Research of Brassica chinensis var. parachinensis under water stress based on machine learning
-
摘要:目的
以油青菜心Brassica chinensis var.parachinensis为试验对象,基于冠层温度研究其生长过程中的水分胁迫变化规律,并利用机器学习方法,以水分胁迫指数(Crop water stress index, CWSI)和光合有效辐射预测光合作用速率。
方法试验期间,在营养生长阶段(V期)和生殖生长阶段(R期)对油青菜心进行不同田间持水量处理,采集冠层温度、空气温湿度数据,建立无蒸腾作用基线(上限方程)、无水分胁迫基线(下限方程),通过经验公式计算CWSI。利用基于密度的空间聚类方法和空气温度研究油青菜心的冠气温差上限分布情况,选取固定值作为上限;以CWSI经验公式为基础,使用不同温度定值的无蒸腾作用基线计算CWSI,验证聚类效果。为更简便获取光合作用速率,使用4种机器学习方法:最邻近节点算法(k-Nearest neighbor,KNN)、支持向量回归(Support vector regression,SVR)、极端梯度提升法(Extreme gradient boosting,XGBoost)、随机森林(Random forest,RF)进行预测,并对比预测效果。
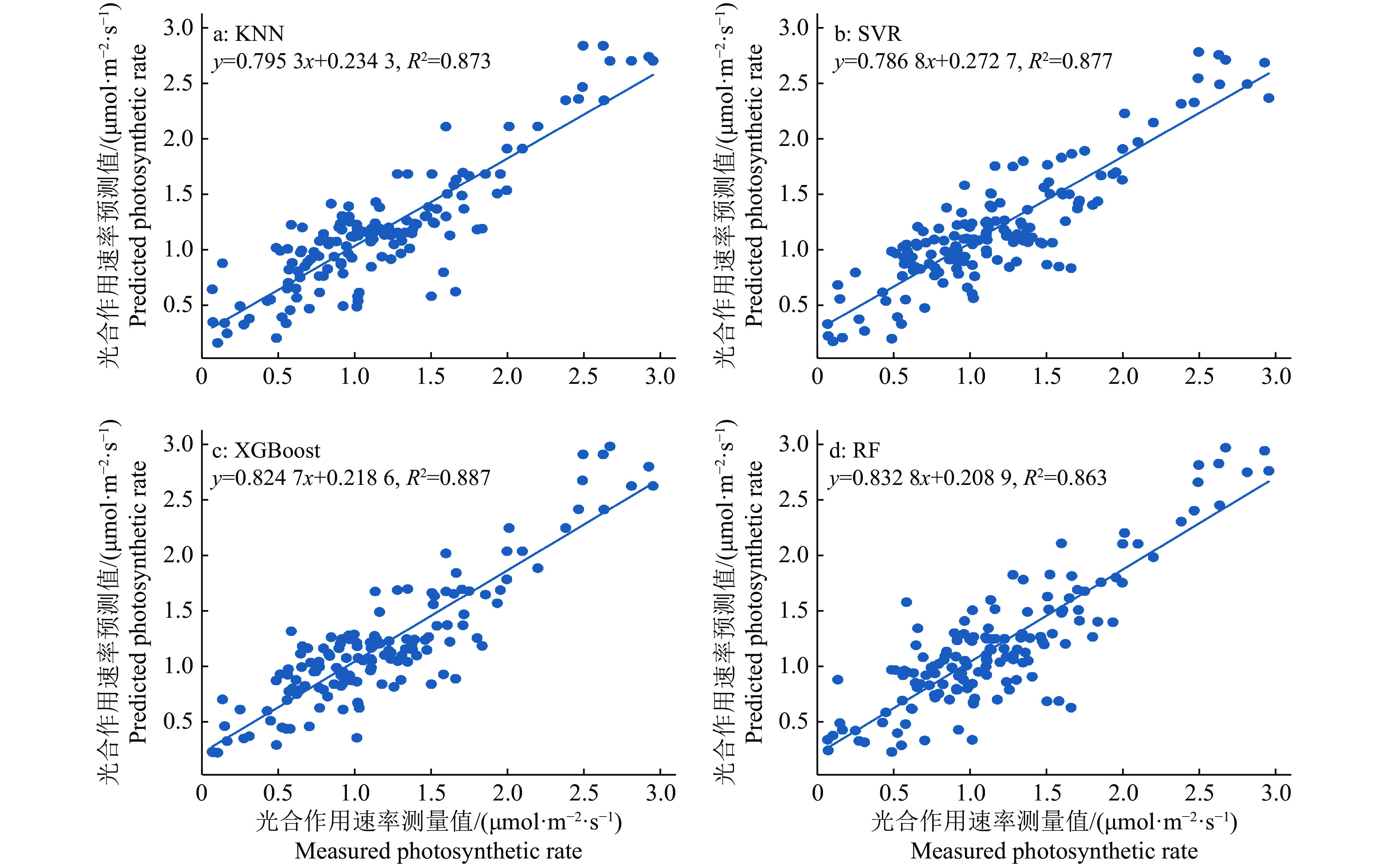
结果在不同田间持水量处理下,CWSI能较好地监测油青菜心水分胁迫状况。通过聚类分析,将V期和R期冠气温差上限分类到2个簇中,得到簇心分别为3.4和4.2 ℃,与CWSI经验公式计算值显著相关,表明使用固定值作为油青菜心冠气温差上限值具有可行性。KNN、SVM、XGBoost和RF预测模型均取得较好效果,相关系数分别为0.873、0.877、0.887和0.863。
结论机器学习方法可用于油青菜心光合作用速率的预测,可以避免使用大型笨重仪器,降低对油青菜心叶片的损伤,减少测量时间。
Abstract:ObjectiveBrassica chinensis var. parachinensis was used as the experimental object to study the change rule of water stress during growth process based on canopy temperature, and the machine learning method was used for predicting the photosynthetic rate based on crop water stress index (CWSI) and photosynthetically active radiation.
MethodDuring the test, the experiment adopted different field capacities for B. chinensis var. parachinensis at the vegetative growth stage (V stage) and reproductive growth stage (R stage), collected the canopy temperature, air temperature and humidity data, established non-transpiration-baseline (upper limit equation), non-water-stress-baseline (lower limit equation), and calculated CWSI by empirical formulation. Cluster method of density-based spatial clustering of application with noise and air temperature were used to study the upper limit distribution of canopy temperature minus air temperature of B. chinensis var. parachinensis, and the fixed values were selected as the upper limit. Based on the CWSI empirical formulation, CWSI was calculated using the non-transpiration-baselines with different temperature fixed values to verify the clustering effect. In order to obtain the photosynthetic rate more easily, four machine learning methods of k-nearest neighbor (KNN), support vector regression (SVR), extreme gradient boosting (XGBoost) and random forest (RF) were used for prediction, and the prediction effects were compared.
ResultUnder different field capacities, CWSI could better monitor the water stress status of B. chinensis var. parachinensis. Through cluster analysis, the upper limit of canopy temperature minus air temperature at V stage and R stage was classified into two clusters, and the cluster centers were 3.4 and 4.2 ℃, respectively, which were significantly correlated with the values calculated by the empirical formula of CWSI, indicating that it was feasible to use a fixed value as the upper limit of canopy temperature minus air temperature in B. chinensis var. parachinensis. The prediction models of KNN, SVM, XGBoost and RF all achieved good results, and the correlation coefficients were 0.873, 0.877, 0.887 and 0.863, respectively.
ConclusionMachine learning can be used for predicting the photosynthetic rate of B. chinensis var. parachinensis, avoid the use of large and cumbersome instruments, reduce the damage to the leaves of B. chinensis var. parachinensis, and reduce the measurement time.
-
森林在改良土壤、涵养水源、改善林下植物多样性等方面发挥着巨大的作用[1]。土壤作为森林的基本组成部分,其土壤物理性质能反映出土壤结构、养分状况和持水、渗透能力,对林地土壤质量评价具有重要意义。林下植物作为森林生态系统的重要结构,其物种多样性是反映植物群落组成和稳定性的重要指标[2],易受立地条件、环境因子和林分郁闭度等因素的影响[3]。由于林下植物具有改良土壤、持水固土以及促进生态系统物质循环的作用[4],因此对两者关系的研究,有利于对森林系统的土壤管理及植物多样性保护。
亚热带人工林作为我国森林生态系统的重要组成之一,具有分布广和面积大的特点[5]。随着社会经济的发展,人类对人工林需求量的快速增长以及经营方式的不当造成人工林出现土壤肥力衰退、水土流失加剧和生态多样性下降等问题。有研究表明,改善林分结构是提高人工林林地质量和土地生产力的途径之一[6]。目前,国内外关于不同林分类型的人工林林地土壤物理性质[7-8]、植被多样性[9-10]已有一定的报道,二者间的关系也在马尾松Pinus massoniana林[11]、草地[12]和热带山地森林[13]中进行过研究,但缺乏对亚热带人工林的土壤物理性质和生物多样性关系的研究。
本文以亚热带人工林最典型的3 种林分类型—阔叶纯林、阔叶混交林和针阔混交林作为研究对象,对其土壤物理性质和生物多样性进行调查研究,以期能有效地揭示不同人工林类型之间的差异及相关因素,为亚热带人工林植被恢复选择合理的措施及优化模式提供参考。
1. 材料与方法
1.1 试验地概况
试验地原为杉木Cunninghamia lanceolata林,位于广东省佛山市云勇生态林养护中心(112°40′E,22°53′N),森林面积1 928.73 hm2。该地属亚热带季风性湿润气候,雨量充沛,年均降雨量为2 100 mm,集中于6—8月,年平均相对湿度80%。土壤为花岗岩发育的酸性赤红壤(pH<5),土层厚达1 m以上,土地肥沃。
2005年起,为了营造生态公益林,对原杉木林进行皆伐改造,引入不同类型乡土树种造林。在贼佬坑林地栽种石笔木Tutcheria championii(P1样地),在白石岗林地栽种以杉木、米老排Mytilaria laosensis为主,以尖叶杜英Elaeocarpus apiculatus、火力楠Michelia macclurel、枫香Liquidambar formosana、盆架子Alstonia scholarisv为辅的针阔混交林(P2样地),在仙人春林地栽种以米老排为主,以红锥Castanopsis hystrix、格木Erythrophleum fordii为辅的阔叶混交林(P3样地),并对以上林分采取相同的管理措施。各样地的基本情况见表1,样地的主要树种生长情况见表2。
表 1 试验林概况Table 1. General situation of the experimental plantations样地 海拔/m 坡向 坡度/(°) 郁闭度 主要树种 P1 172 S 42 0.6 石笔木 P2 259 SE 35 0.6 杉木、米老排等 P3 204 SW 40 0.8 米老排等 表 2 树木生长基本情况1)Table 2. Tree growth status of the experimental plantations样地 树种 胸径/cm 树高/m 冠幅/m P1 石笔木 8.59±2.46 7.00±1.00 5.01±1.00 P2 杉木 8.59±1.82 21.26±3.25 8.95±3.21 米老排 12.28±2.21 14.24±2.21 9.96±2.82 尖叶杜英 9.72±3.37 9.01±2.85 3.97±1.57 火力楠 10.80±2.08 9.60±1.56 4.76±0.91 枫香 11.33±2.78 10.95±2.16 6.13±0.74 盆架子 8.16±0.68 12.80±1.92 6.78±0.97 P3 米老排 9.24±3.83 10.92±4.44 5.32±1.75 红锥 11.43±4.38 12.52±3.07 6.78±2.29 格木 4.30±0.73 4.52±0.17 5.65±0.89 1)表中数据为平均值±标准差 1.2 试验方法
1.2.1 样地设置
2016年5月对试验地3种生态公益林进行调查(林分的林龄均为10年生),每种林地设定3个规格为20 m ×20 m的调查样地。用容积100 cm3的环刀分别在各个标准地的0~20 cm土层采集土壤样品,用于测定土壤的物理性质(3个重复)。按梅花形在每个标准样地内设置5个面积为2 m×2 m的样方,样地调查并记录小样方内全部灌木和草本的种类、盖度、株数及平均高度,用于分析林下植物的物种多样性。
1.2.2 林下植物多样性调查方法
林下植物的物种多样性指数计算包括下列内容[14]:
物种丰富度指数表示一个种群在群落中的个体数目或丰富程度,这里是指某物种在标准地中的总数,即样方内所有物种数目(S);多度为某一种植物的个体总数;盖度=植物地上部分垂直投影面积/样方面积;频度=该种植物出现的样方数目/所有调查的样方总数目;相对多度=某一种植物的个体总数/同一生活型植物个体总数×100%;相对盖度=某一种植物的盖度/所有种的盖度总和×100%;相对频度=某一个种的频度/所有种的频度总和×100%;相对重要值=(相对多度+相对盖度+相对频度)/3。
Simpson指数(D)、Shannon-Wiener指数(H)及Pielou均匀度指数(JSW和JSi)分别为:
$$ D = 1 - {\sum\limits_{i = 1}^s {P_i} ^2}, $$ (1) $$ H = - \sum\limits_{i = 1}^s {(P_i\ln P_i)} , $$ (2) $$ {J_{{\rm{SW}}}} =H/{\rm ln}S, $$ (3) $$ {J_{{\rm{Si}}}} = D/(1 - 1/S), $$ (4) 式中,Pi为第 i 物种的相对重要值。
1.2.3 土壤物理性质的测定
土壤含水量采用烘干法测定[15];土壤容重、土壤总孔隙度、土壤毛管孔隙度等物理性质均采用环刀法测定[16],每个指标重复测定3次。
1.3 数据处理及分析
运用Microsoft Excel 2003对数据进行统计分析和图表绘制,采用SPSS 22.0统计分析软件进行单因素方差分析(One-way ANOVA),并采用Duncan’s法对试验结果进行多重比较。
2. 结果与分析
2.1 林下植物多样性
2.1.1 林下植物种类组成与相对重要值
由表3可知,P1样地林下灌木有5种,其中相对重要值最大的是鸭脚木Schefflera octophylla(24.09%),其次是野牡丹Melastoma candidum(21.27%)。P2样地和P3样地林下灌木种类均为2种,分别为白花酸藤果Embelia ribes和山苍子Litsea cubeba 、银柴Aporosa dioica和南天竹Nandina domestica,它们的相对重要值均为50%。
表 3 林下灌木种类与相对重要值Table 3. Undergrowth shrub layer species and importance value %物种 P1样地 P2样地 P3样地 鸭脚木 Schefflera octophylla 24.09 银柴 Aporosa dioica 19.44 50.00 白花酸藤果 Embelia ribes 19.44 50.00 黑面神 Breynia fruticosa 15.77 野牡丹 Melastoma candidum 21.27 山苍子 Litsea cubeba 50.00 南天竹 Nandina domestica 50.00 由表4可知,P1样地林下草本植物有12种,相对重要值前3位为弓果黍Cyrtococcum patens (24.29%),乌毛蕨Blechnum orientale (21.82%)和蔓生莠竹Microstegium vagans(20.68%)。P2样地林下草本植物有4种,相对重要值大的是乌毛蕨(44.11%)和华南鳞盖蕨Microlepia hancei(43.26%)。P3样地林下草本植物有6种,相对重要值最大的是乌毛蕨(52.30%),其次是半边旗Pteris semipinnata(17.33%)。在3个林地当中,乌毛蕨在每个林地中所占相对重要值均超过20%。
表 4 林下草本种类与相对重要值Table 4. Undergrowth herb layer species and importance value物种 P1样地 P2样地 P3样地 乌毛蕨 Blechnum orientale 21.82 44.11 52.30 玉叶金花 Mussaenda pubescens 4.63 7.42 蔓生莠竹 Microstegium vagans 20.68 半边旗 Pteris semipinnata 7.00 5.49 17.33 弓果黍 Cyrtococcum patens 24.29 华南毛蕨 Cyclosorus parasiticus 3.95 崩大碗 Centella asiatica 1.54 芒箕 Dicranopteris dichotoma 2.10 山菅兰 Dianella ensifolia 3.36 火炭母 Polygonum chinense 1.54 9.87 粽叶芦 Thysanolaena maxima 4.03 金盏银盘 Bidens biternata 5.07 华南鳞盖蕨 Microlepia hancei 43.26 崇澍蕨 Chieniopteris harlandii 7.14 扇叶铁线蕨 Adiantum flabellulatum 4.72 淡竹叶 Lophatherum gracile 8.36 2.1.2 林下植物多样性指数及均匀度指数
林下灌木的多样性指数和均匀度指数列于表5。由表5可知,P1样地的Simpson指数、Shannon-Wiener指数和均匀度指数JSW均最大,P2样地及P3样地的均匀度指数JSi最大。
表 5 林下灌木多样性指数与均匀度指数Table 5. Plant diversity index and evenness index of undergrowth shrub layer样地 Simpson
指数Shannon-Wiener
指数均匀度指数 JSW JSi P1 0.79 1.60 0.62 0.61 P2 0.50 0.69 0.49 1.79 P3 0.50 0.69 0.49 1.79 林下草本的多样性指数和均匀度指数列于表6。由表6可知,P1样地的Simpson指数、Shannon-Wiener指数均最大,P3样地的均匀度指数JSW最大,P2样地的均匀度指数JSi最大。
表 6 林下草本植物多样性指数与均匀度指数Table 6. Plant diversity index and evenness index of undergrowth herb layer样地 Simpson
指数Shannon-Wiener
指数均匀度指数 JSW JSi P1 0.84 2.01 0.32 0.81 P2 0.61 1.07 0.28 0.97 P3 0.67 1.42 0.34 0.87 2.2 3种改造人工生态公益林的土壤物理性质分析
各样地的土壤物理性质见表7。各样地土壤容重大小排序为P1样地(1.33 g·cm–3)>P3样地(1.25 g·cm–3)>P2样地(1.08 g·cm–3),P1和P3样地的土壤容重显著大于P2样地。
表 7 土壤物理性质1)Table 7. Soil physical properties样地 土壤容重/
(g·cm–3)总孔隙度/
%自然含水量(w)/
%毛管持水量(w)/
%毛管孔隙度/
%非毛管孔隙度/
%通气孔隙度/
%P1 1.33±0.010a 49.91±0.363b 24.11±0.499c 30.93±1.543c 4.11±0.192b 45.81±0.344b 46.71±0.435b P2 1.08±0.022b 59.32±0.823a 38.72±0.934a 46.13±1.249a 4.97±0.058a 54.35±0.801a 55.15±0.822a P3 1.25±0.088a 52.78±3.332b 33.32±2.339b 37.84±4.059b 4.71±0.156a 48.07±3.179b 48.63±3.345b 1)表中数据为3次重复的平均值±标准误;同列数据后,凡是具有一个相同小写字母者表示样地间差异不显著(P>0.05,Duncan’s 法) 各样地土壤总孔隙度大小排序为P2样地(59.32%)>P3样地(52.78%)>P1样地(49.91%),P2样地的土壤总孔隙度显著大于其他2个林地。
各样地土壤自然含水量(w)大小排序为P2样地(38.72%)>P3样地(33.32%)>P1样地(24.11%),P3样地的自然含水量显著小于P2样地,但显著大于P1样地。
各样地土壤毛管持水量(w)大小排序为P2样地(46.13%)>P3样地(37.84%)>P1样地(30.93%),且3个样地间差异显著。
各样地土壤毛管孔隙度大小排序为P2样地(4.97%)>P3样地(4.71%)>P1样地(4.11%),P2样地和P3样地显著大于P1样地。
各样地土壤非毛管孔隙度大小排序为P2样地(54.35%)>P3样地(48.07%)>P1样地(45.81%),P2样地的土壤非毛管孔隙度显著大于P1和P3样地。
各样地土壤通气孔隙度排序为P2样地(55.15%)>P3样地(48.63%)>P1样地(46.71%)。P2样地的土壤通气孔隙度显著大于P1和P3样地。
3. 讨论与结论
在3个改造人工林地当中,灌木层的Shannon-Wiener和Simspon指数的平均值(0.99和0.60)均小于草本层(1.50和0.71);灌木层的Pielou均匀度指数JSW和JSi平均值分别为0.53和1.40,均大于草本层(0.31和0.88);阔叶纯林的林下灌木和草本种类及其对应的优势种数量均多于混交林。
本研究的3种林地中,P1样地的土壤容重最大;P2样地的乔木植物种类较多且冠幅、树高和胸径较大;除土壤容重外,其余各项物理指标均表现为P2样地>P3样地>P1样地。
物种多样性是生态系统及生物群落的本质特征[17],能有效反映生物群落和生态系统的复杂性、稳定性,所以生物多样性是生态系统结构和功能的测量指标。Shannon-Wiener指数是反映物种丰富度的指数,物种数量越多,其数值越大;Simpson指数主要反映物种的多度,是一个反映群落优势度的指数,其数值越小,表明群落的优势种越明显。在3个林地中,灌木层的Shannon-Wiener和Simspon指数的平均值均小于草本层,可见各林地中的草本层物种数量多于灌木层。与草本相比,灌木对生长环境的要求更高,随着时间的推移,上层林木不断壮大、林分郁闭度不断增加,使得林下灌木对生长所需的光照、养分等条件的竞争加强[18],这将限制林下灌木层的生长。另外,草本植物的适应性比灌木强,因此林下草本植物发育较好[19]。
Pielou均匀度指数是反映群落均匀度的指标,与群落内物种个数间的差异及各物种个体数量分配均匀程度有关[20],种间的个体数量差异程度越小,则群落的均匀性越高[21]。在3个林地中,灌木层的Pielou均匀度指数(JSW和JSi)平均值均大于草本层,可见各林地中灌木层的物种均匀性高于草本层。由于林下灌木植物的种类数量相对少,且个体较草本植物大,所以其在林地的分布相对均匀。而草本植物虽然数量较多,但是均匀度却不高,这可能是3个林地的郁闭度相对较高,林隙分布不均,促使阴生、蕨类植物的富集所致[10]。此外,草本植物的生活史短暂,对环境变化反应敏感,具有易受干扰和恢复速度快的特点[18],更加深了分布不均的程度。
阔叶纯林的林下灌木和草本植物种类及其对应的优势种数量均多于混交林,这可能是因为P1样地的乔木仅有石笔木1种,林冠层结构简单,林下光照环境多样化,喜阳植物和随机入侵种较多的缘故[11],另外,各样地处于幼林向中龄林过度的阶段,已基本成林,土壤水分条件较好,并且郁闭度较大,为一些阴生植物的生长和定居提供了良好的生存条件,所以阴生植物的种类和数量较多。虽然阔叶纯林单一的结构有利于草本层物种多样性的改善,但草本植物多以林下随机生长为主,均匀度指数较低;另外2种混交林的多样性指数较低,但其林下草本均匀度指数较高,群落结构相对合理[22]。
土壤物理性质与土壤持水和保水能力有着密切关系[23],可以影响土壤的肥力状况。在本研究中,所有林地的土壤总孔隙度在49.91%~59.32%,在中国大部分森林生态系统土壤总孔隙度范围(40%~60%)内[8]。当土壤中总孔隙度在50%左右,并且非毛管孔隙度占 1/5~2/5 时,土壤的通气性、透水性和持水能力较强[24]。本研究的所有样地非毛管孔隙度在92%左右,明显偏高。这说明3种林地的吸、持水分能力低,但土壤的雨水下渗能力较强,有利于减少地表径流,增强土壤的抗冲刷和抗侵蚀能力[25]。土壤容重作为森林土壤物理性质的优良指标,对森林土壤的结构性、土壤的孔性、通水透气性能以及持水能力具有重要影响[26]。通常容重越小,表明土壤可形成的孔隙越大,土壤持水和通气能力越强[8]。由于P1样地仅一种乔木,乔木植物种类不如P2样地丰富,郁闭度又较P3样地小,使其根系对土壤穿插和切割作用较小,回归土壤的枯落物少,林地土壤团粒结构的形成相对缓慢[27],从而导致土壤疏松程度较小,土壤较为紧实,容重相对较大。虽然P1样地的林下草本植物丰富,但是其根系对土壤的穿插和切割作用不及乔木。这是因为乔木的根系数量高于草本植物,并且前者的主根和侧根更发达,根系分布较深,因此对土壤物理性质的改良效果较好[28]。P2样地由于其植物根系对土壤的穿插作用较强,特别是杉木的侧根发达,穿插能力强,所以其土壤容重最小,孔隙多。这也说明杉木与阔叶树种混交,可以改善土壤物理性质,增强水土保持的能力。总体来说,混交林的各项物理性质优于纯林,这与顾宇书等[29]的研究结果相一致。
植物多样性与土壤物理性质关系密切[30]。林下植被的蒸腾作用通常占到森林总蒸腾作用的20%~30%[31],因而林下植被在一定程度上能减少土壤的水分含量[32]。在本研究中,P1样地的林下植物最为丰富且数量最多,土壤含水量较其他林地低,原因可能是林下植物的叶面积较大,植物从土壤吸收的水分需求量增多,导致土壤水分减少。
-
![]()
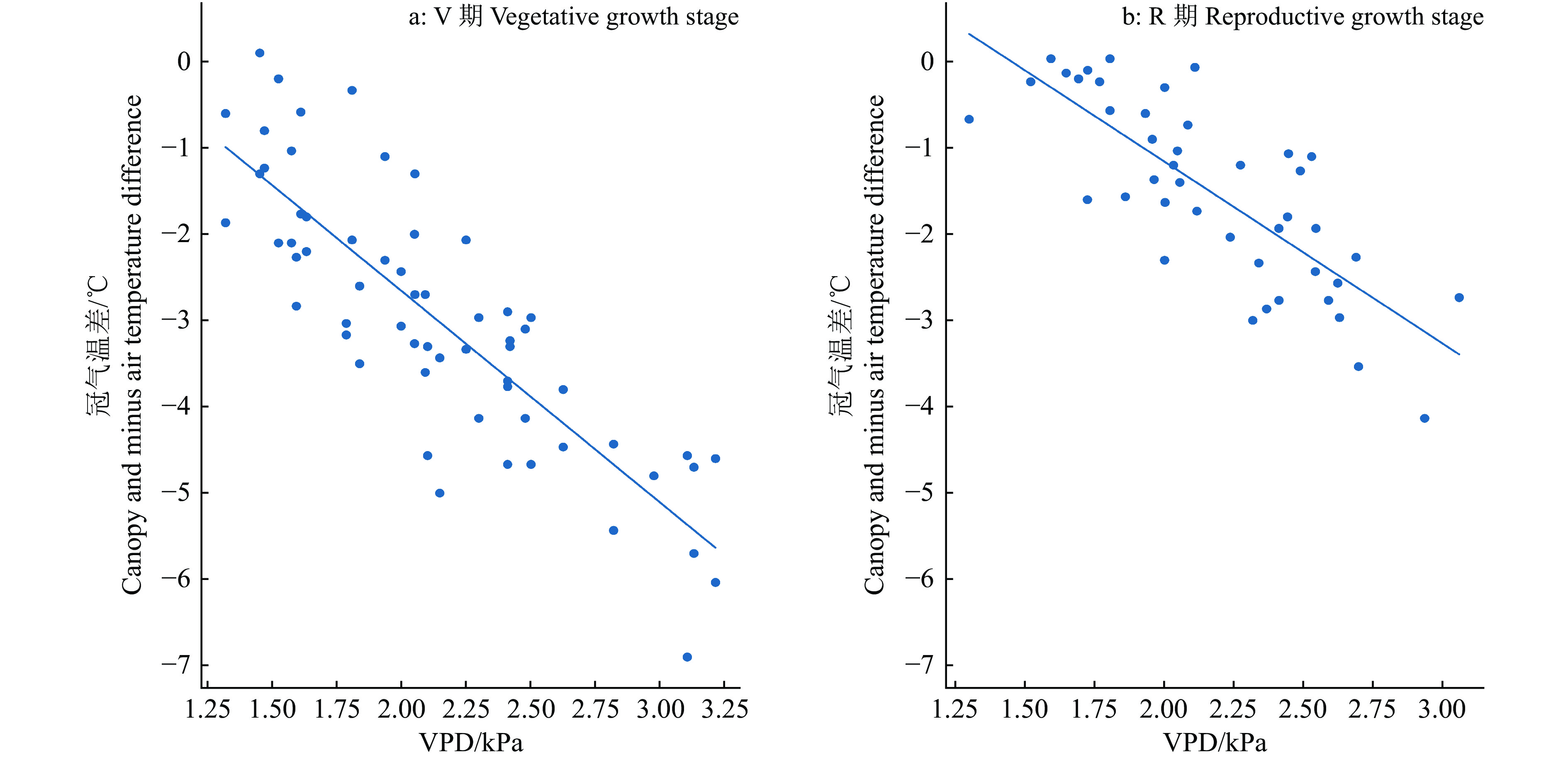
图 2 冠气温差与饱和水汽压差(VPD)的关系
Figure 2. Relationship between canopy and air temperature difference and vapor pressure deficit (VPD)
![]()
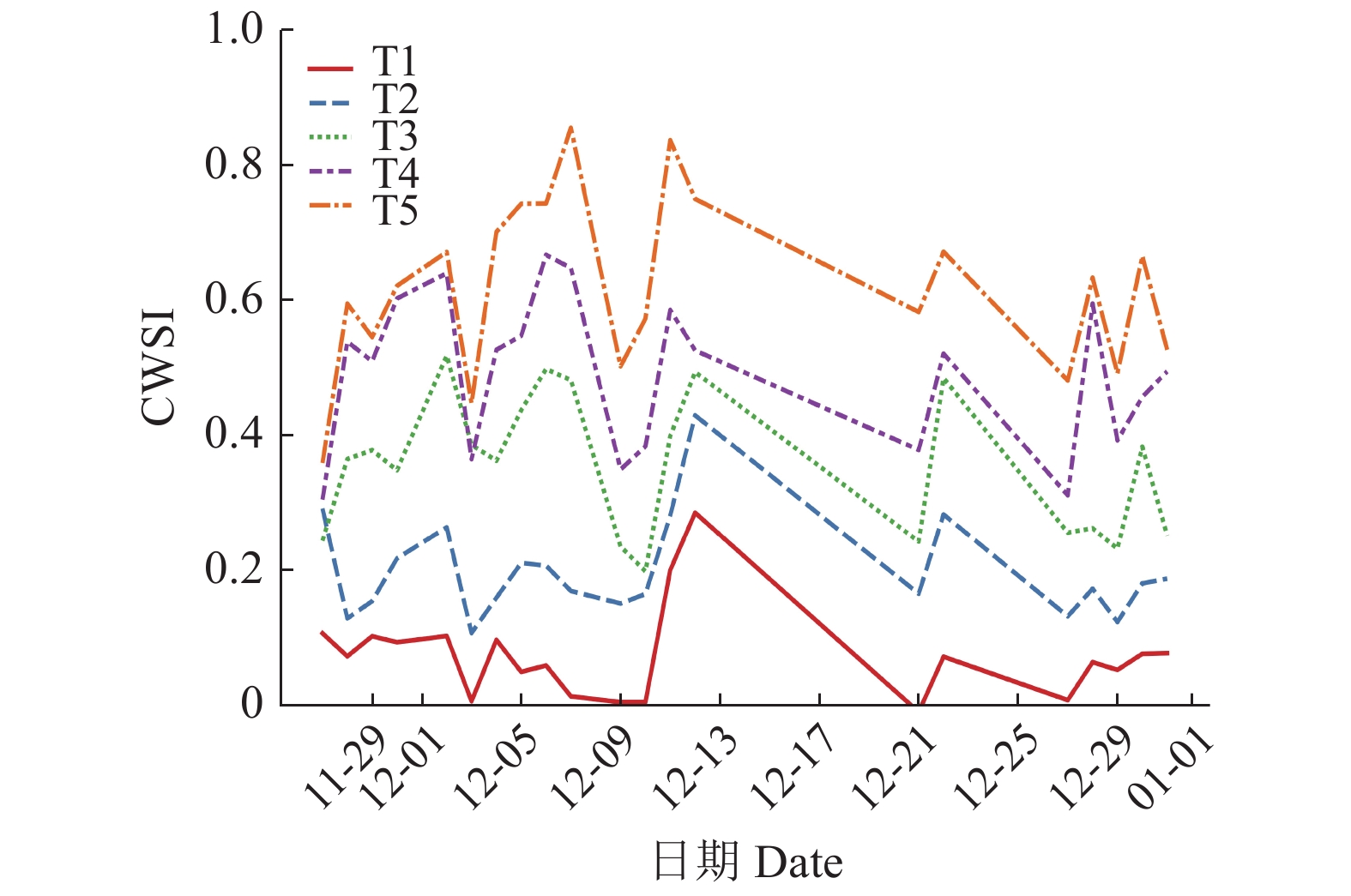
图 3 5种田间持水量处理的水分胁迫指数(CWSI)变化曲线
T1:田间持水量为32.2%;T2~T5:田间持水量分别为T1的85%、70%、55%和40%
Figure 3. Crop water stress index(CWSI) change curves under five field water holding capacities
T1: Field water holding capacity was 32.2%; T2~T5: Field water holding capacity was 85%, 70%, 55% and 40% of T1 respectively
![]()
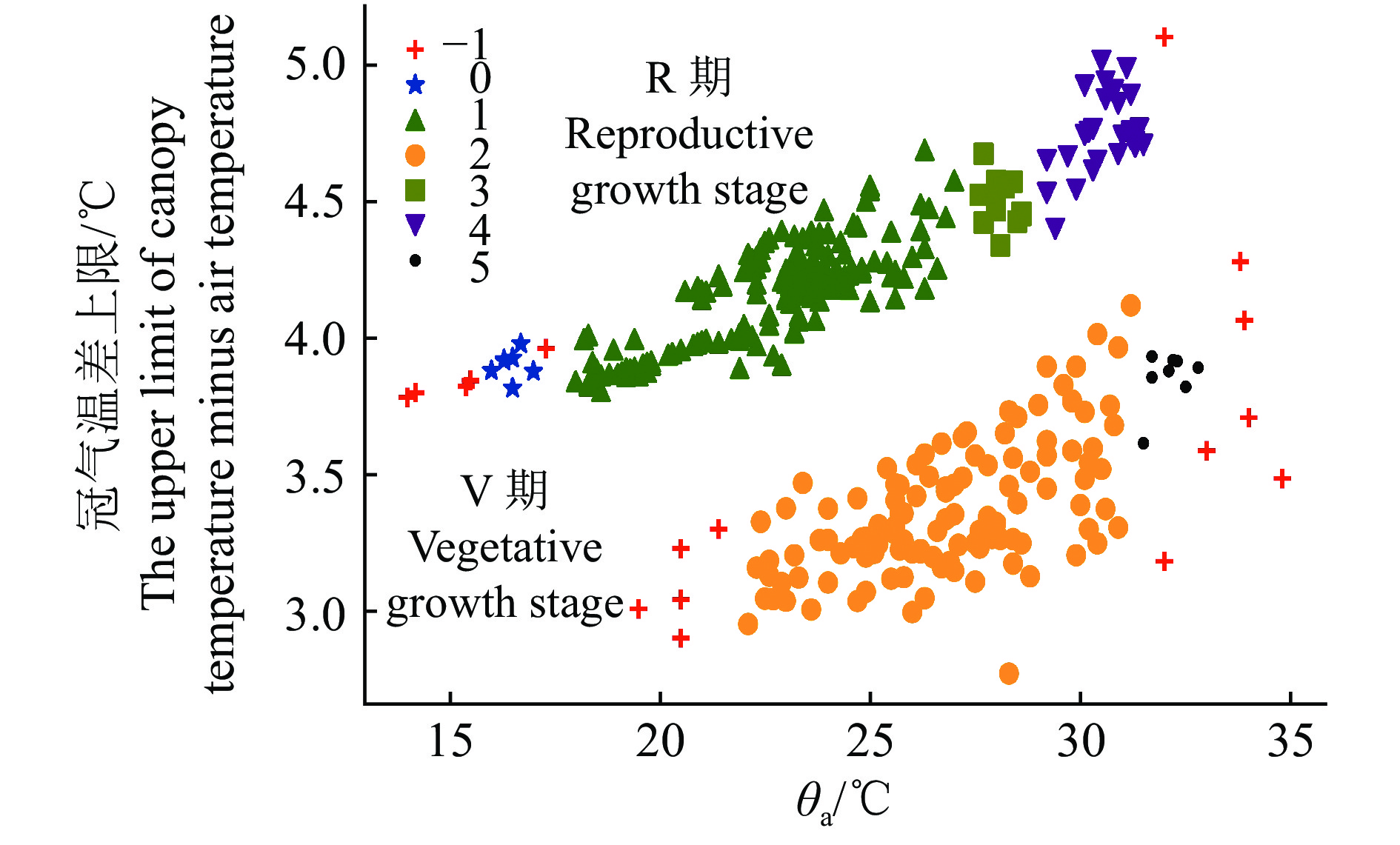
图 4 基于DBSCAN的冠气温差上限聚类
−1表示离群点,0~5表示聚类形成的簇
Figure 4. Clusters of the upper limit of canopy temperature minus air temperature by DBSCAN
−1 means outliers, 0-5 means clusters formed by clusters
![]()
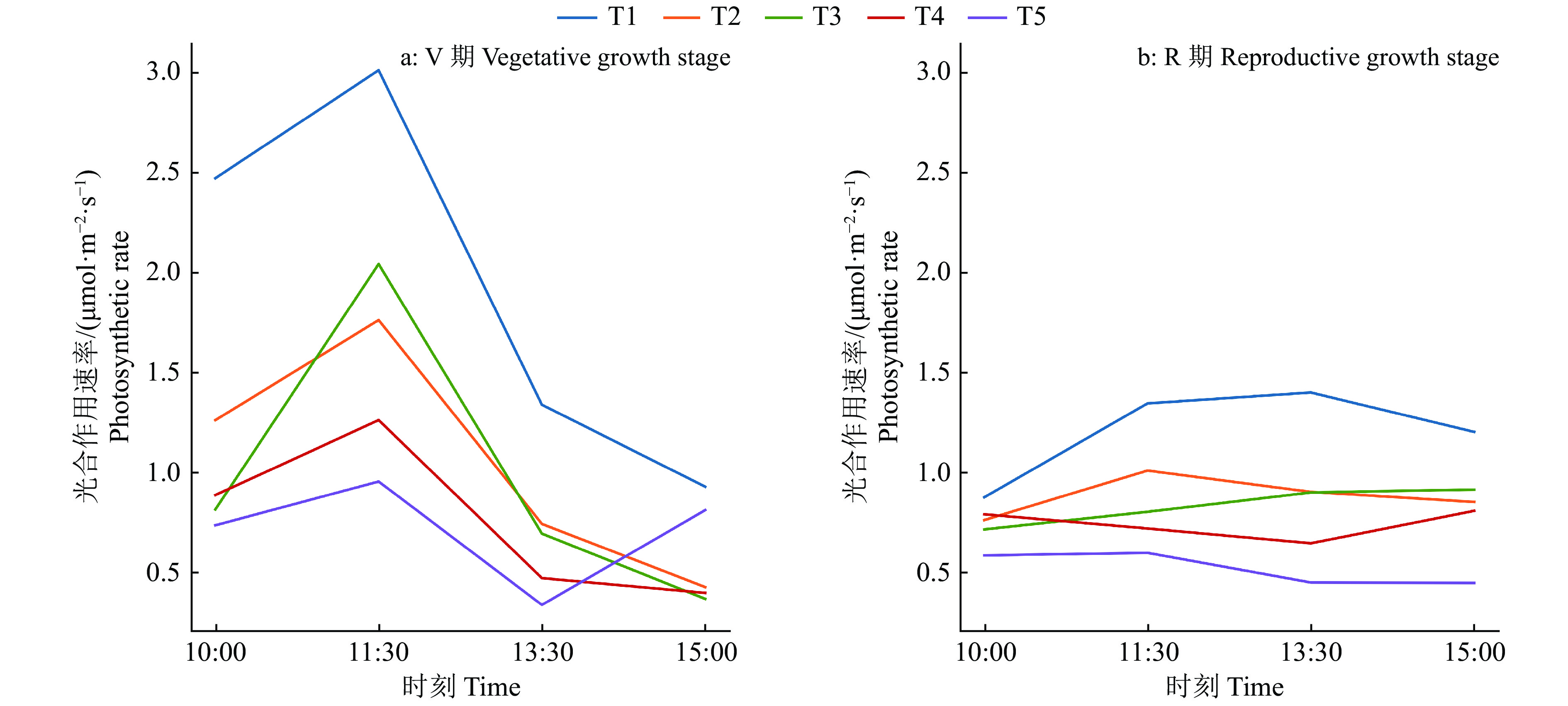
图 5 不同水分胁迫处理的油青菜心光合作用速率日变化
T1:田间持水量为32.2%;T2~T5:田间持水量分别为T1的85%、70%、55%和40%
Figure 5. Diurnal variations of photosynthetic rate of Brassica chinensis in different water stress treatments
T1: Field water holding capacity was 32.2%; T2~T5: Field water holding capacity was 85%, 70%, 55% and 40% of T1 respectively
![]()
图 6 4种模型的光合作用速率测量值与预测值的散点图
Figure 6. The scatter plots of predicted and measured photosynthetic rates based on four models
表 1 不同冠气温差上限的水分胁迫指数(CWSI)误差分析
Table 1 Error statistics of crop water stress index (CWSI) under different upper limits of canopy and air temperature difference
生长期
Stage处理1)
Treatment固定温度/℃
Fixed temperatureCWSI 平均值
Mean与无蒸腾基线的平均值差值2)
Difference with average of non-transpiration-baseline均方根误差
RMSEV T1 2.0 0.129 −0.019 0.042 3.4 0.103 −0.002 0.008 5.0 0.084 0.011 0.027 T5 2.0 0.801 −0.171* 0.179 3.4 0.635 −0.005* 0.029 5.0 0.515 0.115* 0.125 R T1 2.0 0.066 −0.025 0.066 4.2 0.043 −0.002 0.007 5.0 0.038 0.003 0.010 T5 2.0 0.908 −0.375* 0.402 4.2 0.555 −0.021* 0.033 5.0 0.487 0.047* 0.058 1) T1:田间持水量为32.2%, T5:田间持水量为T1的40%;2) “*”表示T5处理下各固定温度的CWSI平均值与无蒸腾基线的CWSI平均值差值达0.05的显著水平(LSD法)
1) T1: Field water holding capacity was 32.2%, T5: Field water holding capacity was 40% of T1; 2) “*” indicates the significant difference at 0.05 level between CWSI average values of each fixed temperature and non-transpiration-baseline in T5 treatment (Least significant difference method) 下载: 导出CSV
下载: 导出CSV
表 2 4种预测模型的误差分析
Table 2 Error statistics of four predicted models
模型
Model决定系数
R2平均绝对误差
MAE均方根误差
RMSEKNN 0.873 0.226 0.294 SVM 0.877 0.234 0.289 XGBoost 0.887 0.227 0.279 RF 0.863 0.239 0.310
下载: 导出CSV
-
[1] 卢宇鹏, 夏岩石, 温少波, 等. 不同熟性菜心品质性状的多样性分析[J]. 广东农业科学, 2020, 47(5): 29-36. [2] 叶红霞. 菜心的栽培季节和栽培方式[J]. 新农村, 2020(10): 26-27. doi: 10.3969/j.issn.1674-8409.2020.10.012 [3] 杨树涛, 黄永文, 刘泳涛, 等. 普宁市菜心生产特色农业气象指标研究[J]. 河南农业, 2018(11): 17-20. [4] 徐燕. 土壤水分胁迫对菜心生理生化指标及气孔发育的影响[D]. 广州: 暨南大学, 2010. [5] IDSO S B, JACKSON R D, PINTER P J, et al. Normalizing the stress-degree-day parameter for environmental variability[J]. Agricultural Meteorology, 1981, 24(1): 45-55.
[6] JACKSON R D, IDSO S B, REGINATO R J, et al. Canopy temperature as a crop water stress indicator[J]. Water Resources Research, 1981, 17(4): 1133-1138. doi: 10.1029/WR017i004p01133
[7] JONE H G. Use of infrared thermometry for estimation of stomatal conductance as a possible aid to irrigation scheduling[J]. Agricultural and Forest Meteorology, 1999, 95(3): 139-149. doi: 10.1016/S0168-1923(99)00030-1
[8] RUD R, COHEN Y, ALCHANATIS V, et al. Crop water stress index derived from multi-year ground and aerial thermal images as an indicator of potato water status[J]. Precision Agriculture, 2014, 15(3): 273-289. doi: 10.1007/s11119-014-9351-z
[9] 崔晓, 许利霞, 袁国富, 等. 基于冠层温度的夏玉米水分胁迫指数模型的试验研究[J]. 农业工程学报, 2005, 25(8): 22-24. doi: 10.3321/j.issn:1002-6819.2005.08.005 [10] 赵福年, 王瑞君, 张虹, 等. 基于冠气温差的作物水分胁迫指数经验模型研究进展[J]. 干旱气象, 2012, 30(4): 522-528. [11] JONES H G, STOLL M, SANTOS T, et al. Use of infrared thermography for monitoring stomatal closure in the field: Application to grapevine[J]. Journal of Experimental Botany, 2002, 53(378): 2249-2260. doi: 10.1093/jxb/erf083
[12] BIAN J, ZHANG Z T, CHEN J Y, et al. Simplified evaluation of cotton water stress using high resolution unmanned aerial vehicle thermal imagery[J]. Remote Sensing, 2019, 11(3): 267. doi: 10.3390/rs11030267.
[13] 王卫星, 罗锡文, 区颖刚, 等. 基于冠层温度的菜心缺水指数模型初步试验研究(英文)[J]. 农业工程学报, 2003, 19(5): 47-50. doi: 10.3321/j.issn:1002-6819.2003.05.010 [14] 孙道宗, 王卫星, 唐劲驰, 等. 茶树水分胁迫建模及试验[J]. 排灌机械工程学报, 2017, 35(1): 65-70. doi: 10.3969/j.issn.1674-8530.15.0249 [15] 张立元, 牛亚晓, 韩文霆, 等. 大田玉米水分胁迫指数经验模型建立方法[J]. 农业机械学报, 2018, 49(5): 233-239. doi: 10.6041/j.issn.1000-1298.2018.05.027 [16] AGAM N, COHEN Y, ALCHANATIS V, et al. How sensitive is the CWSI to changes in solar radiation?[J]. International Journal of Remote Sensing, 2013, 34(17): 6109-6120. doi: 10.1080/01431161.2013.793873
[17] KUMAR N, ADELOYE A J, SHANKAR V, et al. Neural computing modelling of the crop water stress index[J/OL]. Agricultural Water Management, 2020, 239: 1-10. [2021-01-05]. https://doi.org/10.1016/j.agwat.2020.106259.
[18] KHORSANDI A, HEMMAT A, MIREEI S A, et al. Plant temperature-based indices using infrared thermography for detecting water status in sesame under greenhouse conditions[J]. Agricultural Water Management, 2018, 204: 222-233. doi: 10.1016/j.agwat.2018.04.012
[19] 屈振江, 郑小华, 王景红, 等. 渭北旱塬苹果园内外温度变化特征研究[J]. 干旱区地理, 2016, 39(2): 301-308. [20] 吐尔逊·买买提, 谢建华. 基于DBSCAN的农机作业轨迹聚类研究[J]. 农机化研究, 2017, 39(4): 7-11. doi: 10.3969/j.issn.1003-188X.2017.04.002 [21] 谢慧婷. 基于红外热成像技术的生菜缺水指标的研究[D]. 福州: 福建农林大学, 2016. [22] MATESE A, BARALDI R, BERTON A, et al. Estimation of water stress in grapevines using proximal and remote sensing methods[J]. Remote Sensing, 2018, 10(1): 114. doi: 10.3390/rs10010114.
[23] ZHANG L Y, NIUY X, ZHANG H H, et al. Maize canopy temperature extracted from UAV thermal and RGB imagery and its application in water stress monitoring[J]. Frontiers in Plant Science, 2019, 10: 1270. doi: 10.3389/fpls.2019.01270.
[24] SONG X Y, ZHOU G S, HE Q J, et al. Stomatal limitations to photosynthesis and their critical water conditions in different growth stages of maize under water stress[J/OL]. Agricultural Water Management, 2020, 241: 1-12. [2021-01-05]. https://doi.org/10.1016/j.agwat.2020.106330.
[25] 刘煦. 林下参种植光环境的动态预测与评价研究[D]. 长春: 吉林大学, 2014. [26] 陈硕博. 无人机多光谱遥感反演棉花光合参数与水分的模型研究[D]. 杨凌: 西北农林科技大学, 2019. [27] 陈俊英, 陈硕博, 张智韬, 等. 无人机多光谱遥感反演花蕾期棉花光合参数研究[J]. 农业机械学报, 2018, 49(10): 230-239. doi: 10.6041/j.issn.1000-1298.2018.10.026 [28] 宋飞扬, 铁治欣, 黄泽华, 等. 基于KNN-LSTM的PM_(2.5)浓度预测模型[J]. 计算机系统应用, 2020, 29(7): 193-198. [29] BOTULA Y D, NEMES A, MAFUKA P, et al. Prediction of water retention of soils from the humid tropics by the nonparametric k-nearest neighbor approach[J]. Vadose Zone Journal, 2013, 12(2). doi: 10.2136/vzj2012.0123.
[30] MOHAMMADI B, MEHDIZADEH S. Modeling daily reference evapotranspiration via a novel approach based on support vector regression coupled with whale optimization algorithm[J/OL]. Agricultural Water Management, 2020, 237: 1-13. [2020-01-03]. https://doi.org/10.1016/j.agwat.2020.106145.
[31] WANG T T, YANG C H, LIANG J. Soil pH value prediction using UWB radar echoes based on XGBoost[C]//CSPS. International Conference in Communications, Signal Processing, and Systems. Urumqi: CSPS, 2019: 1941-1947.
[32] 白婷, 丁建丽, 王敬哲. 基于机器学习算法的土壤有机质质量比估算[J]. 排灌机械工程学报, 2020, 38(8): 829-834. [33] 罗清元, 杨丹, 刘丽娜, 等. 基于不同环境下的河南省典型区域土壤田间持水量研究[J]. 节水灌溉, 2019, 4929(6): 35-38. doi: 10.3969/j.issn.1007-4929.2019.06.009 [34] MARTIN E, KRIEGEL H P, SANDER J, et al. A density-based algorithm for discovering clusters in large spatial databases with noise[C]//AAAI. Proceedings of the Second International Conference on Knowledge Discovery and Data Mining (KDD-96). California: AAAI, 1996: 226-231.
[35] ALTMAN N S. An introduction to kernel and nearest-neighbor nonparametric regression[J]. The American Statistician, 1992, 46(3): 175-185.
[36] CORTES C, VAPNIK V. Support-vector networks[J]. Machine Learning, 1995, 20(3): 273-297.
[37] CHEN T Q, GUESTRIN C. XGBoost: A scalable tree boosting system[C]//ACM. Proceedings of the 22nd ACM SIGKDD International Conference on Knowledge Discovery and Data Mining. California: ACM, 2016: 785-794.
[38] BREIMAN L. Random forests[J]. Machine Learning, 2001, 45(1): 5-32. doi: 10.1023/A:1010933404324