Establishment of IMSA-LAMP detection method for Ralstonia solanacearu
-
摘要:目的
桑树青枯病是由茄科雷尔氏菌Ralstonia solanacearum引起的一种危害严重的细菌性病害,建立一种快速、灵敏的茄科雷尔氏菌检测方法,对桑树青枯病的有效控制有重要意义。
方法本研究以茄科雷尔氏菌果胶裂解酶基因(Pectate lyase gene)为靶标,基于等温多自配引发扩增技术(Isothermal multiple self-matching-initiated amplification,IMSA)的引物设计原理,结合环介导等温扩增技术(Loop-mediated isothermal amplification,LAMP)的反应体系,建立一种快速有效检测茄科雷尔氏菌的IMSA-LAMP法,并对该方法的最佳反应参数进行了筛选。
结果基于果胶裂解酶基因建立的IMSA-LAMP检测方法在64.5 ℃条件下,45 min内可完成对阳性样品的特异检测,对茄科雷尔氏菌的模板DNA检测灵敏度达200 fg/µL (对应菌为1×102 CFU/mL);对生产上收集的疑似桑树青枯病病样的检出率为87.5%。
结论IMSA-LAMP检测方法具有良好的实用性,可为桑树青枯病的快速检测、诊断与防疫提供新的技术支持。
Abstract:ObjectiveMulberry bacterial wilt is a seriously harmful bacterial disease caused by Ralstonia solanacearum. Therefore, it is of great significance to establish a rapid and sensitive detection method for R. solanacearum to effectively control mulberry bacterial wilt.
MethodPectate lyase gene of R. solanacearum was used as the target. Based on the primer design principle of isothermal multiple self-matching-initiated amplification (IMSA), combining the loop-mediated isothermal amplification (LAMP) reaction system, a rapid and effective IMSA-LAMP method for detection ofR. solanacearum was established. The optimal reaction parameters of this method were screened.
ResultThe IMSA-LAMP method based on pectate lyase gene could complete the specific detection of positive samples within 45 min at 64.5 ℃, and the detection sensitivity of R. solanacearum template DNA was 200 fg/μL (the corresponding bacteria detection sensitivity was 1 × 102 CFU/mL); The detection rate of suspected mulberry bacterial wilt samples collected in production was 87.5%.
ConclusionThis method has good practicability and can provide new technical support for the rapid detection, diagnosis and epidemic prevention of mulberry bacterial wilt.
-
Keywords:
- Mulberry bacterial wilt /
- IMSA-LAMP /
- Ralstonia solanacearum /
- Pectate lyase gene
-
钾是农作物生长必需的三大营养元素之一[1].我国呈现出不同程度的土壤缺钾现状, 且南方较北方的缺钾情况严重, 如广东的水稻土壤、旱地土壤缺钾面积均达到了90%以上[2].同时, 可利用的水溶性钾矿资源短缺, 供给量仅占需求量的30% ~ 40%, 进口已成为钾肥的主要来源[3].我国的非水溶性钾矿资源却非常丰富, 达到了100亿t[4].充分开发利用这部分资源, 将极大缓解我国的缺钾现状.我国对非水溶性钾矿已做了较多的研究[5-6], 然而大部分的提钾工艺复杂, 成本高, 难以推广.近年来, 基于活化概念的理化促释技术提供了一条新型研发思路[7].已有的研究结果表明, 活化后钾长石的钾释放量显著提高, 可部分替代氯化钾而肥效不减[8-9].开展活化钾矿的钾素释放机理与规律的理论研究, 尤其是对动力学规律的研究, 对其肥力评价指标的建立具有重要意义.
钾的释放既受含钾矿物类型的影响, 也受钾释放的溶液环境(如各种离子种类和浓度)的影响[10].范钦桢[11]发现铵态氮肥中的NH4+会抑制土壤中非交换态钾和结构钾的释放, 土壤中常见的阳离子有NH4+、Na+、Ca2+等, 而这些阳离子同样会影响非水溶性钾矿的释放.王瑾[12]研究了不同阳离子盐溶液对黑云母、白云母、正长石等非水溶性钾矿钾释放的影响, 发现随着离子种类、钾矿类型的不同, 钾的释放也呈现不同的释放特征.本研究选取2种钾矿及其QN活化钾矿为研究材料, 采用NH4Cl和NaCl这2种阳离子盐溶液做浸提剂对其连续振荡提取, 建立活化钾矿的钾释放动力学模型, 旨在从动力学角度研究活化钾矿的高效释放特征, 为评价活化钾矿的植物有效性提供理论依据.
1. 材料与方法
1.1 材料
供试的非水溶性钾矿为钾长石和富钾页岩, 其中, 钾长石w (K2O)为8.57%, 取自广东五华; 富钾页岩w(K2O)为11.42%, 取自河北张家口.2种钾矿经风干、磨细后过100目筛备用.
活化钾矿的制备方法:分别称取上述钾矿20 g, 加入质量分数为5%的QN活化剂, 再加入2 mL蒸馏水, 混合研磨5 min, 风干、磨细, 过100目筛备用.其中, QN活化剂为含Na+、不含K+的无机活化剂.
浸提剂分别为10 mmol·L-1的NH4Cl和NaCl.
1.2 方法
准确称取钾矿及活化钾矿0.500 0 g于离心管中, 分别加入50 mL不同的浸提剂溶液, 对照加入去离子水(H2O), 摇匀, 在振荡机上振荡15 min, 取出后5 000 r·min-1离心.倒出全部上清液, 用火焰光度法测定溶液钾的含量.残渣中分别加入50 mL上述溶液, 重复浸提步骤, 钾长石、活化钾长石连续提取10次, 页岩、活化页岩连续提取15次.每个处理设3个重复.上述提取次数均根据实际浸提过程中到达平衡附近的时间确定.
1.3 释放动力学模型
一级动力学模型:y = a-ae-bx,
双常数模型:y = axb,
扩散模型:y = a+ bx0.5,
Elovich模型:y = a+ blnx.
上述模型中, x为浸提时间, y为钾矿的累积释钾量, a、b为模型常数[13-16].
1.4 数据处理方法
数据的处理、分析和制图分别采用Excel、Spass13.0、Matlab7.1等软件.
2. 结果与分析
2.1 连续振荡条件下活化钾矿的钾释放
如图 1所示, 各浸提剂浸提钾矿的释放均表现为前期快速, 之后缓慢释放的变化趋势, 其中NH4Cl浸提时, 钾矿及活化钾矿在30 min左右即完成了快速释放, 进入了缓慢释放阶段.NH4Cl和NaCl在浸提钾长石、活化钾长石时, 均在45 min左右达到了缓慢释放阶段; H2O在浸提富钾页岩、活化页岩时, 快速释放阶段为0 ~ 90 min, 90 min之后为缓慢释放阶段, 而NaCl浸提到45 min左右时, 富钾页岩、活化页岩即进入了缓慢释放阶段.
![图 1 不同浸提剂下活化钾矿的钾释放变化]() 图 1 不同浸提剂下活化钾矿的钾释放变化Figure 1. Variations of K releases of activated insoluble potassium ores with different cation solutions
图 1 不同浸提剂下活化钾矿的钾释放变化Figure 1. Variations of K releases of activated insoluble potassium ores with different cation solutions连续浸提的过程中, 活化钾矿与钾矿表现出一致的浸提规律.初始阶段, 钾长石、活化钾长石的钾释放量表现为:NH4Cl>NaCl>H2O, 随着浸提时间的延长, 释钾量逐渐减少, 到达释钾平衡附近时, 3种浸提剂的累积释钾量表现为:NaCl>NH4Cl≈H2O; 富钾页岩、活化页岩的整个动态释钾过程均表现为: H2O>NaCl>NH4Cl.
对钾矿及其活化钾矿的累积释钾量分析可知, 在H2O、NH4Cl、NaCl浸提下, 活化钾长石的累积释钾量分别是钾长石的2.3、2.0和1.7倍, 活化页岩的累积释钾量分别是富钾页岩的2.5、3.3和2.5倍, 所以活化钾矿的累积释钾能力大于钾矿.由图 1还可以看出, 富钾页岩的累积释钾能力大于钾长石、活化页岩的累积释钾能力大于活化钾长石.
2.2 钾矿的释钾动力学模型
由图 1中钾矿释钾的动态数据, 建立不同浸提剂钾矿的钾释放动力学模型, 拟合结果如表 1所示.其中, 模型拟合的优劣取决于拟合性, 即计算值与实测值的符合程度, 常用相关系数(R)和标准差(S)来评定, R越大、S越小拟合性越好.由表 1可以看出, 除钾长石的一级动力学拟合方程R达显著水平外, 其余拟合方程的R均达到极显著水平, 相关系数在0.698 6 ~ 0.997 3之间.累积释钾量的计算值与实测值之间的S在11.35 ~ 295.90之间, 表明4个模型均能很好的拟合钾矿及活化钾矿的动态释钾过程.
表 1 连续振荡条件下活化钾矿释钾的动力学模型1)Table 1. The kinetics model of K release of activated potassium ores with successive extraction
双常数模型、一级动力学模型、扩散模型和Elovich模型拟合钾长石的累积释钾量, 拟合R的平均值分别为0.971 3、0.832 2、0.953 3和0.978 2;拟合S的平均值分别为15.48、22.99、19.49和13.30, 双常数模型和Elovich模型两者间的R和S基本没有差别, 所以, 钾长石的最优释放动力学模型是Elovich模型或双常数模型.
对于活化钾长石, 4种模型拟合R的平均值分别为0.948 4、0.920 1、0.900 3和0.958 1;拟合S的平均值分别为22.35、26.28、31.07和20.50, 与钾长石的结果类似, 活化钾长石的最优释放动力学模型亦为Elovich模型或双常数模型.
对页岩, 4种模型拟合R的平均值分别为0.979 4、0.906 4、0.974 6和0.981 7;S的平均值分别为:64.68、88.84、72.36和51.36, 钾的释放动力学模型拟合性表现为:Elovich模型>双常数模型>扩散模型>一级动力学模型.所以, 页岩的最优释放动力学模型为Elovich模型.
对活化页岩, 4种模型拟合R的平均值分别为0.974 3、0.849 7、0.956 2和0.981 6;S的平均值分别为126.70、194.30、169.11和95.79, 动力学方程拟合性:Elovich模型>双常数模型>扩散模型>一级动力学模型.所以, 活化页岩的最优释放动力学模型是Elovich模型.
4种模型拟合时, H2O、NH4Cl、NaCl浸提钾长石的方程S均表现为:NaCl>H2O>NH4Cl, 活化钾长石亦表现出相同规律.一级动力学模型拟合时, 3种浸提剂之间的S相差不大, 说明除了一级动力学模型, 其他3种动力学模型拟合钾长石、活化钾长石时, NH4Cl浸提下的模型拟合性均优于NaCl.4种模型拟合页岩时浸提剂间的S均表现出:H2O>NaCl>NH4Cl, 浸提活化页岩时亦表现出相同规律, 说明4种模型拟合下, NH4Cl浸提页岩、活化页岩的拟合性均优于NaCl.
2.3 动力学模型参数与钾矿释钾关系
由上述分析可知, 双常数模型、一级动力学模型、扩散模型和Elovich模型均具有较好的拟合性, 模型中的参数对于活化钾矿中钾素在盐溶液持续作用下的释放特征具有重要的意义.
拟合方程的参数见表 2.双常数方程的参数a表示释放过程的初始瞬时速率[17-18], a值越大, 钾矿释钾的初始瞬时速率越大.通过比较双常数方程的a值可以看出, 活化钾矿的钾初始释放速率显著高于未活化钾矿, 在H2O、NH4Cl和NaCl的浸提条件下, 活化钾长石的钾初始释放速率分别比钾长石增加了3.7、1.1和1.6倍; 活化页岩的钾初始释放速率分别比富钾页岩增加了4.7、6.0和4.0倍.
表 2 活化钾矿的动力学模型拟合参数Table 2. The kinetics model fitting parameters of activated potassium ores
对Elovich方程求导, 可得到Elovich速率方程: y = b/x, 由b值可以求出任一时间的释放速率[17, 19-20].b值越大, 钾矿释钾的速率越大.表 2可以看出, NH4Cl浸提钾矿及活化钾矿的b显著小于其他浸提剂, 表明NH4Cl浸提的钾释放速率远小于其他浸提剂.比较钾长石和活化钾长石的b发现, 除了NaCl浸提活化钾长石的b小于钾长石, H2O和NH4Cl浸提活化钾长石的b分别比钾长石增加了10.3%和71.2%.H2O、NH4Cl和NaCl浸提活化页岩的b分别比页岩增加了61.7%、102.4%和86.3%.
一级动力学方程的a值表示释放过程的最大平衡释放量[16, 22-23], 由表 2可以看出, H2O、NH4Cl和NaCl连续浸提活化钾长石钾的一级动力学方程的a分别比钾长石增加了139.6%、99.6%和73.3%;活化页岩钾的a分别比富钾页岩增加了1.3、2.4和1.6倍.
动力学参数的分析可以看出, 活化后钾矿中钾的初始释放速率、平均释放速率以及最大平衡释放量均有所提高.其中, 初始释放速率增大是活化钾矿中钾的最大平衡释放量增多的主要原因.
3. 讨论与结论
各浸提剂下, 钾矿的累积释钾能力及动力学参数均表现出了富钾页岩大于钾长石、活化页岩大于活化钾长石, 说明矿物类型不同, 其释钾能力差异较大.这可能与矿物结构有关, 钾长石矿物结构为无水架状结构铝硅酸盐矿物, 钾原子的位置位于晶格内部, 钾原子落在10个氧原子所组成的穴中, 与6个氧原子相距0.285 nm, 因此阻碍了钾的释放[10].页岩结构较为复杂, 如辽宁省朝阳地区的页岩包含了多种以独立矿物形式出现的含钾矿物, 主要成分是钾长石, 约80%的钾赋存于钾长石中, 其次为白云母、伊利石等, 约20%的钾赋存于云母类矿物中[24], 而长石类与云母类相比, 由于钾离子处在相邻四面体的空隙中, 释钾较为困难[15].
本研究结果表明, 钾长石、活化钾长石的钾初始释放量表现为:NH4Cl>NaCl>H2O, 累积释钾量表现为:NaCl>NH4Cl≈H2O.一开始, NH4+的提取能力强于Na+, 之后提取能力减弱, 这可能是因为与Na+、Ca2+等水化半径较大的离子相比, NH4+与K+有几乎相同的离子半径和水化能[25-26], 甚至在电性、化合价、释放与固定机制等多方面都具有相似之处[27], 故NH4+更容易置换矿物表面及边缘、楔形位点吸附的钾, 当矿物的速效钾含量较高时, 交换能力最强[28].随着浸提时间的延长, 矿物表面和边缘处吸附的钾较少, 水化半径较大的Na+提钾能力大于NH4+, 这是因为Na+虽然因其水化半径较大不易置换楔形位点上吸附的K+, 但能置换一部分矿物晶层表面吸附的K+ [29-30]; Na+、Ca2+等水化半径较大的盐离子可以撬开矿物晶层, 使得一部分易释放的非交换性钾释放出来[28].本试验中, H2O浸提下页岩、活化页岩的累积释钾量大于2种盐溶液, 这可能是因为页岩复杂的结构中存在大量的层状结构, K+通过直接的扩散而不需要离子交换就可以释放出来.
通过动力学模型来拟合钾矿释钾的过程并描述其释钾规律鲜见报道.王瑾等[15]研究发现用不同有机酸连续浸提黑云母、正长石等钾矿, 其释放的最优动力学模型为双常数模型或Elovich模型.为了准确、定量地描述某种元素的动态释放过程, 有必要建立或引用各种数学模型, 吕晓男等[30]通过多个动力学模型拟合电超滤方法下土壤钾释放的动态过程发现, Elovich方程的参数b和双常数方程的参数a与土壤速效钾和大麦相对产量之间存在显著或极显著相关.本试验建立了活化钾矿的释放动力学模型, 同时对动力学模型的参数进行了比较, 发现活化后钾矿的钾初始释放速率、平均释放速率、最大平衡释放量均显著增大, 并从动力学角度定量评价了活化后钾矿的释钾效果.至于参数能否作为评价其生物有效性的指标还需进一步的研究.
本试验得到如下结论:1)浸提剂浸提活化钾长石的钾初始释放量表现为:NH4Cl>NaCl>H2O; 累积释放量表现为:NaCl>NH4Cl≈H2O; 浸提活化页岩的整个动态释钾过程均表现为:H2O>NaCl>NH4Cl.在盐溶液的连续振荡浸提下, 活化钾矿的累积释钾能力大于钾矿, 富钾页岩及活化页岩的累积释钾能力大于相同处理下的钾长石.2)钾长石、活化钾长石钾的最优释放动力学模型是Elovich模型或双常数模型; 页岩、活化页岩的最优动力学模型是Elovich模型.除了一级动力学模型拟合钾长石、活化钾长石时, NH4Cl、NaCl浸提剂间拟合性无差异外, 其他情况时, NH4Cl浸提下的模型拟合性均优于NaCl.3)通过动力学模型参数比较, 从动力学角度定量评价了钾矿的QN活化效果, 结果表明, 活化后钾矿的钾初始释放速率、平均释放速率以及最大平衡释放量均有显著提高.
-
![]()
![]()
图 2 IMSA-LAMP 6组引物对茄科雷尔氏菌的检测结果
Figure 2. Detection results of Ralstonia solanacearum under six primer groups of IMSA-LAMP
![]()
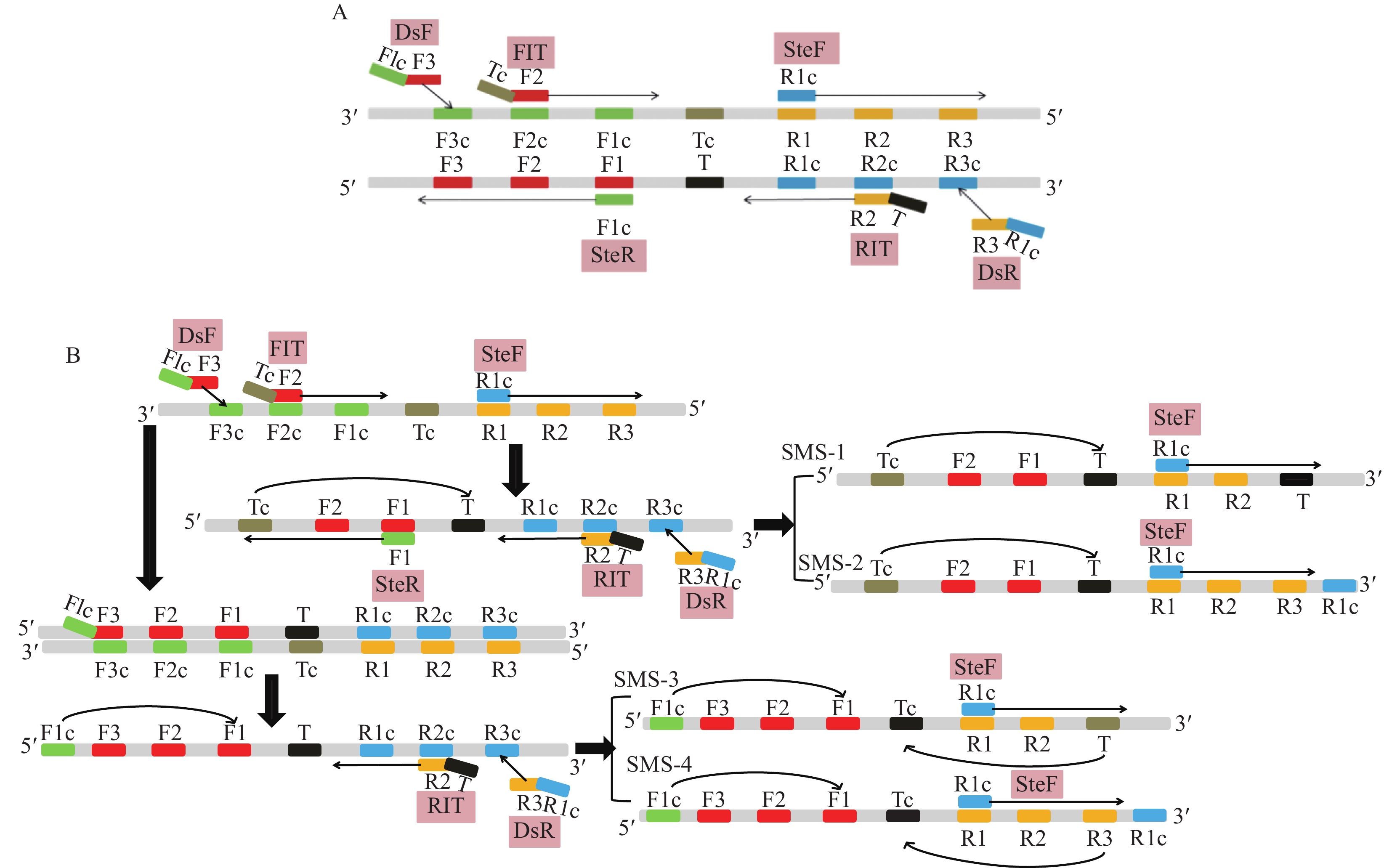
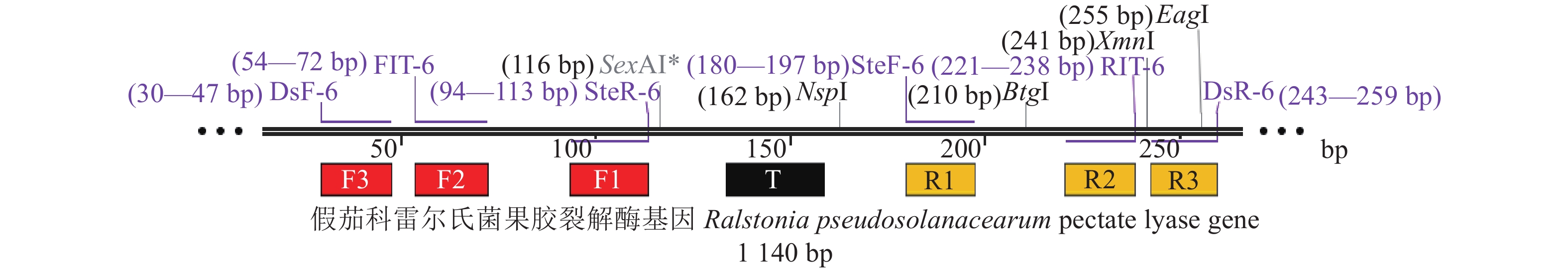
图 3 IMSA-LAMP引物组6的6条检测引物的位置及7个特异识别位点
F1:CAGCGCGCAGTACAACTGCT,F2:GAATCACGATGCGGGTTCC, F3:CCATTTCCAGACGCCCTC,T:TCAAAGCGAGCAAGCTGTTCGGGAA,R1:CGACAACGCCGGCCAGAA,R2:GTCATCATGGCGGCTCGA,R3:GTTCGGTGCAAACGGCC
Figure 3. Locations of six primers of primer group 6 and their seven specific recognition sites in IMSA-LAMP
![]()
图 4 茄科雷尔氏菌IMSA-LAMP特异性检测结果
1~6:茄科雷尔氏菌(YZqk1、YLqk1、XZqk2、XCqk4、LZqk5、YDqk6);7:阴沟肠杆菌(XCYG-001);8:克雷伯氏菌(LCKL-001);9:菠萝泛菌(LCFJ-001);10:铜绿假单胞菌(YD-001);11:丁香假单胞菌(ZJDX-003);12:多黏类芽孢杆菌(YD-002);13:短小杆菌(YDDX-001);14:芽孢杆菌(YD-003);15:无菌水;16、17:健康桑树DNA
Figure 4. Specific detection results of IMSA-LAMP for Ralstonia solanacearum
1−6: Ralstonia solanacearum (YZqk1, YLqk1, XZqk2, XCqk4, LZqk5, YDqk6); 7: Enterobacter cloacae (XCYG-001); 8: Klebsiella sp. (LCKL-001); 9: Pantoea ananas (LCFJ-001); 10: Pseudomonas aeruginosa (YD-001); 11: Pseudomonas syringae (ZJDX-003); 12: Paenibacillus polymyxa (YD-002); 13: Bacillus pumilus (YDDX-001); 14: Bacillus sp. (YD-003); 15: Sterile water; 16−17: DNA of healthy mulberry
![]()
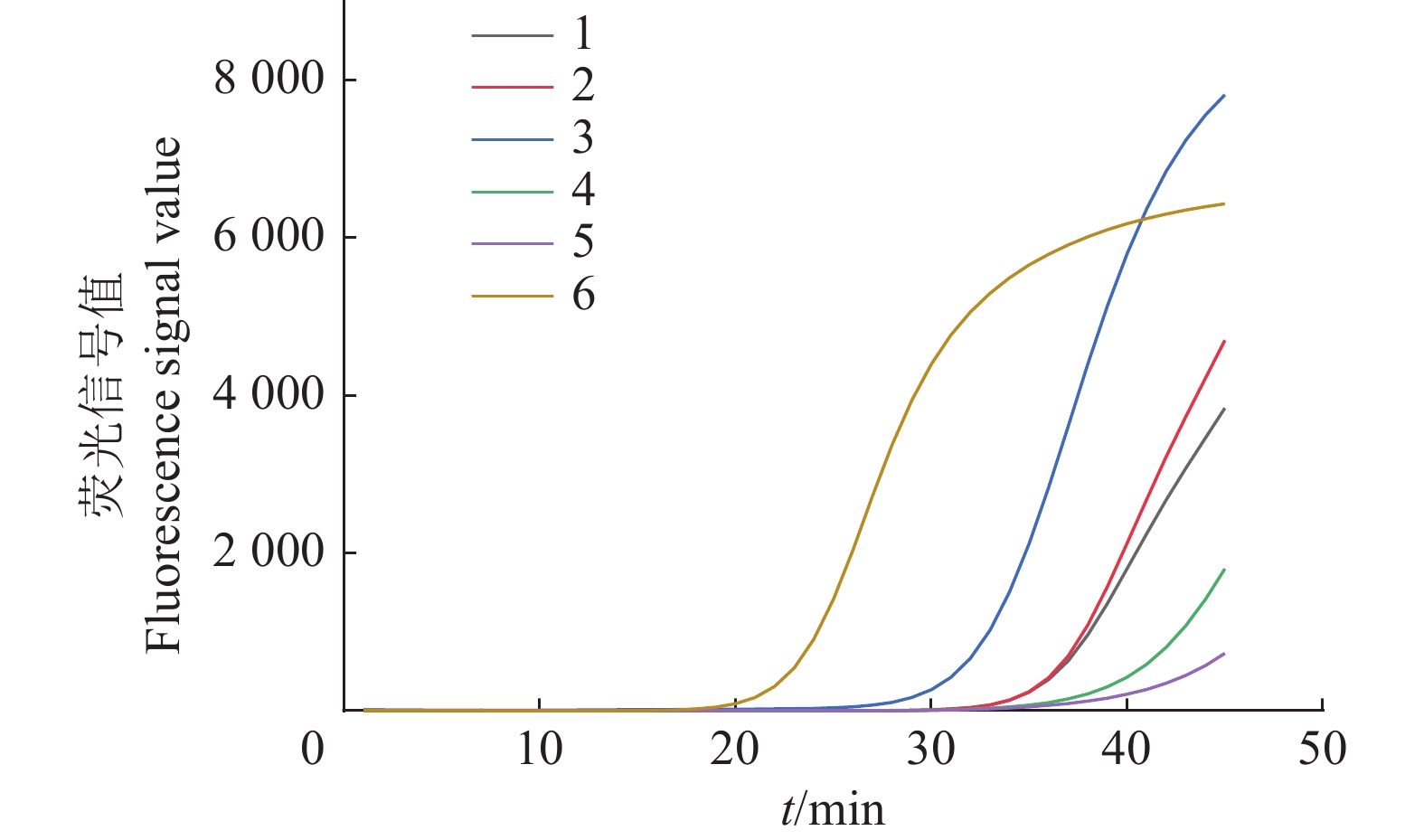
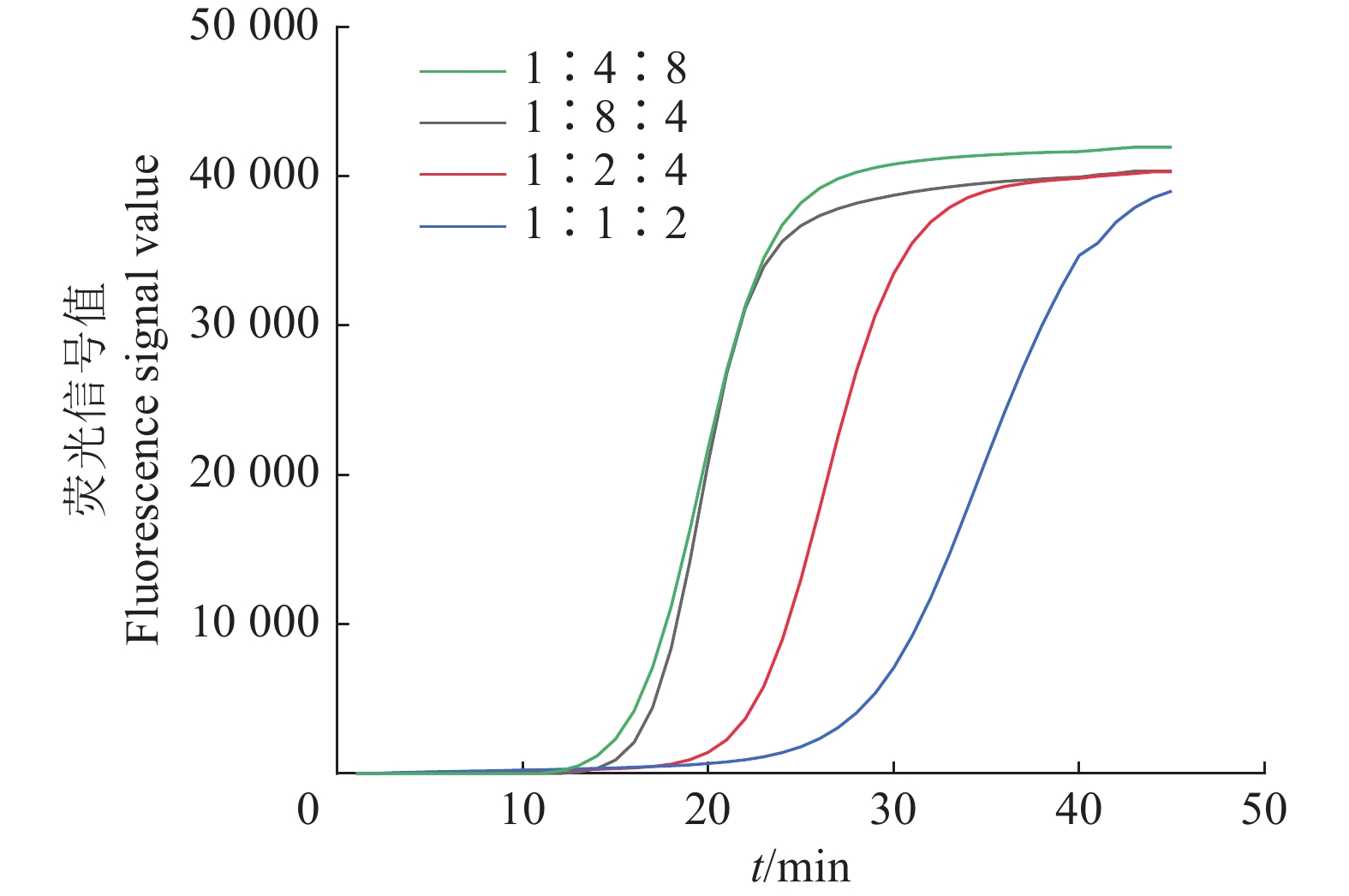
图 5 不同外、茎、内引物浓度比例下茄科雷尔氏菌IMSA-LAMP检测结果
Figure 5. IMSA-LAMP detection results ofRalstonia solanacearum under different concentration ratios of outer, stem and inner primers
![]()
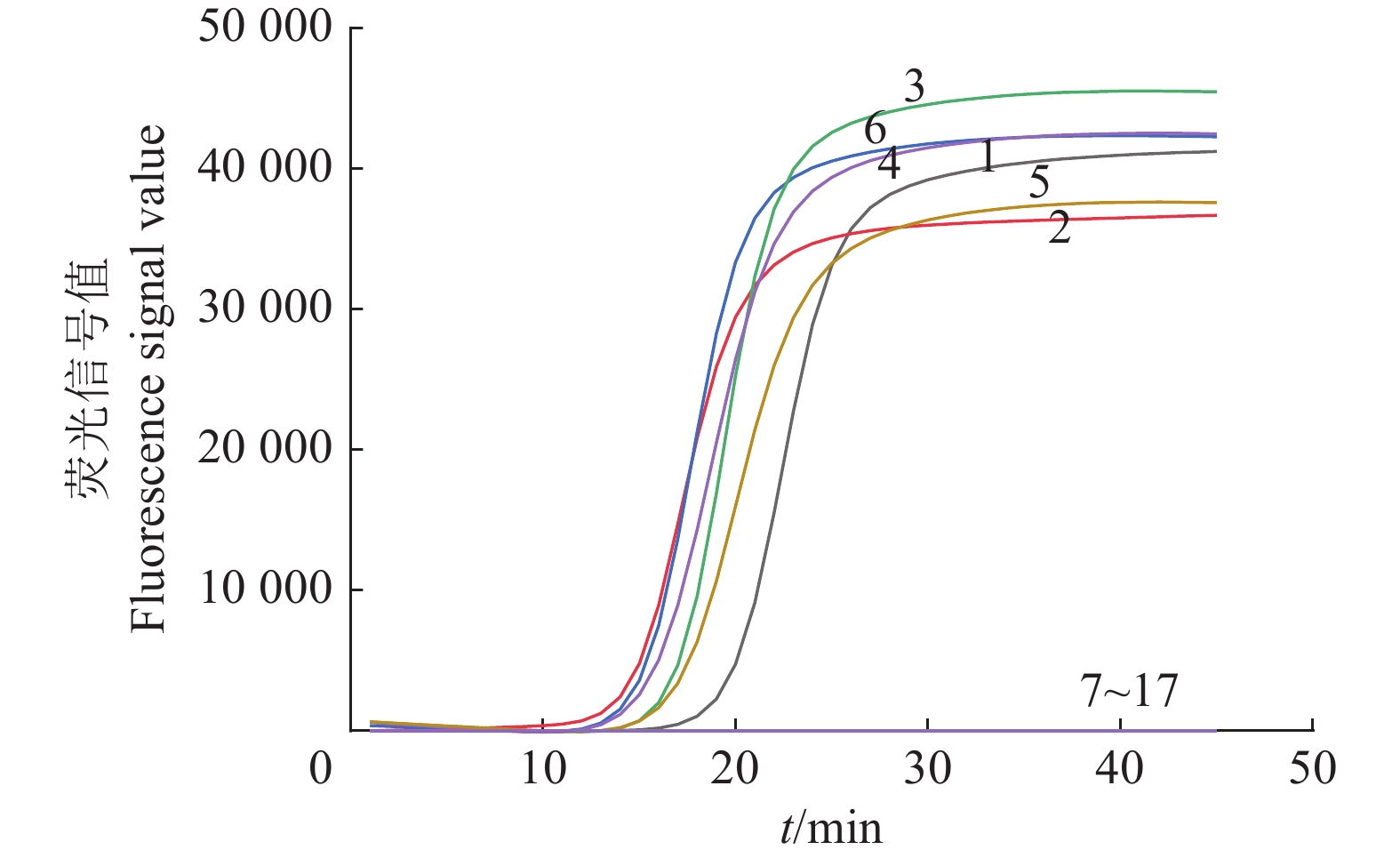
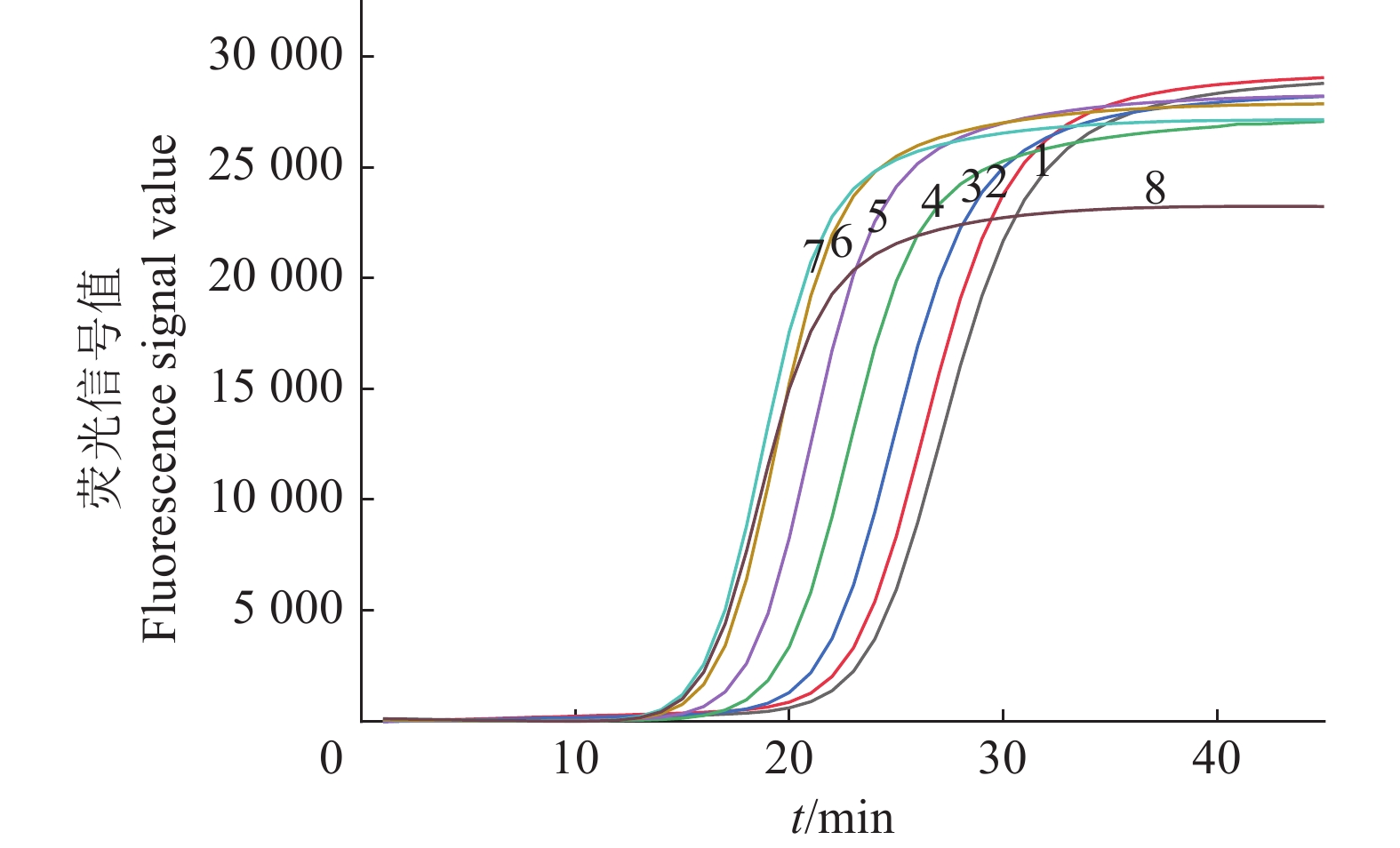
图 6 不同反应温度茄科雷尔氏菌IMSA-LAMP检测结果
1:61.5 ℃,2:62.0 ℃,3:62.5 ℃,4:63.0 ℃,5:63.5 ℃,6:64.0 ℃,7:64.5 ℃,8:65.0 ℃
Figure 6. Detection results of IMSA-LAMP for Ralstonia solanacearum at different reaction temperatures
![]()
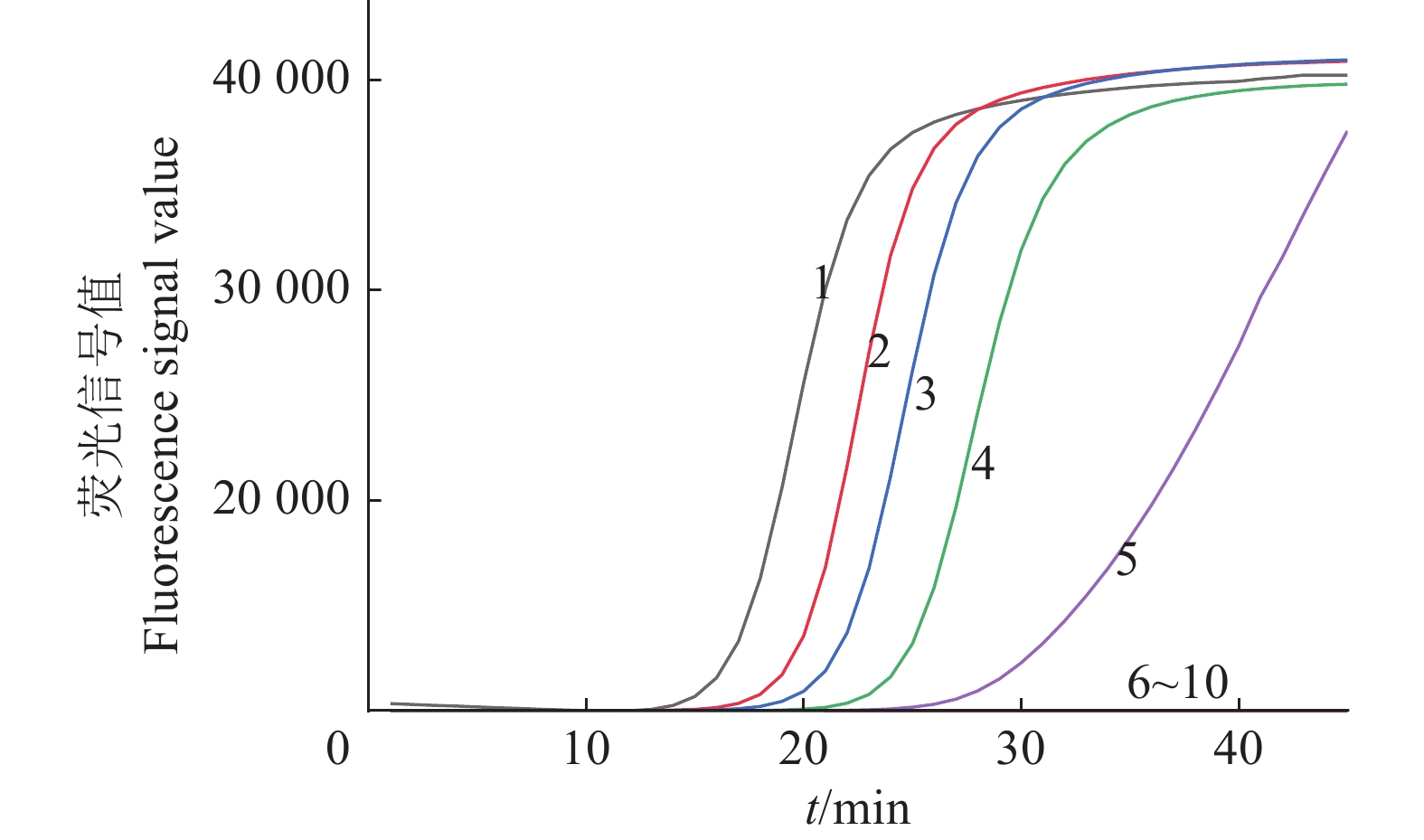
图 7 IMSA-LAMP对茄科雷尔氏菌灵敏度的检测结果
1:2 ng/μL (1×106 CFU/mL);2:200 pg/μL (1×105 CFU/mL);3:20 pg/μL (1×104 CFU/mL);4:2 pg/μL (1×103 CFU/mL);5:200 fg/μL (1×102 CFU/mL);6:20 fg/μL (1×10 CFU/mL);7:2 fg/μL (1×10−1 CFU/mL);8:200 ag/μL (1×10−2 CFU/mL);9:无菌水;10:健康桑树DNA
Figure 7. Sensitivity test results of IMSA-LAMP for Ralstonia solanacearum
1: 2 ng/μL (1×106 CFU/mL); 2: 200 pg/μL (1×105 CFU/mL); 3: 20 pg/μL (1×104 CFU/mL); 4: 2 pg/μL (1×103 CFU/mL); 5: 200 fg/μL (1×102 CFU/mL); 6: 20 fg/μL (1×10 CFU/mL); 7: 2 fg/μL (1×10−1 CFU/mL); 8: 200 ag/μL (1×10−2 CFU/mL); 9: Sterilized water; 10: DNA of healthy mulberry
![]()
图 8 PCR对茄科雷尔氏菌灵敏度检测结果
1:2 ng/μL (1×106 CFU/mL);2:200 pg/μL (1×105 CFU/mL);3:20 pg/μL (1×104 CFU/mL);4:2 pg/μL (1×103 CFU/mL);5:200 fg/μL (1×102 CFU/mL);6:20 fg/μL (1×10 CFU/mL);7:2 fg/μL (1×10−1 CFU/mL);8:200 ag/μL (1×10−2 CFU/mL);9:无菌水;10:健康桑树DNA
Figure 8. Sensitivity test results of PCR for Ralstonia solanacearum
1: 2 ng/μL (1×106 CFU/mL); 2: 200 pg/μL (1×105 CFU/mL); 3: 20 pg/μL (1×104 CFU/mL); 4: 2 pg/μL (1×103 CFU/mL); 5: 200 fg/μL (1×102 CFU/mL); 6: 20 fg/μL (1×10 CFU/mL); 7: 2 fg/μL (1×10−1 CFU/mL); 8: 200 ag/μL (1×10−2 CFU/mL); 9: Sterilized water; 10: DNA of healthy mulberry
![]()
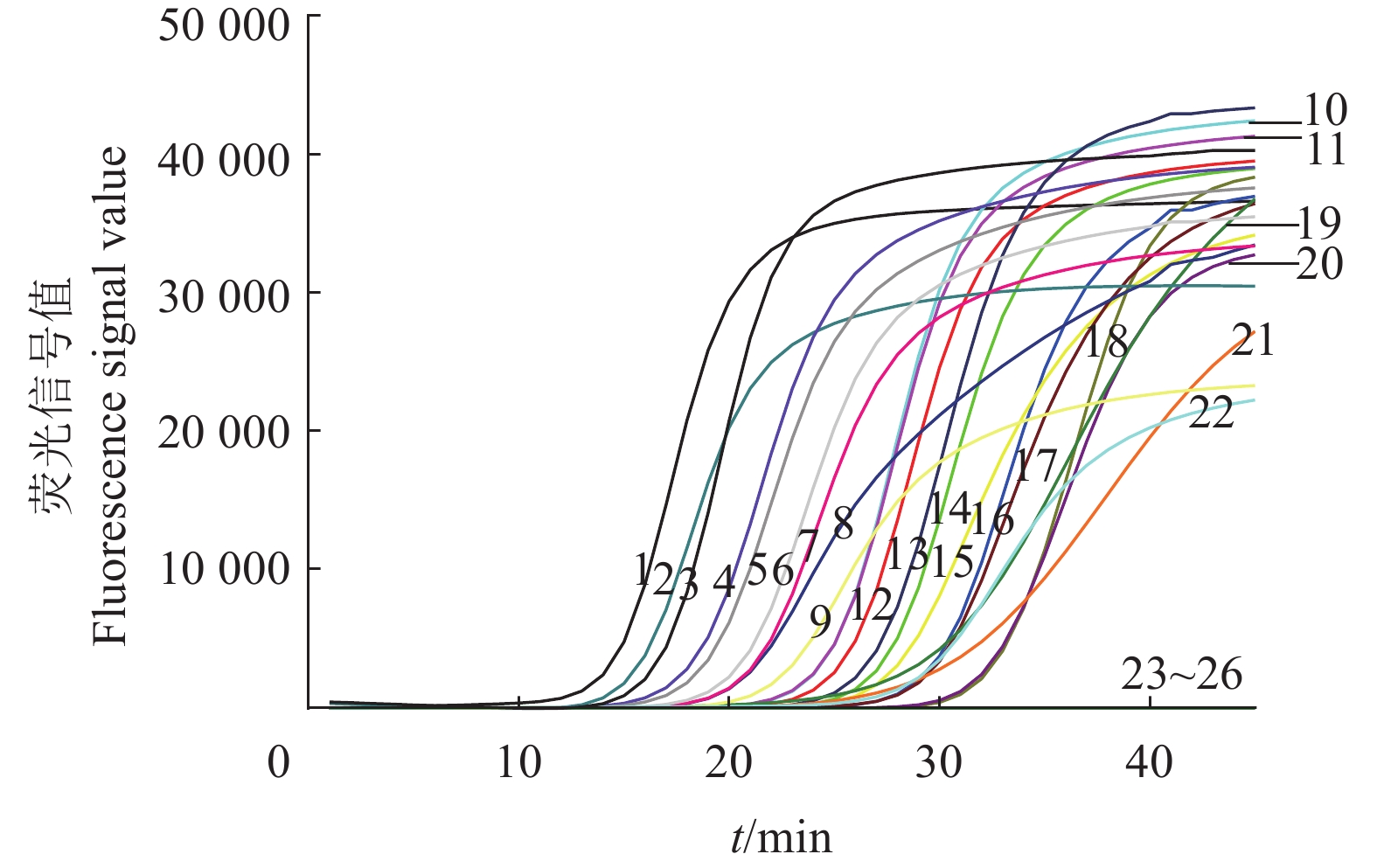
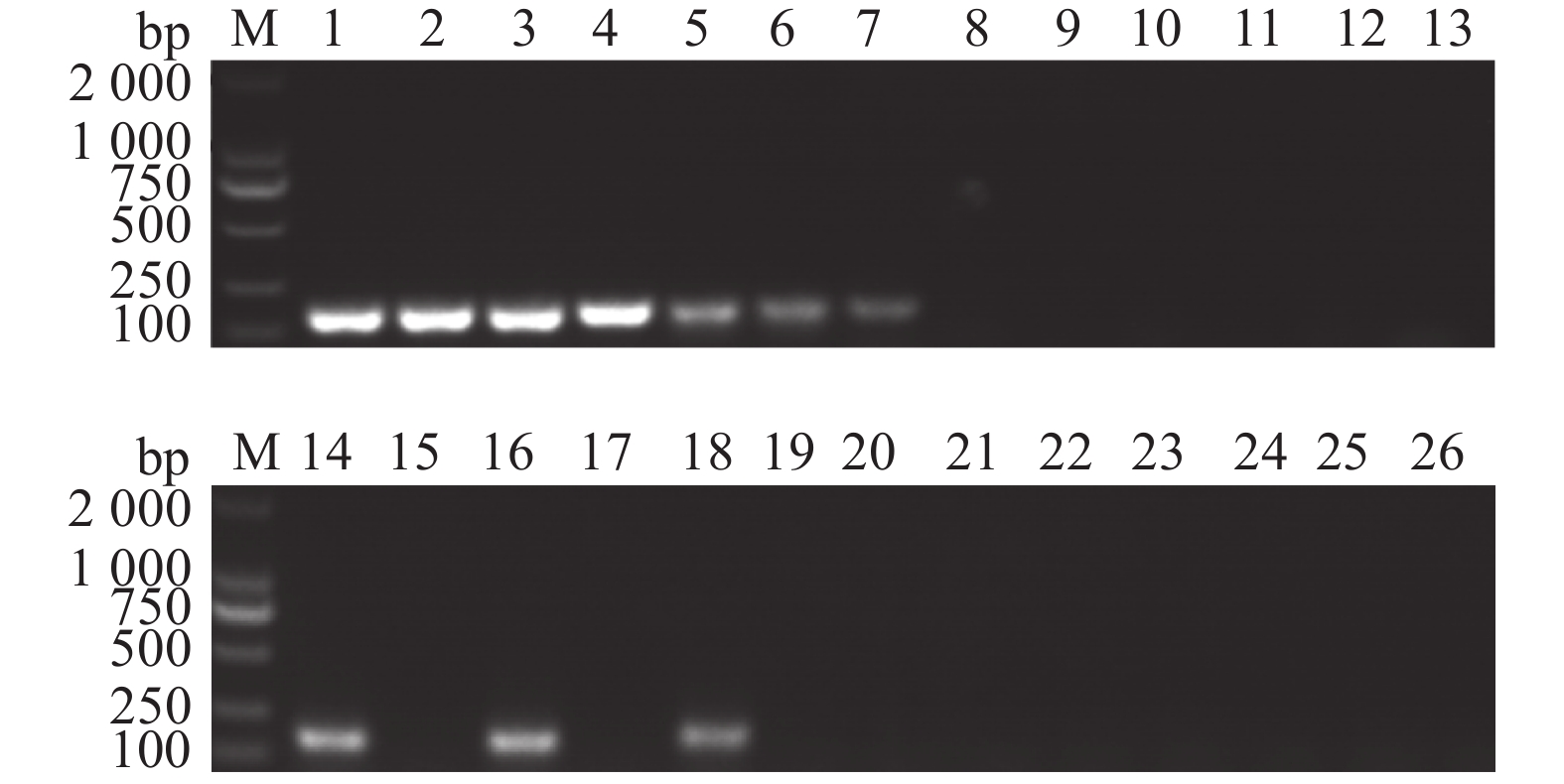
图 9 IMSA-LAMP对不同地区疑似桑树青枯病病样的检测结果
1:茄科雷尔氏菌YZqk1;2~5:广西来宾;6~9:广西宜州;10~12:广东英德;13~14:广东阳山;15~16:广东罗定;17~18:广东广州;19~21:广西都安;22~23:广西环江;24~25:广西忻城;26:健康桑树DNA
Figure 9. Detection results of suspected mulberry bacterial wilt samples from different districts by IMSA-LAMP
1: Ralstonia solanacearum YZqk1; 2–5: Laibin of Guangxi; 6–9: Yizhou of Guangxi; 10–12: Yingde of Guangdong; 13–14: Yangshan of Guangdong; 15–16: Luoding of Guangdong; 17–18: Guangzhou of Guangdong; 19–21: Du'an of Guangxi; 22–23: Huanjiang of Guangxi; 24–25: Xincheng of Guangxi; 26: Healthy mulberry DNA
![]()
图 10 PCR对不同地区疑似桑树青枯病病样的检测结果
1:茄科雷尔氏菌YZqk1;2~5:广西来宾;6~9:广西宜州;10~12:广东英德;13~14:广东阳山;15~16:广东罗定;17~18:广东广州;19~21:广西都安;22~23:广西环江;24~25:广西忻城;26:健康桑树DNA
Figure 10. Detection results of suspected mulberry bacterial wilt samples from different districts by PCR
1: Ralstonia solanacearum YZqk1; 2–5: Laibin of Guangxi; 6–9: Yizhou of Guangxi; 10–12: Yingde of Guangdong; 13–14: Yangshan of Guangdong; 15–16: Luoding of Guangdong; 17–18: Guangzhou of Guangdong; 19–21: Du'an of Guangxi; 22–23: Huanjiang of Guangxi; 24–25: Xincheng of Guangxi; 26: Healthy mulberry DNA
表 1 桑树青枯病田间病样
Table 1 Field samples of mulberry bacterial wilt
地区 District 坐标 Coordinate 品种 Cultivar 数量 Quantity 广东阳山 Yangshan, Guangdong E112.641°,N24.465° 伦教40 Lunjiao 40 2 广东罗定 Luoding, Guangdong E111.570°,N22.768° 伦教40 Lunjiao 40 2 广东广州 Guangzhou, Guangdong E113.359°,N23.170° 伦教40 Lunjiao 40 2 广东英德 Yingde, Guangdong E113.454°,N24.179° 伦教40 Lunjiao 40 3 广西都安 Du’an, Guangxi E108.105°,N23.931° 粤桑11 Yuesang 11 3 广西来宾 Laibin, Guangxi E109.234°,N23.727° 粤桑11 Yuesang 11 4 广西环江 Huanjiang, Guangxi E108.258°,N24.826° 农桑14 Yuesang 11 2 广西忻城 Xincheng, Guangxi E108.665°,N24.066° 粤桑11 Yuesang 11 2 广西宜州 Yizhou, Guangxi E108.636°,N24.485° 粤桑11 Yuesang 11 4  下载: 导出CSV
下载: 导出CSV
表 2 用于扩增茄科雷尔氏菌果胶裂解酶基因的引物序列
Table 2 Primer sequences used for amplifying pectate lyase genes of Ralstonia solanacearum
引物组编号
Primer mix number引物名称
Primer name引物序列(5′→3′)
Primer sequence (5′→3′)长度/bp
Length1 DsF-1 GTGTCTCAGTCCCAGTGTGGCCTAGTTGGTGGGGTAAAGGC 41 DsR-1 GACAATGGGGGCAACCCTGATCCCAGGTATTACCCAGTGCGA 42 FIT-1 CTGCCTCCCGTAGGAGCCTGCCAAGGCGAACGATCAGTAG 40 RIT-1 CAGGCTCCTACGGGAGGCAGTCTTTCCGGACAAAAGTGCT 40 SteF-1 GACAATGGGGGCAACCCTGATC 22 SteR-1 GTGTCTCAGTCCCAGTGTGGC 21 2 DsF-2 GTACTGCGCGCTGGAGCTGGGAACAACCATTTCCAGACGC 40 DsR-2 GGAACATGCTGAACCAGCCGGGGCCGTTTGCACCGAACC 39 FIT-2 AGCTTGCTCGCTTTGAGCTGCCCTCGACGGGGAATCAC 38 RIT-2 GCAGCTCAAAGCGAGCAAGCTGCCGCCATGATGACCG 37 SteF-2 GGAACATGCTGAACCAGCCGGG 22 SteR-2 GTACTGCGCGCTGGAGCTGG 20 3 DsF-3 GTACTGCGCGCTGGAGCTGGACCATTTCCAGACGCC 36 DsR-3 GGAACATGCTGAACCAGCCGGGGCCGTTTGCACCGAACC 39 FIT-3 GAACAGCTTGCTCGCTTTGAGCTCTCGACGGGGAATCACGA 41 RIT-3 AGCTCAAAGCGAGCAAGCTGTTCGCCGCCATGATGACCG 39 SteF-3 GGAACATGCTGAACCAGCCGGG 22 SteR-3 GTACTGCGCGCTGGAGCTGG 20 4 DsF-4 AACATGCTGAACCAGCCGGGCGCCGTTTGCACCGAACC 38 DsR-4 GTACTGCGCGCTGGAGCTGGACAACCATTTCCAGACGCC 39 FIT-4 GAACAGCTTGCTCGCTTTGAGCTTCGACGGGGAATCACGA 40 RIT-4 AGCTCAAAGCGAGCAAGCTGTTCCGAGCCGCCATGATGAC 40 SteF-4 AACATGCTGAACCAGCCGGGC 21 SteR-4 GTACTGCGCGCTGGAGCTGG 20 5 DsF-5 AGCAGTTGTACTGCGCGCTGGCCATTTCCAGACGCCCTC 39 DsR-5 CGACAACGCCGGCCAGAAGGCCGTTTGCACCGAAC 35 FIT-5 CGAACAGCTTGCTCGCTTTGAGGAATCACGATGCGGGTTCC 41 RIT-5 TCAAAGCGAGCAAGCTGTTCGTCGAGCCGCCATGATGAC 39 SteF-5 CGACAACGCCGGCCAGAA 18 SteR-5 AGCAGTTGTACTGCGCGCTGG 21 6 DsF-6 AGCAGTTGTACTGCGCGCTGCCATTTCCAGACGCCCTC 38 DsR-6 CGACAACGCCGGCCAGAAGGCCGTTTGCACCGAAC 35 FIT-6 CGAACAGCTTGCTCGCTTTGAGAATCACGATGCGGGTTCC 40 RIT-6 TCAAAGCGAGCAAGCTGTTCGGGAATCGAGCCGCCATGATGAC 43 SteF-6 CGACAACGCCGGCCAGAA 18 SteR-6 AGCAGTTGTACTGCGCGCTG 20
下载: 导出CSV
表 3 外、茎、内引物体积配比
Table 3 Volume ratio of outer, stem and inner primers
μL 外、茎、内引物体积比
Volume ratio of primers
(Outer∶Stem∶Inner)外引物
Outer primer茎引物
Stem primer内引物
Inner primer无菌水
Sterilized ddH2O总和
TotalDsF DsR SteF SteR FIT RIT 1∶4∶8 2 2 8 8 16 16 48 100 1∶2∶4 2 2 4 4 8 8 72 100 1∶1∶2 2 2 2 2 4 4 84 100 1∶8∶4 2 2 16 16 8 8 48 100
下载: 导出CSV
-
[1] LIM S H, CHOI C I. Pharmacological properties of Morus nigra L. (black mulberry) as a promising nutraceutical resource[J]. Nutrients, 2019, 11(2): 437. doi: 10.3390/nu11020437.
[2] YUAN Q, ZHAO L. The mulberry (Morus alba L. ) fruit: A review of characteristic components and health benefits[J]. Journal of Agricultural and Food Chemistry, 2017, 65(48): 10383-10394. doi: 10.1021/acs.jafc.7b03614
[3] CAI M, MU L, WANG Z, et al. Assessment of mulberry leaf as a potential feed supplement for animal feeding in P. R. China[J]. Asian-Australasian Journal of Animal Sciences, 2019, 32(8): 1145-1152. doi: 10.5713/ajas.18.0671
[4] 王越, 童金林, 叶伟清, 等. 桑青枯病发生原因及防治对策[J]. 中国蚕业, 2006, 27(4): 25-26. doi: 10.3969/j.issn.1007-0982.2006.04.009 [5] 肖练章, 朱志德, 何坤良, 等. 桑细菌性青枯病发病规律研究[J]. 广东蚕丝通讯, 1982(2): 3-5. [6] 朱燕, 叶志毅, 吕志强, 等. 桑树青枯病的分布危害和防治的研究进展[J]. 蚕桑通报, 2005, 36(1): 6-9. doi: 10.3969/j.issn.0258-4069.2005.01.002 [7] 毛铿祖. 广东桑青枯病的发生与防治[J]. 中国蚕业, 1996, 30(4): 64-66. [8] 赖文姜, 曾宪铭, 谭炳安, 等. 桑青枯病病原细菌的鉴定[J]. 华南农学院学报, 1982, 3(1): 66-73. [9] LOWE-POWER T M, KHOKHANI D, ALLEN C. How Ralstonia solanacearum exploits and thrives in the flowing plant xylem environment[J]. Trends in Microbiology, 2018, 26(11): 929-942. doi: 10.1016/j.tim.2018.06.002
[10] BRAGARD C, DEHNEN-SCHMUTZ K, DI SERIO F, et al. Pest categorisation of the Ralstonia solanacearum species complex[J]. EFSA Journal, 2019, 17(2): 5618. doi: 10.2903/j.efsa.2019.5618.
[11] NAKANO M, ICHINOSE Y, MUKAIHARA T. Ralstonia solanacearum type III effector RipAC targets SGT1 to suppress effector-triggered immunity[J]. Plant and Cell Physiology, 2020, 61(12): 2067-2076.
[12] SUN T, WU W, WU H, et al. Ralstonia solanacearum elicitor RipX induces defense reaction by suppressing the mitochondrial atpA gene in host plant[J]. International Journal of Molecular Sciences, 2020, 21(6): 2000. doi: 10.3390/ijms21062000.
[13] SCHÖNFELD J, HEUER H, VAN ELSAS J D, et al. Specific and sensitive detection of Ralstonia solanacearum in soil on the basis of PCR amplification of fliC fragments[J]. Applied and Environmental Microbiology, 2003, 69(12): 7248-7256. doi: 10.1128/AEM.69.12.7248-7256.2003
[14] OPINA N, TAVNER F, HOLLWAY G, et al. A novel method for development of species and strain-specific DNA probes and PCR primers for identifying Burkholderia solanacearum(formerly Pseudomonas solanacearum)[J]. Asia Pacific Journal of Molecular Biology and Biotechnology, 1997, 5(1): 19-30.
[15] PIRC M, ALIČ Š, DREO T. Rapid loop-mediated isothermal amplification for detection of the Ralstonia solanacearum species complex bacteria in symptomatic potato tubers and plants[J]. Methods in Molecular Biology, 2021, 2354: 401-413.
[16] KYOSEI Y, YAMURA S, NAMBA M, et al. Antigen tests for COVID-19[J]. Biophysics and Physicobiology, 2021, 18: 28-39. doi: 10.2142/biophysico.bppb-v18.004
[17] 黄雯, 徐进, 张昊, 等. 植物青枯菌LAMP检测方法的建立[J]. 中国农业科学, 2016, 49(11): 2093-2102. doi: 10.3864/j.issn.0578-1752.2016.11.006 [18] KUBOTA R, VINE B G, ALVAREZ A M, et al. Detection of Ralstonia solanacearum by loop-mediated isothermal amplification[J]. Phytopathology, 2008, 98(9): 1045-1051. doi: 10.1094/PHYTO-98-9-1045
[19] 马学军, 丁雄, 聂凯, 等. 新型等温多自配引发扩增技术(IMSA): CN104388581A[P]. 2015-03-04. [20] 丁雄. 新型等温核酸扩增技术(IMSA)的建立及其对传染病病原EV71、CVA16、H7N9 和HIV-1的快速检测应用[D]. 广州: 华南理工大学, 2014. [21] ZHAI J, YAN Z, PING F, et al. Establishment and application of isothermal amplification techniques for the detection of heat-stable I enterotoxin of enterotoxigenic Escherichia coli[J]. PLoS One, 2020, 15(4): e0230881. doi: 10.1371/journal.pone.0230881
[22] 张梦. 溧阳市肺结核病流行情况及等温多自配引发扩增技术在检测结核分枝杆菌中的应用分析[D]. 苏州: 苏州大学, 2018. [23] 乐振窍, 许泽仰, 张细玲, 等. FQ-PCR与IMSA检测转基因豆奶外源基因的比较研究[J]. 大豆科学, 2018, 37(6): 943-949. [24] 王琪, 徐文娟, 石盼盼. IMSA技术快速检测肠出血大肠杆菌O157∶H7方法的建立及应用[J]. 食品工业科技, 2021, 42(17): 263-269. [25] 杨宏宇. 桑枯萎病病原鉴定及分子生物学研究[D]. 广州: 华南农业大学, 2018. [26] DING X, NIE K, SHI L, et al. Improved detection limit in rapid detection of human enterovirus 71 and coxsackievirus A16 by a novel reverse transcription-isothermal multiple-self-matching-initiated amplification assay[J]. Journal of Clinical Microbiology, 2014, 52(6): 1862-1870. doi: 10.1128/JCM.03298-13
[27] 董朝霞, 于翠, 邓文, 等. 桑树青枯病的发生与防治研究进展[J]. 北方蚕业, 2019, 40(4): 1-7. doi: 10.3969/j.issn.1673-9922.2019.04.001 [28] WANG D, YU J, WANG Y, et al. Development of a real-time loop-mediated isothermal amplification(LAMP) assay and visual LAMP assay for detection of African swine fever virus(ASFV)[J]. Journal of Virological Methods, 2020, 276: 113775. doi: 10.1016/j.jviromet.2019.113775.
[29] HUANG W E, LIM B, HSU C C, et al. RT-LAMP for rapid diagnosis of coronavirus SARS-CoV-2[J]. Microbial Biotechnology, 2020, 13(4): 950-961. doi: 10.1111/1751-7915.13586
[30] TECHATHUVANAN C, DRAUGHON F A, D'SOUZA D H. Loop-mediated isothermal amplification (LAMP) for the rapid and sensitive detection of Salmonella Typhimurium from pork[J]. Journal of Food Science, 2010, 75(3): M165-M172. doi: 10.1111/j.1750-3841.2010.01554.x
[31] 刘吉平, 程伟, 宋小景, 等. 一种家蚕蚕卵微孢子虫的LAMP检测引物及其应用: CN104372082A[P]. 2015-02-25. [32] 刘吉平, 周轶楠, 孙勋勋. 一组桑花叶病的LAMP检测引物及试剂盒: CN109055616A[P]. 2018-12-21. [33] 杨宏宇, 周轶楠, 孙勋勋, 等. 桑源阴沟肠杆菌LAMP检测方法的建立和应用[J]. 蚕业科学, 2019, 45(3): 321-330. [34] CHEN X, WANG H, LIU C, et al. Technical note: Development of a closed-tube isothermal multiple self-matching-initiated amplification assay for visual detection of Staphylococcus aureus in milk samples[J]. Journal of Dairy Science, 2021, 104(3): 3569-3574. doi: 10.3168/jds.2020-19023
[35] LIU W, YUAN C, ZHANG L, et al. Establishment and application of isothermal multiple-self-matching-initiated amplification (IMSA) in detecting Type II heat-labile enterotoxin of Escherichia coli[J]. PLoS One, 2019, 14(5): e0216272. doi: 10.1371/journal.pone.0216272
[36] KUBICEK C, STARR T, GLASS N. Plant cell wall-degrading enzymes and their secretion in plant-pathogenic fungi[J]. Annual Review of Phytopathology, 201, 52: 427-451.
[37] CHARKOWSKI A O, ALFANO J R, PRESTON G, et al. The Pseudomonas syringae pv. tomato HrpW protein has domains similar to harpins and pectate lyases and can elicit the plant hypersensitive response and bind to pectate[J]. Journal of Bacteriology, 1998, 180(19): 5211-5217.
[38] OKIRO L A, TANCOS M A, NYANJOM S G, et al. Comparative evaluation of LAMP, qPCR, conventional PCR, and ELISA to detect Ralstonia solanacearum in Kenyan potato fields[J]. Plant Disease, 2019, 103(5): 959-965. doi: 10.1094/PDIS-03-18-0489-RE
[39] 李信申, 黄小梅, 吴淑秀, 等. 植物青枯病菌环介导等温扩增快速检测技术研究[J]. 生物技术通报, 2021, 37(1): 272-281. [40] GOU H, BIAN Z, CAI R, et al. The colorimetric isothermal multiple-self-matchinginitiated amplification using cresol red for rapid and sensitive detection of porcine circovirus 3[J]. Frontiers in Veterinary Science, 2020, 7: 407. doi: 10.3389/fvets.2020.00407.
[41] ZHOU Y, YANG H, LIU J. Complete genome sequence of Enterobacter roggenkampii srain KQ-01, isolated from bacterial wilt-resistant mulberry cultivar YS283[J]. Plant Disease, 2021, 105(3): 688-690. doi: 10.1094/PDIS-07-20-1468-A
[42] 王继承. 桑枯萎病中菠萝泛菌的分离鉴定及防治研究[D]. 广州: 华南农业大学, 2018. [43] 罗龙辉, 王继承, 刘吉平. 桑细菌性枯萎病病原菌的分离鉴定与全基因组序列分析[J]. 植物保护, 2022, 48(1): 44-51.