Identification and fungicide sensitivity of pathogen causing anthracnose of pepper
-
摘要:目的
明确辣椒炭疽病病原菌Colletotrichum spp.种类,并筛选防治药剂。
方法采集贵州省贵阳市花溪区具有典型炭疽病症状的‘党武’辣椒叶片和果实,分离病原菌;采用单孢分离法、柯赫氏法则、形态学特征结合ITS、GADPH、CHS-1和ACT多基因序列分析,明确病原菌的致病能力及分类地位;采用菌丝生长速率法测定病原菌对6种化学杀菌剂和6种生物杀菌剂的敏感性。
结果病原菌鉴定为斯高维尔炭疽菌Colletotrichum scovillei;室内药剂敏感性测定发现12种杀菌剂对C. scovillei均有一定的抑制作用,75%(w)肟菌·戊唑醇WDG、10%(w)苯醚甲环唑WDG和250 g/L吡唑醚菌酯SC的抑制效果最好,EC50分别为0.254、0.731、0.745 mg/L。其次是200 g/L异硫氰酸烯丙酯SL、3%(w)中生菌素WP、10 g/L申嗪霉素SC、80 g/L宁南霉素AS、200 g/L异硫氰酸烯丙酯EW和10 g/L蛇床子素ME,EC50分别为1.238、1.307、1.711、2.929、3.175和2.191 mg/L。将10%(w)苯醚甲环唑WDG和250 g/L吡唑醚菌酯SC按照1∶4、2∶3的体积比复配时,联合毒力测定和评价结果显示两者复配对该病菌具有协同增效作用。
结论本研究可为辣椒生产中辣椒炭疽病的田间药剂防治提供参考。
Abstract:ObjectiveTo identify the pathogenic fungi of pepper anthracnose and screen the fungicides with strong inhibitory effects.
MethodThe typical leaves and fruits with anthracnose symptoms were collected from the peppers named ‘Dangwu’ in Huaxi, Guiyang, Guizhou. The pathogen was isolated. Its pathogenicity and classification were determined by monospore isolation, Koch’s postulates, and morphological characteristics combined with multi-locus (ITS, ACT, GAPDH andCHS-1) phylogenetic analysis. The sensitivities of pathogen to six chemical fungicides and six biological fungicides were investigated by the mycelial growth rate method.
ResultThe pathogen causing pepper ‘Dangwu’ anthracnose in Huaxi was identified as Colletotrichum scovillei. The indoor fungicide sensitivity test showed that 12 fungicides all had certain inhibitory effect against C. scovillei. The inhibition effects of 75%(w) trifloxystrobin·tebuconazole WDG, 10%(w) difenoconazole WDG and 250 g/L pyraclostrobin SC were higher with the EC50 of 0.254, 0.731 and 0.745 mg/L, respectively, followed by 200 g/L allylisothiocyanate SL, 3% (w) zhongshengmycin WP, 10 g/L phenazine-1-carboxylic acid SC, 80 g/L ningnanmycin AS, 200 g/L allylisothiocyanate EW and 10 g/L osthol ME with the EC50 of 1.238, 1.307, 1.711, 2.929, 3.175 and 2.191 mg/L, respectively. 10% (w) difenoconazole WDG and 250 g/L pyraclostrobin SC mixed at the volume ratio of 1∶4 or 2∶3 showed an obvious synergistic effect on inhibiting the pathogen based on the joint toxicity test and evaluation results.
ConclusionThis study can provide references for the field prevention and control of pepper anthracnose using pesticides.
-
动物的采食行为是维持机体能量稳态的基础,畜禽生产中获得充足的食物是其生长发育的前提。动物采食量受中枢调控,其中胃肠道状态是决定畜禽食欲的关键部位。揭示饥饿状态下鸡食欲调控的潜在肠−脑轴机制可为如何提高鸡采食量提供理论依据。下丘脑弓状核作为食欲调控中枢[1-2]存在大量的促采食的刺鼠相关蛋白(Agouti-related protein,AgRP)/神经肽 Y(Neuropeptide Y,NPY)神经元和抑采食的前阿片黑色皮质素(Proopiomelanocortin,POMC)/可卡因−苯丙胺调节转录肽(Cocaine and amphetamine regulated transcript,CART)神经元[3-6]。影响动物食欲的因素有很多,遗传、环境因素、机体健康以及肠道充盈状态等均能影响动物采食量[7]。其中胃肠道作为营养物质暂时储存和消化吸收的关键部位,存在大量食欲调控信号[8]。这些食欲调控信号一方面通过血液循环被中枢所感应[2, 9],另一方面被肠道迷走感觉传入神经元直接感应,经脑干孤束核最终将信号投递至食欲调控中枢,肠道和中枢间的这种信息传递被称为“肠−脑轴” [8, 10]。肠道迷走感觉神经作为假单极双向神经元,位于结状神经节处的胞体分别向中枢孤束核和肠道发出轴突,其中肠道迷走神经末梢存在多种受体感应肠道各种理化信号,例如游离脂肪酸受体2 (FFAR2)、生长激素促分泌素受体(GHSR)、胆囊收缩素受体(CCKR),以及炎症受体TLR4等 [10-12]。
肠道健康对机体维持高食欲具有重要作用,维持肠道平衡可以维持机体正常食欲,反之肠道菌群紊乱等原因导致肠道健康受损则会引起采食量降低[13-14],而肠道屏障是肠道发挥其正常生物学功能的重要前提[15-16],肠道黏膜屏障包括肠上皮细胞及胞间连接,其中,紧密连接是肠上皮细胞间的细胞旁通路的主要屏障,闭合(Occludin)和紧密连接(Claudin)家族成员是影响其功能的主要封闭蛋白,二者与胞浆蛋白相互作用维持紧密蛋白的完整性[17-18]。当肠道出现炎症时,会导致Claudin蛋白结构变化,进而引起肠道屏障功能性障碍,并且受致病菌侵害也会导致肠道屏障通透性增加[19];动物炎症性肠病会导致肠道隐窝改变、小肠绒毛萎缩或变平以及一系列的形态学变化[20]。大量研究发现间歇性饥饿有助于维持肠道及肠道屏障的完整性[21-22]。
此外,胃肠道中上皮基质和微生物群落共调控生成活性氧,导致生成H2O2;而H2O2是维持正常细胞稳态和生理功能所必需的第二信使[23]。Miller等[24]研究发现,结肠内壁中的细胞会释放H2O2(而非氧气)来限制微生物的生长,H2O2可以协同其他物质在肠道黏膜上形成保护,防止菌群紊乱或肠道炎症对机体造成损伤,并且可以治疗肠道炎症,恢复机体正常生理功能。然而,目前并不清楚短期饥饿是否影响肠道炎症水平和屏障功能、是否被迷走感觉神经所感应。本研究旨在揭示禁食后肠道炎症水平和肠道屏障变化,以及提高食欲的潜在机制,并提供理论基础和试验依据。
1. 材料与方法
1.1 试验动物与试验设计
选用20只1日龄初生黄羽肉鸡[25-28](购于广东省清远市凤翔麻鸡发展有限公司生产基地),试验前称体质量并排序,随后按配对随机设计的原则将体质量相近的小鼠分为2组:对照组和禁食组,每组10只黄羽肉鸡,正常饲喂饲料至5日龄并采样。采样前12 h,禁食组禁食,对照组正常采食。禁食12 h后收集小肠肠道内容物检测H2O2水平,采集黄羽肉鸡结状神经节(Nodose ganglia,NG),检测炎症和食欲相关受体的表达;采集十二指肠、空肠和回肠及其肠道黏膜,检测黄羽肉鸡肠道形态、闭锁小带蛋白−1 (Zonula,ZO-1)、闭合蛋白 (Occludens-1,OCC)、紧密连接蛋白(Claudin-1) 以及炎症因子的表达。
1.2 测定指标与方法
1.2.1 小肠肠道内容物
分离小肠,区分十二指肠、空肠和回肠,取部分肠道轻轻挤压,将内容物收集于 2 mL 离心管中,使用过氧化氢测定试剂盒(A064-1-1,南京建成生物工程研究所)检测H2O2水平。
1.2.2 扫描电子显微镜(SEM)
取一段1 cm长的空肠,剪开后平铺,用生理盐水轻轻清洗内容物,而后修剪为5 mm边长的正方形放于保存液中,于4 ℃条件下保存。而后脱水、干燥,进行电镜扫描。
1.2.3 苏木精−伊红(HE) 染色
小肠分离后剪取约 3 cm 空肠中段放于 40 g/L 的多聚甲醛中固定,按照常规方法制作石蜡切片,HE染色,光学显微镜下拍照,然后用Image软件测取肠道绒毛长度(lv)和隐窝深度(dc),每个切片取 3~5 个视野,取其平均值计算绒毛长度与隐窝深度比值(lv/dc)。
1.2.4 小肠黏膜及 NG 的 RNA 提取、逆转录和荧光定量PCR (q-PCR)
小肠黏膜及NG总 RNA 使用 RNA 提取试剂盒(R4130-02,广州美基生物科技有限公司)和 TRIzol 试剂提取。1 g 总 RNA 按试剂盒说明书用 4× Reverse Transcription Master Mix(EZB-RT2GQ,美国 EZBioscience 生物技术有限公司)逆转录成 cDNA。引物序列见表1,按照2× SYBR Green qPCR Master Mix(A0012-R2,美国 EZBioscience 生物技术有限公司)说明书配制反应体系:10 μL 的体系中含有 5 μL 2× Color SYBR Green qPCR Master Mix、3.6 μL dd H2O、1 μL cDNA、0.4 μL 引物工作液;使用 Applied Biosystems QuantStudio 3 实时 PCR 系统并按照以下程序反应:95 ℃预热 5 min;95 ℃ 10 s,60 ℃ 30 s,循环 40 次。根据对照组 β-actin mRNA 表达进行归一化处理[15]。
表 1 实时荧光定量PCR所用引物Table 1. Primers used for quantitative real-time PCR基因
Gene上游引物序列(5′→3′)
Forward primer sequence下游引物序列(5′→3′)
Reverse primer sequence序列号
Accession numberβ-actin CTGTGCCCATCTATGAAGGCTA ATTTCTCTCTCGGCTGTGGTG L08165 AgRP CTCTTCCCAGGCCAGACTTG GCAGAAGGCGTTGAAGAACC XM_046925680.1 CCKAR AGCTCTTCTGCCAACCTGAT GTGTAGGACAGCAGGTGGAT NM_001081501.2 Claudin-1 TGGAGGATGACCAGGTGAAG TGTGAAAGGGTCATAGAAGG NM_001013611.2 CART CGAGAGAAGGAGCTGATCGA AGAAAGGAGTTGCACGAGGT XM_046937244.1 FFAR2 GCACTCTCTTTATGGCTGCC GGATTCCCTGGTCTTGGTCA XM_040693461.2 IL-1 CCTCCTCCAGCCAGAAAGTG CGGTAGAAGATGAAGCGGGT XM_015297469.3 IL-4 CCCCAGGTGTAGGCTCTAGT ACTCTGTCATTGCTGCTCCC XM_040683457.2 IL-6 ACCCGAGCTCTTTGGTGATG CGTGCCCTCTGTTTGTACCT XM_025143427.3 IL-10 GCTGCCAAGCCCTGTT CCTCAAACTTCACCCTCA NM_001004414.4 GHSR ATTAGTGCTGGCCCCATCTT CGGACCGATGTTCTTCCTCT XM_046923539.1 MC4R AGGGGTCATCATCACATGCA GATGGCCCCTTTCATGTTGG NM_001031514.2 NPY GTGCTGACTTTCGCCTTGTC ATCTCTGCCTGGTGATGAGG NM_205473.2 Occludin TGGAGGAGTGGGTGAAGAAC ATCCTTCCCCTTCTCCTCCT XM_046904540.1 POMC AGAGGAAGGCGAGGAGGAAA GTAGGCGCTTTTGACGATGG XM_046914234.1 TLR-4 GGCTCAACCTCACGTTGGTA AGTCCGTTCTGAAATCCCGT NM_001030693.2 TNF-α TTCTATGACCGCCCAGTT CAGAGCATCCAACGCAAAA XM_046920820.1 NPY2R GGCCATCATCTCCTATGCCT GGAAGCCAACTGACAGCAAA NM_001398092.1 ZO-1 TCATCCTTACCGCCGCATAT GTTGACTGCTCGTACTCCCT XM_046925214.1 1.3 数据统计与分析
所有数据均以平均值±标准误差(Mean±SE)表示。用GraphPad Prism 8.0 软件进行统计分析。采用 t 检验对2组均值进行差异显著性分析。
2. 结果与分析
2.1 禁食后下丘脑内食欲肽相关受体表达变化
通过 q-PCR 检测下丘脑内食欲肽相关基因表达,结果发现,与对照组相比,雏鸡禁食12 h后促采食食欲肽基因AgRP (P<0.05)和 NPY (P<0.01)的 mRNA 相对表达量均显著上调(图1),提示雏鸡饥饿模型构建成功。
![图 1 黄羽肉鸡禁食12 h后下丘脑内食欲肽相关受体表达的变化]() 图 1 黄羽肉鸡禁食12 h后下丘脑内食欲肽相关受体表达的变化“*”和“**”分别表示差异达到 0.05和0.01的显著水平(t检验)Figure 1. Expression changes of orexin-related receptors in hypothalamus of yellow-feathered broilers after 12 h of fasting“*” and “**” indicate that the difference reaches 0.05 and 0.01 significance levels respectively (t test)
图 1 黄羽肉鸡禁食12 h后下丘脑内食欲肽相关受体表达的变化“*”和“**”分别表示差异达到 0.05和0.01的显著水平(t检验)Figure 1. Expression changes of orexin-related receptors in hypothalamus of yellow-feathered broilers after 12 h of fasting“*” and “**” indicate that the difference reaches 0.05 and 0.01 significance levels respectively (t test)2.2 禁食对黄羽肉鸡空肠肠道形态的影响
空肠肠绒毛电镜扫描及分析结果如图2A、3A、3B 所示,观察发现雏鸡禁食12 h 后,同对照组相比空肠肠绒毛表面更加完整,单位面积内绒毛总数更多、受损更少并且排列更加整齐。空肠 HE 染色及分析结果如图2B、3C、3D 所示,与正常采食的雏鸡相比,禁食后雏鸡的隐窝深度和lv/dc均无明显变化,但是对照组绒毛有明显损伤,而禁食组绒毛排列整齐、长度更长。
![图 2 黄羽肉鸡禁食12 h对空肠肠道绒毛形态的影响]() 图 2 黄羽肉鸡禁食12 h对空肠肠道绒毛形态的影响Figure 2. Effects of fasting for 12 h on jejunum intestinal villus morphology of yellow-feathered broilers
图 2 黄羽肉鸡禁食12 h对空肠肠道绒毛形态的影响Figure 2. Effects of fasting for 12 h on jejunum intestinal villus morphology of yellow-feathered broilers![图 3 黄羽肉鸡禁食12 h后空肠肠道绒毛形态变化的电镜扫描结果(A、B)和HE 染色结果(C、D)统计]() 图 3 黄羽肉鸡禁食12 h后空肠肠道绒毛形态变化的电镜扫描结果(A、B)和HE 染色结果(C、D)统计Ⅰ:对照组,Ⅱ:禁食组;“*”和“**”分别表示差异达到 0.05和0.01的显著水平(t检验)Figure 3. Statistics of the scanning electron microscopy results (A, B) and HE staining results (C, D) for the morphological changes of jejunum intestinal villi of yellow-feathered broilers after fasting for 12 hⅠ: Control, Ⅱ: Fasting group; “*” and “**” indicate that the difference reaches 0.05 and 0.01 significance levels respectively (t test)
图 3 黄羽肉鸡禁食12 h后空肠肠道绒毛形态变化的电镜扫描结果(A、B)和HE 染色结果(C、D)统计Ⅰ:对照组,Ⅱ:禁食组;“*”和“**”分别表示差异达到 0.05和0.01的显著水平(t检验)Figure 3. Statistics of the scanning electron microscopy results (A, B) and HE staining results (C, D) for the morphological changes of jejunum intestinal villi of yellow-feathered broilers after fasting for 12 hⅠ: Control, Ⅱ: Fasting group; “*” and “**” indicate that the difference reaches 0.05 and 0.01 significance levels respectively (t test)2.3 禁食对黄羽肉鸡肠道屏障的影响
由图4 可知,与对照组相比,禁食12 h后雏鸡小肠黏膜中紧密蛋白标志性基因ZO-1和Occludin mRNA的相对表达量均显著上调(P<0.05),在十二指肠中,Claudin-1 的mRNA相对表达量也显著上调(P<0.05)。
![图 4 黄羽肉鸡禁食12 h后小肠肠道黏膜紧密蛋白的mRNA相对表达量变化]() 图 4 黄羽肉鸡禁食12 h后小肠肠道黏膜紧密蛋白的mRNA相对表达量变化“*”和“**”分别表示差异达到 0.05和0.01的显著水平(t检验)Figure 4. mRNA relative expression changes of intestinal mucosal compact protein in small intestine of yellow-feathered broilers after 12 h fasting“*” and “**” indicate that the difference reaches 0.05 and 0.01 significance levels respectively (t test)
图 4 黄羽肉鸡禁食12 h后小肠肠道黏膜紧密蛋白的mRNA相对表达量变化“*”和“**”分别表示差异达到 0.05和0.01的显著水平(t检验)Figure 4. mRNA relative expression changes of intestinal mucosal compact protein in small intestine of yellow-feathered broilers after 12 h fasting“*” and “**” indicate that the difference reaches 0.05 and 0.01 significance levels respectively (t test)2.4 禁食对黄羽肉鸡肠道炎症水平的影响
由图5可知,黄羽肉鸡禁食12 h后,与对照组相比,十二指肠、空肠和回肠黏膜上炎症因子IL-1、IL-6和TNF-α的 mRNA表达量无明显变化,但是空肠黏膜抗炎因子IL-4和IL-10的 mRNA表达量均有显著升高(P<0.01)。并且空肠和回肠内容物中H2O2浓度均有不同程度的增加(图3 D )。
![图 5 黄羽肉鸡禁食12 h后小肠炎症因子mRNA相对表达量及H2O2浓度变化]() 图 5 黄羽肉鸡禁食12 h后小肠炎症因子mRNA相对表达量及H2O2浓度变化图D中,DU:十二指肠,Anterior JE:空肠前段,Middle JE:空肠中段,Posterior JE:空肠后段,Anterior IL:回肠前段,Posterior IL:回肠后段;“*”和“**”分别表示差异达到 0.05和0.01的显著水平(t检验)Figure 5. Changes in mRNA relative expressions of intestinal inflammatory factors and H2O2 concentrations in yellow-feathered broilers after 12 h of fastingIn figure D, DU: Duodenum, Anterior JE: Anterior jejunum, Middle JE: Middle jejunum, Posterior JE: Posterior jejunum, Anterior IL: Anterior ileum , Posterior IL: Posterior ileum; “*” and “**” indicate that the difference reaches 0.05 and 0.01 significance levels respectively (t test)
图 5 黄羽肉鸡禁食12 h后小肠炎症因子mRNA相对表达量及H2O2浓度变化图D中,DU:十二指肠,Anterior JE:空肠前段,Middle JE:空肠中段,Posterior JE:空肠后段,Anterior IL:回肠前段,Posterior IL:回肠后段;“*”和“**”分别表示差异达到 0.05和0.01的显著水平(t检验)Figure 5. Changes in mRNA relative expressions of intestinal inflammatory factors and H2O2 concentrations in yellow-feathered broilers after 12 h of fastingIn figure D, DU: Duodenum, Anterior JE: Anterior jejunum, Middle JE: Middle jejunum, Posterior JE: Posterior jejunum, Anterior IL: Anterior ileum , Posterior IL: Posterior ileum; “*” and “**” indicate that the difference reaches 0.05 and 0.01 significance levels respectively (t test)2.5 禁食后雏鸡NG内受体表达的变化
由图6A 可知,与对照组相比,雏鸡禁食12 h后 NG 内肠道炎症因子IL-4的受体基因IL-4R的mRNA相对表达量显著上调(P<0.01)。由图6B 可知,禁食组雏鸡NG内食欲相关受体基因的mRNA相对表达量有所增加,其中FFAR2和神经肽2受体(NPY2R)表达量增加显著(P<0.01)。
![图 6 黄羽肉鸡禁食12 h后结状神经节内炎症(A)与食欲(B)相关受体mRNA相对表达量]() 图 6 黄羽肉鸡禁食12 h后结状神经节内炎症(A)与食欲(B)相关受体mRNA相对表达量“**”表示差异达到0.01的显著水平(t检验)Figure 6. mRNA relative expression of inflammation-related (A) and orexin-related (B) receptors in nodose ganglia of yellow-feathered broilers after 12 h of fasting“**” indicates that the difference reaches 0.01 significance level (t test)
图 6 黄羽肉鸡禁食12 h后结状神经节内炎症(A)与食欲(B)相关受体mRNA相对表达量“**”表示差异达到0.01的显著水平(t检验)Figure 6. mRNA relative expression of inflammation-related (A) and orexin-related (B) receptors in nodose ganglia of yellow-feathered broilers after 12 h of fasting“**” indicates that the difference reaches 0.01 significance level (t test)3. 讨论与结论
已有研究发现,特异性激活下丘脑弓状核AgRP神经元显著提高动物采食量[29],诱导肥胖发生[30],而消除AgRP神经元则会导致厌食症[31]。因此,本研究首先检测了下丘脑弓状核食欲肽表达变化,结果发现短期禁食后黄羽肉鸡下丘脑 AgRP/NPY表达显著上调(P < 0.05),而POMC有下降趋势(P = 0.07),提示黄羽肉鸡饥饿模型构建成功。
肠道健康对机体维持高食欲具有重要作用,而肠道炎症则会影响肠道代谢水平、破坏微生物平衡[32]以及肠道屏障的完整性[33],甚至会影响中枢神经系统中神经肽的分泌,大量研究发现间歇性饥饿有助于维持肠道及肠道屏障的完整性[21-22]。据报道,胃肠道中上皮基质和微生物群落共调控生成活性氧,导致H2O2形成;而H2O2是维持正常细胞稳态和生理功能所必需的第二信使[23]。本试验通过检测小肠不同肠段内容物的H2O2浓度发现,短期禁食导致禁食组空肠和回肠内容物中H2O2浓度均有不同程度的增加,推测饥饿状态下肠道可能通过生成适量H2O2维持肠道稳定。为进一步验证这一假设,我们通过电镜扫描、HE染色以及q-PCR结果发现,短期禁食并未对肠道形态造成损伤,且由于缺少食物影响,肠道绒毛排列更加紧凑整齐。我们推测,机体短期禁食后尚未引发肠道疾病,并且在肠道饥饿状态下,因肠道营养物质缺乏,机体可能出于自我保护机制防止肠道毒素等有害因子进入机体,从而紧密连接增强,即肠道物理屏障增强,且抗炎因子的表达增加,降低空肠损伤比例,避免肠道受损,以抵抗禁食给机体带来的不良影响,维持肠道正常的生理功能,这对维持较高食欲至关重要。
大量研究报道,肠道食欲调控信号不仅可以通过血液信号被中枢所识别,还可以被肠道迷走感觉传入神经元直接感应,经肠−脑轴最终将信号投递至食欲调控中枢[10-12]。本试验结果发现,与对照组相比,雏鸡禁食12 h后结状神经节内IL-4受体基因的mRNA相对表达量显著上调,和肠道黏膜抗炎因子表达变化相对应;提示机体在饥饿状态下,可能通过提高肠道抗炎能力以及增强物理屏障来抵抗由禁食所导致的轻微炎症,维持肠道健康。
此外,结状神经节内食欲相关受体基因的mRNA相对表达量有所增加,其中FFAR2和NPY2R表达量增加显著(P < 0.01),推测黄羽肉鸡饥饿后由于AgRP和NPY表达量增加[4],并且FFAR2和NPY2R表达增加,二者将肠道饥饿信号传递至中枢神经系统,提高动物食欲进而促进采食量增加。
综上所述,饥饿可引起肠道抗炎因子水平升高,并维持肠道屏障完整性,同时促进迷走感觉神经末梢抗炎因子受体表达,最终引起食欲增强。
-
![]()
图 1 菌株HGUP LJ169致病性测定
A和C:无菌水对照;B:刺伤法接种叶片发病症状;D:刺伤法接种果实发病症状
Figure 1. Pathogenicity test of HGUP LJ169
A, C: Sterile water control; B: Symptoms of leaf onset by prick inoculation; D: Symptoms of fruit onset by prick inoculation
![]()
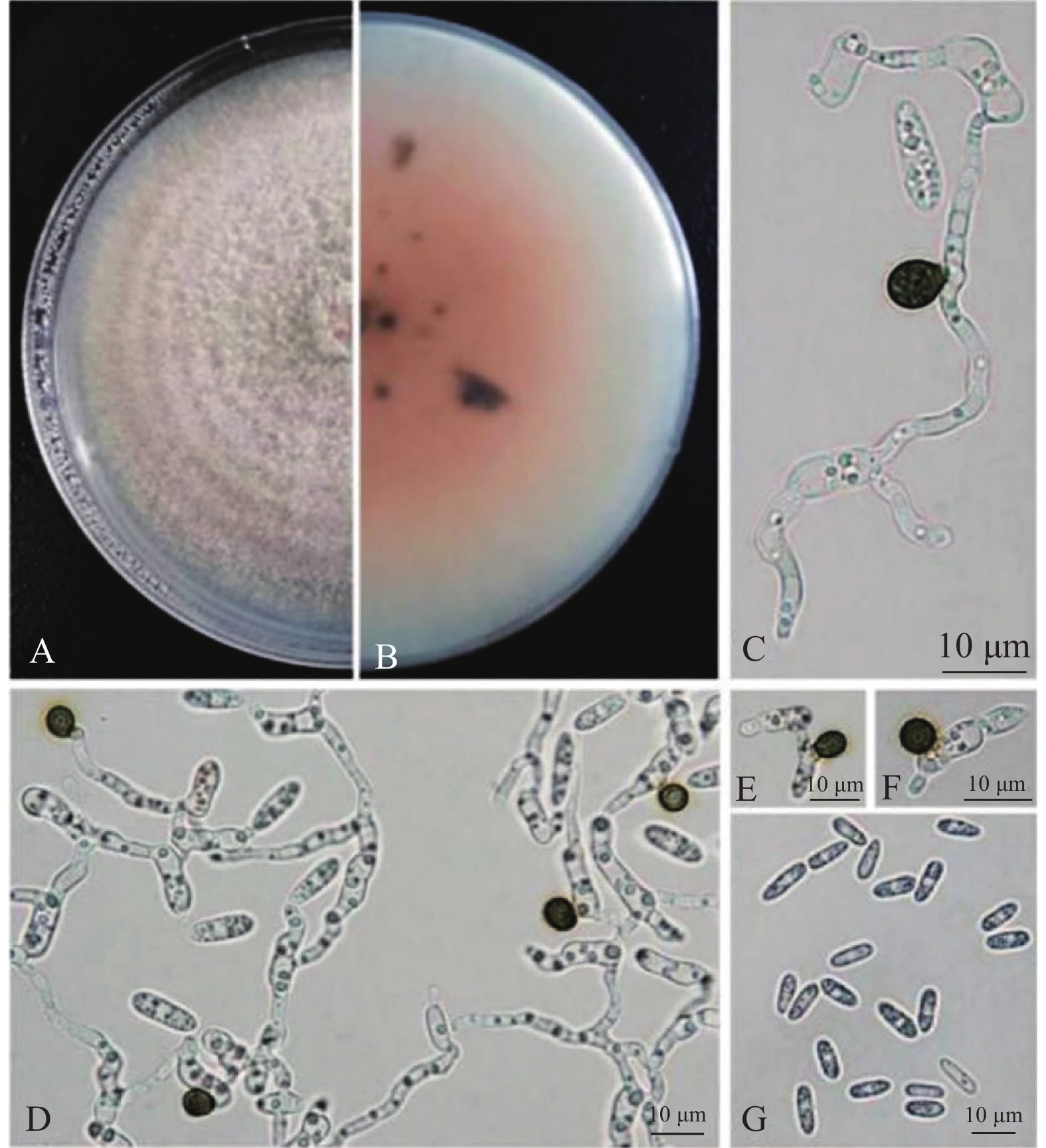
图 2 菌株HGUP LJ169的形态特征
A、B:菌落形态;C~F:分生孢子附着胞;G: 分生孢子
Figure 2. Morphological characteristics of HGUP LJ169
A, B: Colony morphology; C-F: Appressorium; G: Conidia
![]()
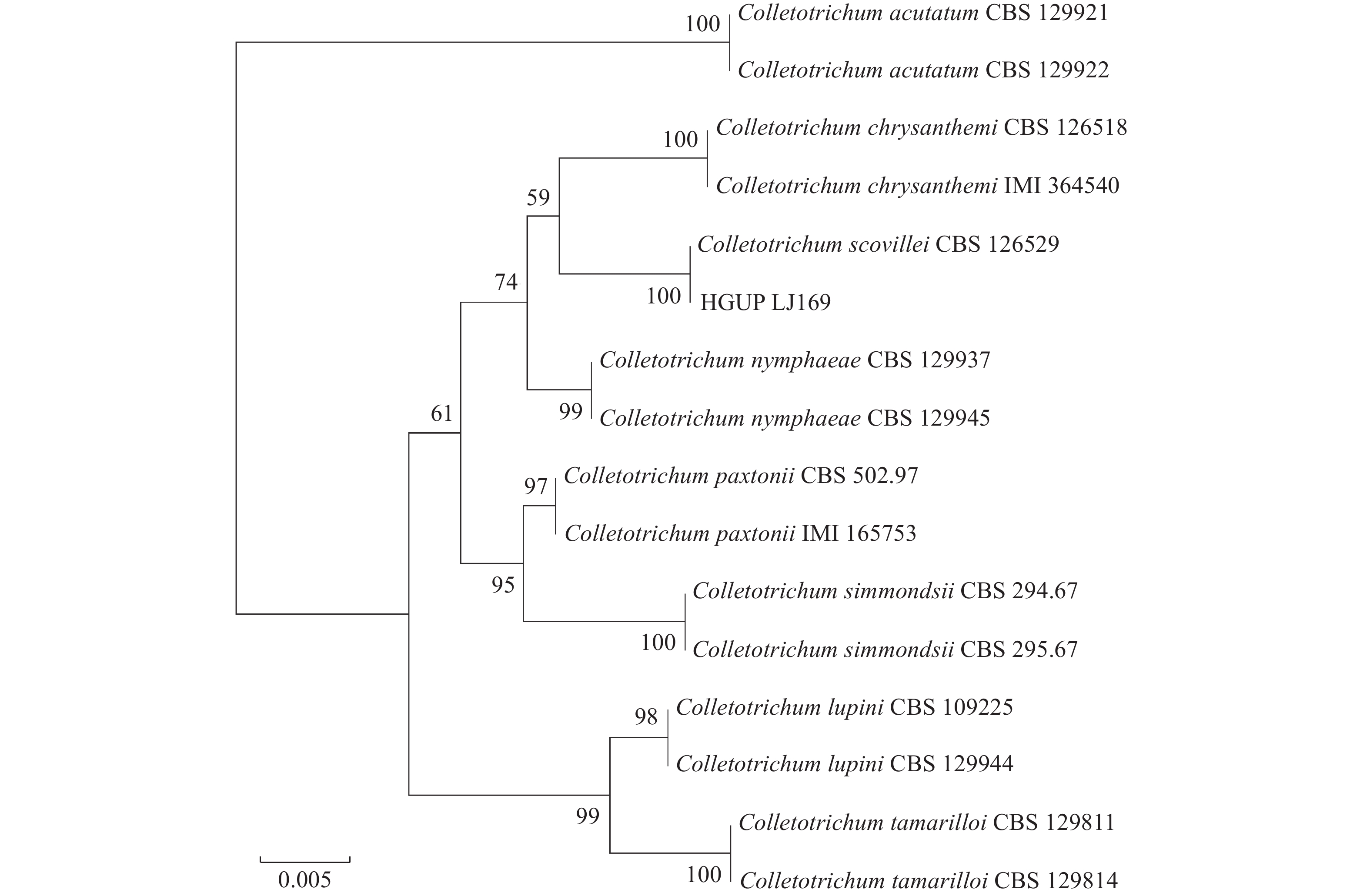
图 3 基于ITS、ACT、CHS-1、GAPDH基因序列的菌株系统发育树
标尺表示遗传距离,分支上的数字表示支持率(%)
Figure 3. Phylogenetic tree of bacterial strains based on ITS, GADPH, CHS-1 and ACT gene sequences
The scale bar shows the genetic distance,and the number on the node shows the Bootstrap support rate (%)
表 1 本研究所用的引物
Table 1 Primers used in this study
基因 Gene 引物 Primer 引物序列 (5′→3′) Primer sequence ITS ITS1[15] TCCGTAGGTGAACCTGCGG ITS4[15] TCCTCCGCTTATTGATATGC ACT ACT-512F[16] ATGTGCAAGGCCGGTTTCGC ACT-783R[16] TACGAGTCCTTCTGGCCCAT CHS-1 CHS-79F[16] TGGGGCAAGGATGCTTGGAAGAAG CHS-354R[16] TGGAAGAACCATCTGTGAGAGTTG GAPDH gpd1[17] CAACGGCTTCGGTCGCATTG gpd2[17] GCCAAGCAGTTGGTTGTGC  下载: 导出CSV
下载: 导出CSV
表 2 多基因序列分析所用菌株信息1)
Table 2 Species used for multi-gene phylogeny analysis in this study
种名 Species 菌株编号 Strain No. 序列号 GenBank No. ITS GAPDH CHS-1 ACT Colletotrichum acutatum CBS 129921 JQ948380 JQ948711 JQ949041 JQ949701 CBS 129922 JQ948381 JQ948712 JQ949042 JQ949702 C. chrysanthemi IMI 364540 JQ948273 JQ948603 JQ948934 JQ949594 CBS 126518 JQ948271 JQ948601 JQ948932 JQ949592 C. fioriniae CBS 127600 JQ948308 JQ948638 JQ948969 JQ949629 CBS 127599 JQ948309 JQ948639 JQ948970 JQ949630 C. lupini CBS 109225 NR111730 JQ948485 JQ948816 JQ949476 CBS 129944 MH865693 JQ948508 JQ948839 JQ949499 C. nymphaeae CBS 129945 MH865694 JQ948531 JQ948862 JQ949522 CBS 129937 MH865686 JQ948583 JQ948914 JQ949574 C. paxtonii CBS 502.97 JQ948286 JQ948616 JQ948947 JQ949607 IMI 165753 JQ948285 JQ948615 JQ948946 JQ949606 C. scovillei CBS 126529 NR111737 JQ948597 JQ948928 JQ949588 HGUP LJ169 MW741882 MW751812 MW751811 MW751810 C. simmondsii CBS 294.67 JQ948277 JQ948607 JQ948938 JQ949598 CBS 295.67 JQ948278 JQ948608 JQ948939 JQ949599 C. tamarilloi CBS 129814 JQ948184 JQ948514 JQ948845 JQ949505 CBS 129811 JQ948185 JQ948515 JQ948846 JQ949506 1)加粗字体表示本研究分离得到的菌株 1)The bold fonts show the isolated strain of this study
下载: 导出CSV
表 3 供试杀菌剂及有效成分质量浓度
Table 3 Fungicides and the contentrations of active ingredients used in this study
杀菌剂 Fungicide 有效成分质量浓度/(mg·L−1) Concentration of active ingredient 药剂类型 Type of drug 75% (w)肟菌·戊唑醇 WDG 75% (w) Trifloxystrobin·tebuconazole WDG 0.075 0.150 0.300 0.600 1.200 甲氧基氨基甲酸酯类+三唑类 Methoxycarbamate carbamate+triazol 10% (w)苯醚甲环唑 WDG 10% (w) Difenoconazole WDG 0.15 0.30 0.60 1.20 2.40 三唑类 Triazol 250 g/L吡唑醚菌酯 SC 250 g/L Pyraclostrobin SC 0.15 0.30 0.60 1.20 2.40 甲氧基氨基甲酸酯类 Methoxycarbamate carbamate 227 g/L二氰蒽醌 SC 227 g/L Dithianon SC 37.50 75.00 150.00 300.00 600.00 醌类 Quinones 500 g/L丙环唑 ME 500 g/L Propiconazol ME 16.70 33.40 66.80 133.60 267.20 三唑类 Triazol 250 g/L溴菌腈 EC 250 g/L Bromothalonil EC 14.00 28.00 56.00 112.00 224.00 溴代氰烷烃类 Bromocyanoalkanes 10 g/L蛇床子素 ME 10 g/L Osthole ME 0.98 3.91 15.63 62.50 250.00 植物源 Plant source 10 g/L申嗪霉素 SC 10 g/L Phenazine-1-carboxylic acid SC 0.98 3.91 15.63 62.50 250.00 微生物源 Microorganism source 80 g/L宁南霉素 AS 80 g/L Ningnanmycin AS 0.98 3.91 15.63 62.50 250.00 微生物源 Microorganism source 3% (w)中生菌素 WP 3% (w) Zhongshengmycin WP 0.98 3.91 15.63 62.50 250.00 微生物源 Microorganism source 200 g/L异硫氰酸烯丙酯 EW 200 g/L Allylisothiocyanate EW 0.80 1.60 3.20 6.40 12.80 植物源 Plant source 200 g/L异硫氰酸烯丙酯 SL 200 g/L Allylisothiocyanate SL 0.40 0.80 1.60 3.20 6.40 植物源 Plant source
下载: 导出CSV
表 4 12种杀菌剂对Colletotrichum scovillei的抑制效果
Table 4 Inhibitory effects of twelve fungicides against Colletotrichum scovillei
杀菌剂 Fungicide 毒力回归方程1)Regression equation of toxicity EC50/ (mg·L−1) EC50 95%置信限/ (mg·L−1) 95% confidence limit of EC50 相关系数 (R) Correlation coefficient 75% (w)肟菌·戊唑醇WDG 75% (w) Trifloxystrobin·tebuconazole WDG y=5.5671+0.9525x 0.254 0.222~0.291 0.9924 10% (w)苯醚甲环唑WDG 10% (w) Difenoconazole WDG y=5.9588+0.8439x 0.731 0.623~0.843 0.9884 250 g/L吡唑醚菌酯SC 250 g/L Pyraclostrobin SC y=5.6501+0.5764x 0.745 0.658~0.843 0.9959 227 g/L二氰蒽醌SC 227 g/L Dithianon SC y=4.2691+0.9029x 64.490 40.020~103.921 0.9408 500 g/L丙环唑ME 500 g/L Propiconazol ME y=4.3835+0.9116x 47.453 40.574~55.499 0.9912 250 g/L溴菌腈EC 250 g/L Bromothalonil EC y=4.8736+0.4457x 19.218 17.062~21.648 0.9948 10 g/L蛇床子素ME 10 g/L Osthole ME y=3.5437+1.0864x 2.191 1.195~4.0156 0.9656 10 g/L申嗪霉素SC 10 g/L Phenazine-1-carboxylic acid SC y=3.5916+1.1421x 1.711 1.418~2.063 0.9956 80 g/L宁南霉素AS 80 g/L Ningnanmycin AS y=3.1609+1.2538x 2.929 1.659~5.171 0.9715 3% (w)中生菌素WP 3%(w) Zhongshengmycin WP y=3.6943+1.1698x 1.307 0.675~2.531 0.9586 20 g/L异硫氰酸烯丙酯 EW 200 g/L Allylisothiocyanate EW y=3.8994+1.9167x 3.175 3.499~4.022 0.9981 200 g/L异硫氰酸烯丙酯SL 200 g/L Allylisothiocyanate SL y=4.8799+1.2955x 1.238 1.027~1.493 0.9906 1) x为药剂剂量取对数,y为死亡率转换成概率 1) x: Log value of the agent dose, y: Probability converted form mortality
下载: 导出CSV
表 5 苯醚甲环唑WDG和吡唑醚菌酯SC对Colletotrichum scovillei的联合毒力作用
Table 5 Co-toxicity of the mixed difenoconazole WDG and pyraclostrobin SC to Colletotrichum scovillei
V(吡唑醚菌酯):V(苯醚甲环锉) V(Difenoconazole): V(Pyraclostrobin) 毒力回归方程1)Regression equation of toxicity 相关系数 (R) Correlation coefficient EC50/ (mg·L−1) EC50 95%置信限/ (mg·L−1) 95% confidence limit of EC50 共毒系数 Co-toxicity coefficient 效果 Effect 4∶1 y=5.1605+0.8151x 0.9969 0.117 0.099~0.137 635.953 增效 Synergy 3∶2 y=5.6904+0.9141x 0.9964 0.176 0.154~0.201 420.795 增效 Synergy 1∶1 y=5.3989+1.7171x 0.9707 0.586 0.440~0.780 125.992 增效 Synergy 2∶3 y=5.2299+1.3827x 0.9892 0.682 0.566~0.822 108.012 相加 Additive effect 1∶4 y=5.1727+1.5551x 0.9895 0.774 0.624~0.962 95.836 相加 Additive effect 1) x为药剂剂量取对数,y为死亡率转换成概率 1) x: Log value of the agent dose, y: Probability converted form mortality
下载: 导出CSV
-
[1] 程曦, 杨潇, 董瑶. 2020年贵州辣椒产业产加销规模均位列全国第一[N/OL]. 多彩贵州网, 2021-09-16. http://news.gog.cn/system/2021/09/16/017981892.shtml. [2] 刁永朝, 蔡磊. 辣椒病原真菌多样性及其影响因素研究[C]//中国菌物学会. 中国菌物学会第七届全国会员代表大会暨2017年学术年会摘要集. 宜昌: 中国菌物学会, 2017: 230. [3] 李小霞, 肖仲久. 贵州省辣椒炭疽病病原菌鉴定及室内毒力测定[J]. 广东农业科学, 2011, 38(17): 55-57. doi: 10.3969/j.issn.1004-874X.2011.17.020 [4] 王妮, 尹显慧, 彭丽娟, 等. 辣椒炭疽病病原鉴定及其杀菌剂毒力测定[J]. 植物保护, 2019, 45(4): 216-223. [5] 杨佳文, 赵尊练, 张管曲, 等. 陕西线辣椒炭疽病原菌的鉴定及生物学特性研究[J]. 西北农业学报, 2017, 26(11): 1695-1705. doi: 10.7606/j.issn.1004-1389.2017.11.017 [6] LIU F L, TANG G T, ZHENG X J, et al. Molecular and phenotypic characterization of Colletotrichum species associated with anthracnose disease in peppers from Sichuan Province, China[J]. Scientific Reports, 2016, 6(1): 32761. doi: 10.1038/srep32761
[7] 魏立娟. 辣椒炭疽病菌的鉴定、综合防治及互作后辣椒基因差异表达[D]. 兰州: 甘肃农业大学, 2019. [8] 周建波, 殷辉, 吕红, 等. 8种不同类型药剂对辣椒炭疽病的田间防治效果[J]. 中国瓜菜, 2020, 33(11): 72-76. doi: 10.3969/j.issn.1673-2871.2020.11.016 [9] 吕宁, 周光海, 陈云, 等. 滴施生物药剂对棉花生长、黄萎病防治及土壤微生物数量的影响[J]. 西北农业学报, 2018, 27(7): 1056-1064. doi: 10.7606/j.issn.1004-1389.2018.07.018 [10] 蒋细良, 王劲波, 王慧敏, 等. 中生菌素对水稻悬浮细胞过氧化物酶基因转录表达的影响[J]. 中国生物防治, 2006, 22(3): 207-210. [11] 方中达. 植病研究方法[M]. 3版. 北京: 中国农业出版社, 1998. [12] CHOI Y W, HYDE K D, HO W H. Single spore isolation of fungi[J]. Fungal Diversity, 1999, 3: 29-38.
[13] THAN P P, JEEWON R, HYDE K D, et al. Characterization and pathogenicity of Colletotrichum species associated with anthracnose on chili (Capsicum spp. ) in Thailand[J]. Plant Pathology, 2008, 57: 562-572. doi: 10.1111/j.1365-3059.2007.01782.x
[14] 刘方玲. 四川辣椒炭疽病的病原学研究[D]. 成都: 四川农业大学, 2015. [15] WHITE T J, BRUNS T, LEE S, et al. PCR Protocols: A Guide to Methods and Applications [M]. San Diego: Academic Press, 1990: 315-322.
[16] CARBONE I, KOHN L M. A method for designing primer sets for speciation studies in filamentous ascomycetes[J]. Mycologia, 1999, 91(3): 553-556. doi: 10.1080/00275514.1999.12061051
[17] GUERBER J C, LIU B, CORRELL J C, et al. Characterization of diversity in Colletotrichum acutatum sensulato by sequence analysis of two gene introns, mtDNA and intron RFLPs, and mating compatibility[J]. Mycologia, 2003, 95(5): 872-895. doi: 10.1080/15572536.2004.11833047
[18] DAMM U, CANNON P F, WOUNDENBERG J H C, et al. The Colletotrichum acutatum species complex[J]. Studies in Mycology, 2012, 73(1): 37-113.
[19] TAMURA K, DUDLEY J, NEI M, et al. MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0[J]. Molecular Biology and Evolution, 2007, 24(8): 1596-1599. doi: 10.1093/molbev/msm092
[20] 黄清臻, 崔安义, 徐之明, 等. 共毒系数的简易计算及其应用[J]. 医学动物防制, 1993, 9(2): 86-87. [21] TOPOREK S M, KEINATH A P. First report of Colletotrichum scovillei causing anthracnose fruit rot on pepper in South Carolina, United States[J]. Plant Disease, 2020, 105(4): 1222.
[22] OO M M, LIM G, JANG H A, et al. Characterization and pathogenicity of new record of anthracnose on various chili varieties caused by Colletotrichum scovillei in Korea[J]. Mycobiology, 2017, 45(3): 184-191. doi: 10.5941/MYCO.2017.45.3.184
[23] ZHAO W, WANG T, CHEN Q Q, et al. First report of Colletotrichum scovillei causing anthracnose fruit rot on pepper in Anhui Province, China[J]. Plant Disease, 2016, 100(10): 2168.
[24] CAIRES N P, PINHO D B, SOUZA J S C, et al. First report of anthracnose on pepper fruit caused by Colletotrichum scovillei in Brazil[J]. Plant Disease, 2014, 98(10): 1437.
[25] KANTO T, UEMATSU S, TSUKAMOTO T, et al. Anthracnose of sweet pepper caused by Colletotrichum scovillei in Japan[J]. Journal of General Plant Pathology, 2014, 80(1): 73-78. doi: 10.1007/s10327-013-0496-9
[26] LI Q L, BU J Y, SHU J Y, et al. Colletotrichum species associated with mango in Southern China[J]. Scientific Reports, 2019, 9(1): 18891. doi: 10.1038/s41598-019-54809-4
[27] 王薇. 苹果炭疽叶枯病病原学及苹果炭疽病侵染来源研究[D]. 杨凌: 西北农林科技大学, 2017. [28] 任璐, 周建波, 刘慧平, 等. 辣椒炭疽病菌Colletotrichum gloeosporioides对啶氧菌酯的敏感基线及抗性突变体生物学性状[J]. 植物保护, 2017, 43(6): 29-37. doi: 10.3969/j.issn.0529-1542.2017.06.005 [29] 石玉星, 曹俊宇, 任璐, 等. 辣椒炭疽病菌抗啶氧菌酯突变体渗透压敏感性及交互抗性[J]. 山西农业科学, 2017, 45(6): 1002-1005. doi: 10.3969/j.issn.1002-2481.2017.06.34 [30] 陈聃, 时浩杰, 吴慧明, 等. 浙江省葡萄炭疽菌对甲基硫菌灵和戊唑醇的抗药性研究[J]. 果树学报, 2013, 30(4): 665-668.