as a dominant-negative inhibitor of virus infectivity[J]. Research in Veterinary Science, 2015, 98: 121-126. doi: 10.1016/j.rvsc.2014.11.003
[10] 王若洁. 鸭坦布苏病毒E蛋白Domain III的原核表达及其免疫原性的初步研究[D]. 合肥: 安徽农业大学, 2020. [11] 张琳. 鸭坦布苏病毒E蛋白抗原表位的鉴定[D]. 泰安: 山东农业大学, 2013. [12] KOLASKAR A S, KULKARNI-KALE U. Prediction of three-dimensional structure and mapping of conformational epitopes of envelope glycoprotein of Japanese encephalitis virus[J]. Virology, 1999, 261(1): 31-42. doi: 10.1006/viro.1999.9859
[13] 李秀丽, 提金凤, 路云建, 等. 鸭坦布苏病毒对雏鸭免疫系统的影响[J]. 中国兽医学报, 2017, 37(2): 211-217. [14] 提金凤, 李志杰, 刁有祥, 等. 坦布苏病毒诱导昆明鼠脾细胞凋亡[J]. 中国兽医学报, 2021, 41(4): 640-645. [15] PAN Y, CHENG A, ZHANG X, et al. Transcriptome analysis of duck embryo fibroblasts for the dynamic response to duck tembusu virus infection and dual regulation of apoptosis genes[J]. Aging, 2020, 12(17): 17503-17527.
[16] 于观留. 坦布苏病毒感染对鸭胚成纤维细胞生物学功能的影响及其机制研究[D]. 泰安: 山东农业大学, 2019. [17] 林昀. 坦布苏病毒感染的鸭胚成纤维中长链非编码RNA的表达谱变化及其功能初步研究[D]. 泰安: 山东农业大学, 2020. [18] 林委卫, 陈雪明, 李晨曦, 等. 鸭坦布苏病毒E蛋白具有独特交叉反应性中和表位的鉴定[J]. 中国兽医学报, 2021, 41(1): 26-32. -
期刊类型引用(8)
1. 杜丹超,刘顺民,蒲占湑,吕佳,鹿连明. 爪哇棒束孢MSC-F1对柑橘全爪螨的毒力测定及液体发酵工艺优化. 中国生物防治学报. 2025(01): 44-53 .  百度学术
百度学术
2. 鹿连明,杜丹超,胡秀荣,蒲占湑,刘顺民,吕佳,黄振东,陈国庆. 爪哇虫草MSC-f1的分离鉴定及其对多种柑橘害虫的毒力. 植物保护学报. 2025(02): 468-478 . 百度学术
3. 庞素芸,禹豹,杨新泉,胡琼波. 豆天蛾养殖过程中病原真菌的分离与鉴定. 福建农林大学学报(自然科学版). 2024(04): 451-457 . 百度学术
4. 肖勇,吴雨洪,靖湘峰,张杰,李振宇. 我国黄曲条跳甲综合治理研究进展. 植物保护. 2023(02): 22-31+64 . 百度学术
5. 谢梅琼,王龙江,何余容,吕利华. 玫烟色棒束孢转录组测序及潜在致病相关基因分析. 浙江农业学报. 2023(09): 2169-2180 . 百度学术
6. 高新菊,张蒙萌,王恒亮,祖均怀,杨文佳,乔耀淑. 联苯·啶虫脒对黄曲条跳甲的生物活性及防治效果. 农药. 2022(01): 57-60+64 . 百度学术
7. 吴煜,邓娇,张晓娜,黄凯丰,陈庆富. 黑唇平背叶蜂幼虫的形态、危害特征及杀虫菌种的筛选. 西南大学学报(自然科学版). 2022(04): 62-69 . 百度学术
8. 樊春丽,罗来凤,温文照,韦文飞,韦继光. 1株桉大毛虫病原真菌的分离鉴定及其寄主范围测定. 南方农业学报. 2022(08): 2175-2185 . 百度学术
其他类型引用(3)