Development of a molecular marker for sex identification of Spodoptera frugiperda
-
摘要:目的
开发用于草地贪夜蛾Spodoptera frugiperda性别鉴定的分子标记引物。
方法依据草地贪夜蛾雌、雄虫性信息素结合蛋白(Pheromone-binding protein, PBP)基因片段序列的差异设计特异性性别鉴定引物;利用引物对经过形态学鉴定的草地贪夜蛾雌、雄样本进行PCR验证。
结果2对引物分别对对应的雌、雄虫样本均扩增出450 bp左右的特异性条带。
结论开发的分子标记引物可以快速有效地对各个时期的草地贪夜蛾进行性别鉴定,为研究草地贪夜蛾雌、雄个体差异提供一种有效工具。为制定基于性别鉴定的草地贪夜蛾的有效治理防控策略提供参考。
Abstract:ObjectiveTo develop molecular marker primers for sexual identification of Spodoptera frugiperda.
MethodBased on the difference in pheromone-binding protein (PBP) gene sequences of male and female S. frugiperda, the primers were designed for distinguishing sexual distinction specifically. Using the primers, the male and female S. frugiperda samples already identified by morphology were further verified by PCR.
ResultSpecific bands of about 450 bp for the corresponding female and male samples were amplified by two pairs of primers respectively.
ConclusionThe molecular marker primers can be used to discriminate the sex of S. frugiperda in various developmental stages efficiently, and provide an effective tool for the study of the individual distinction between different sexual insects. This study provide a reference for formulating the strategy to control S. frugiperda effectively based on sexual distinction.
-
珠江是中国境内第3长河流,全长2 400 km,为中国第2大年流量的河流,为西江、北江、东江和珠江三角洲诸河的总称。其干流西江发源于云南省东北部马雄山,流经云南、贵州、广西、广东4省(区)及香港、澳门特别行政区,北江和东江水系几乎全部在广东境内。珠江流域在中国境内面积44.21万km2,另有1.1万余km2在越南境内。颜素珠等[1-2]于20世纪80年代末就开展了广东省珠江流域水生植物资源调查,共调查到水生维管植物131种,隶属44科81属,其中沉水植物33种,挺水植物70种,浮水植物28种。据国家林业局统计,我国沉水植物约有49种2变种,隶属11科15属[3]。北江地处珠江上游地区,为珠江三大水系之一,流域面积广阔,但有关水生植物资源的研究报道很少。本项目组(藉欧盟国际合作项目“河流上游水域资源的保护与可持续利用”)在广东省韶关市北江流域主要河段开展了沉水植物群落物种多样性及保护现状调查研究。本研究结果可为广东省韶关市及其邻近地区沉水植物资源的保护提供参考。
1. 研究地概况
北江为珠江水系支流,发源于江西省信丰县石碣大茅山,流入广东省南雄县境内称浈江(又名浈水),至韶关武江汇合后始称北江,此以下为中游,流至清远白庙后为下游,在佛山三水思贤窖与西江汇合注入珠江三角洲河网区流入南海,主要支流有武江、连江、滃江、潖江、滨江、绥江等。北江干流长468 km,集水面积86 710 km2,广东省境内为42 879 km2,纵贯韶关、清远、佛山和广州,占粤境内珠江流域面积的38%。韶关市为广东省境内北江源头流域,面积18 463 km2,年平均气温19.20 ℃,极高温42 ℃,极低温-4 ℃,年平均降水量1 200~1 600 mm。
2. 研究方法
2.1 野外调查方法
在踏查的基础上,在广东省韶关市沉水植物群落较集中的地段设置样带进行物种调查。每隔2 m设置1个面积为1 m×1 m的小样方,记录样方中的沉水植物种类、高度、盖度和水深。5个点分别位于北江浈水水域仁化县周田(SG1)、曲江区北江水域曲江区大坑口(SG2)、浈江区武江水域犁市(SG3)、乳源县南水(SG4和SG5)。
2.2 重要值和物种多样性
重要值(Ⅳ)=相对高度+相对盖度+相对频度。其中:相对高度=(某一物种高度/全部物种高度)×100%,相对盖度=(某个物种的盖度/所有物种的总盖度)×100%,相对频度=(某一物种的频度/所有物种的频度之和)×100%。
物种多样性包括丰富度(S)、多样性指数(H)和均匀度指数(E)。其中:S=物种数, H=-∑PilnPi, E=H/lnS。重要值能较综合地衡量其多样性,选用Ⅳ进行Pi计算,即Pi=Ⅳi/Ⅳ。
2.3 水样采集与检测
用水流测速计(南京圣荣仪器设备有限公司)分别在有草区和无草区测量水流速,每隔30 s读数1次,连续测量3 min,同时取水样3份,水的透明度使用透明度仪测量。
水样检测方法[4]:水样的pH测试使用玻璃电极法;化学需氧量(CODCr)使用重铬酸钾法;NH4+-N使用纳氏试剂分光光度法;总氮(TN)使用过硫酸钾氧化-紫外分光光度法;总磷(TP)使用过硫酸钾氧化-钼锑抗分光光度法;铜(Cu)、锌(Zn)使用火焰原子吸收分光光度法;铅(Pb)、镉(Cd)、镍(Ni)使用石墨炉原子吸收分光光度法;六价铬(Cr)使用二苯碳酰二肼分光光度法。
2.4 数据分析
将调查样方与其沉水植物重要值建立数据库。应用SPSS 19.0软件中的系统聚类分析法(Hierarchical cluster)进行样点物种相似性的聚类分析,不同样点物种相似性的差异采用离差平方和法(Ward法)[5]进行计算。
3. 结果与分析
3.1 调查区的沉水植物种类、分布及重要值
北江流域的沉水植物群落呈零星分布,仅见于人为活动较少的河岸边或河中央小岛周围,在水面交通和挖沙较频繁的区域少见。分布区域的水深约0.1~1.3 m,但在水体透明度较高的水域,沉水植物可生长在水深2~3 m的区域。在韶关市属县市5个调查区域的水域中,共记录了12种沉水植物,隶属于6科7属,包括11种被子植物和1种轮藻科植物(表 1)。在5个调查样点中,以眼子菜科植物种类最丰富,水鳖科和茨藻科次之。其中,水鳖科黑藻Hydrilla verticillata的出现频率最高,穗状狐尾藻Myriophyllum spicatum和菹草Potamogeton crispus次之,密刺苦草Vallisneria denseserrulata普遍见于韶关各水域(表 1)。
表 1 调查样点的沉水植物种类及分布Table 1. The species and distribution of submerged plants at survey sites
物种丰富度和多样性以韶关市浈江区武江SG3、乳源县南水SG4和SG5群落最高,共9种;韶关仁化浈水SG1群落的种类最贫乏,为密刺苦草单优群落,仅偶见少量的菹草。其中,微齿眼子菜Potamogeton maackianus(黄川腾等,201108002,CANT),为广东省新记录。
重要值分析结果表明:北江流域沉水植物群落以竹叶眼子菜P.wrightii、黑藻和密刺苦草为优势,以竹叶眼子菜的重要值最大,黑藻和密刺苦草次之,金鱼藻Ceratophyllum demersum最小(表 2)。
表 2 调查样点沉水植物的重要值Table 2. Importance value of submerged plants at survey sites
3.2 调查区沉水植物的地理成分分析
在所记录的6个沉水植物科中,除水鳖科为泛热带分布外,其余的科均为世界分布型;在所记录的6个沉水植物属中,除黑藻属为旧世界分布和苦草属为泛热带分布外,其余的金鱼藻属和大茨藻属2属均为世界分布。
从种的全球分布来看,竹叶眼子菜、密刺苦草、微齿眼子菜、眼子菜Potamogeton distinctus和小茨藻Najas minor 5种为亚洲特有种,其余种类均为跨洲分布(表 3)。其中,金鱼藻和大茨藻均为世界分布,但在调查区极少见;小眼子菜P.pusillus的全球分布也很广泛,除澳洲以外的大洲均有分布,但在本调查区中的分布面积较小;菹草的全球分布也较广,见于非洲以外的各大洲,在北江流域有分布,但种群通常较小;除了微齿眼子菜外,亚洲分布的眼子菜、密刺苦草和竹叶眼子均较常见,为北江流域的优势种(表 3)。
表 3 北江流域沉水植物种的地理成分1)Table 3. The geographical composition of submerged plant species in Beijiang River valley
3.3 调查区沉水植物的物候特点
实地调查发现,北江流域的沉水植物均为1年生草本植物,冬季至早春为衰亡期,花期或果期在夏季至秋季,眼子菜科植物和穗状狐尾藻通常是花果同期,多集中在7—11月(表 4)。
表 4 沉水植物的物候观察1)Table 4. The phenological observation of submerged plants
3.4 调查区沉水植物的物种多样性比较
物种多样性分析结果(表 5)表明,北江流域调查样点的沉水植物,除仁化县浈水水域SG1仅1物种外,其余4水域物种丰富度为5~10,多样性指数为1.34~1.96,均匀度指数为0.69~0.87,以韶关南水水域SG4和SG5的沉水植物群落的物种最丰富,物种多样性也高,浈水水域的物种丰富度和多样性最低。
表 5 各样点的沉水植物丰富度及多样性Table 5. Species richness and diversity of the submerged plants at different survey sites
3.5 调查区各样点的沉水植物群落的物种相似性
聚类分析结果(图 1)也表明,除单优群落外,地理位置相近的植物群落的物种相似性较高,如乳源南水水域的2个样点(SG4和SG5)物种相似性较高,优先聚在一起;曲江区大坑口(SG2)与武江水域(SG3)的沉水植物群落相似性也较高,它们先聚在一起;浈江区水域(SG1)物种最贫乏,最后才与其他样点聚在一起。
![图 1 基于沉水植物重要值的群落相似性的聚类分析]() 图 1 基于沉水植物重要值的群落相似性的聚类分析Figure 1. Clustering analysis on the community similarities based on importance value of submerged plants
图 1 基于沉水植物重要值的群落相似性的聚类分析Figure 1. Clustering analysis on the community similarities based on importance value of submerged plants3.6 调查区沉水植物群落区的水流及水质特性
5个调查区的水草区和非水草区的水质特性调查结果(表 6~表 8)表明,除了SG4水域为人工硬质护岸,水流较快外,其余水域均为自然的土质护岩,水草生境的水流较慢。水草通常生长在水位小于1.1 m、水流较缓、可透光的区域。除SG1外,冬季和夏季的水位稍有变化:SG4和SG5以夏季水位较高, SG2和SG5的水位冬季较高(表 6)。
表 6 不同季节水草群落水流速度、水位和透明度Table 6. Comparisons of water speed, height, and transparence of the survey sites in different seasons 表 7 调查群落水域的水质比较Table 7. Comparison of water quality of the survey sites
表 7 调查群落水域的水质比较Table 7. Comparison of water quality of the survey sites 表 8 调查群落水域的水质微量元素比较Table 8. Comparisons of water trace elements of the survey sites
表 8 调查群落水域的水质微量元素比较Table 8. Comparisons of water trace elements of the survey sites
不同水样的pH、总氮(TN)、总磷(TP)、铵氮(NH4+-N)含量和化学需氧量(CODCr)见表 7。不同调查点的水质有所不同,同一调查点夏季与冬季的水质也有不同程度的变化。各调查点水样呈中性,pH 6.93~7.61,以大坑口样点SG2的pH最低,南水水域的pH稍高,不同季节间的变化趋势不明显。变化较大的指标为TN、TP、NH4+-N含量和CODCr。整体来看,南水2个调查点SG4和SG5的水质最好,除总氮和铵氮外,其余指标均达到或接近国家Ⅰ级地表水的质量水平(国家地表水环境标准GB3828-2002);其余调查点水质的总氮和化学需氧量处于国家地表水Ⅱ~Ⅲ级之间。从同一水域不同季节的变化情况来看,TP、NH4+-N含量和CODCr以冬季较高,TN则以夏季稍高。
所有样本的重金属含量均达到国家Ⅰ级水的标准。在冬季,多数重金属元素含量高于夏季,但也有例外(表 8)。
4. 讨论
4.1 北江沉水植物的资源保护及其影响因素
在5个样点中,共记录了沉水植物12种,包括11种被子植物和1种轮藻。其中,微齿眼子菜为广东省首次记录。北江流域的沉水植物以世界广布型地理成分为主。其原因一方面是河流的水生环境比较稳定,缺少天然屏障,有利于沉水植物的扩散;另一方面也可能与人为干扰和环境退化导致沉水植物群落物种日益衰退甚至消失有关。
沉水植物完全依赖水生环境,对环境胁迫的反应最敏感[6]。城市化与工业化导致的河流水体受污染和采沙筑坝等人为活动是河流生物多样性发展的主要威胁[7-11]。而水体总磷、正磷酸盐、化学需氧量、透明度和水深等环境因子均与沉水植物群落分布密切相关[12-15]。许振成等[16]、吕文英等[17]检测分析了北江中上游底泥的重金属污染情况,揭示了本研究所涉及的几条北江支流(南水、武江、浈水和大坑口)底泥均存在不同程度的镉污染,以大坑口水域的潜在生态危害指数最高(1374.3),反映了其潜在生态危害已达到很强的程度;浈江水域的潜在生态危害指数次之(529.8),其潜在生态危害程度为强;南水和武江水域的潜在生态危害指数分别为293.5和290,潜在生态危害强度中等。杨晓云等[18]揭示了北江底栖动物也表现出与底泥相似的重金属污染。本研究结果揭示了曲江区北江水域的大坑口(SG2)和仁化浈水水域的周田(SG1)水域水质较差,总磷、总氮和CODCr均较高,重金属含量也明显高于南水区,反映了这些水域的水质受到一定的农业和工业污染,再加上周边洗矿的污染,导致其沉水植物物种丰富度和多样性较低。与大坑口、周田和犁市相比,南水(SG4和SG5)水域的水体很清澈,主要污染源来源于生活污水,除了总氮外,水质均达到Ⅰ级水的质量标准,其沉水植物丰富度和多样性较高,反映了水质较好的水域有利于沉水植物多样性的发展。武江犁市(SG3)沉水植物群落的干扰主要来自挖沙和大坝提升了水位(物理因素的干扰),其总磷低于大坑口和周田,其沉水植物群落的物种丰富度和多样性均居中等水平。由此可见,北江的水质污染、挖沙和大坝均影响着当地沉水植物多样性的发展,是造成北江上游水生生态资源退化的主要原因,而水质污染的影响更严重。
王书航等[19]研究了蠡湖水体氮、磷时空变化及差异性,发现蠡湖水体中氮、磷浓度不稳定,TN和TP含量呈现自东向西依次递减的趋势,在季节上,TN和TP含量呈现夏季、秋季较高,而冬季、春季低的特点。本研究结果显示, 总氮也具有夏季高于冬季的趋势,但TP、NH4+-N含量及CODCr、多数重金属含量则呈现冬季高于夏季的趋势。王书航等[19]认为蠡湖水体在秋冬季出现较高总磷浓度的原因与秋季藻类大量死亡有关。而本研究区水体中的藻类相对较少,影响水体总磷和总氮的因素主要来自农业和生活污水,而冬季水流动缓慢,是导致冬季水中磷浓度高于夏季的主要原因。
4.2 北江流域的物种相似性及与邻近水域的比较
曾宪锋[20]在粤东地区记录了金鱼藻、穗状狐尾藻和菹草等6种沉水植物种类,它们在北江流域均有出现,相似度较高。与长江流域的武汉东湖[21]和广西钦州市[22]比较,北江流域沉水植物的种类也比较接近,即均由适应性较强、分布较广的物种组成。北江流域沉水植物群落均为泛化种的原因:一方面是由水域环境自然条件差异较小的特性所决定,另一方面也与长期的人为活动,对自然水域环境干扰频繁,导致一些特化或适应性窄的物种衰退或消失有关。
4.3 加强北江流域沉水植物保护的建议
水生植物(特别是沉水植物)依赖水环境,它们不仅可为鱼类等各种水生动物提供良好的栖息环境,而且还可为其提供食物,而鱼类等水生动物也可通过与沉水植物的相互作用调节沉水植物的种群结构。此外,水生植物对修复和改善水环境也有重要作用[23-24]。因此,保护沉水植物群落及其物种多样性对维持江河流域水生资源多样性及渔业资源的可持续发展具有重要的意义。为了促进北江渔业的可持续发展,韶关市每年举行“休渔放生节”。休渔、放生等活动对北江上游渔业资源的维持有一定的作用,但对水生植物群落的保护也应该受到重视。基于本研究结果,我们对于北江流域水生植物资源的保护提出以下几点建议:
1) 加强合理规划,减少对自然水域的人为干扰,为北江上游水生生物资源的可持续发展提供良好的生态环境。近年北江上游已建立了多个大型水电站,对当地自然资源利用和减灾防灾具有重要的战略意义,但过多的水电站大坝导致河流潮汐和水量的剧烈变化,会直接或间接地影响水生生物种群的自然繁衍。因此,水电站的建设必须综合考虑其对周边水生生物资源的影响,尽可能地避免或减缓水电站建设工程对当地水生资源的破坏。此外,北江上游的挖沙活动较频繁,挖沙不仅会影响河床的地质结构,而且对水生植物及其他底栖类动物也有严重的干扰,建议将一些水生生物资源较丰富的重要区域列为禁止挖沙区。根据韶关市“十二五”林业发展规划,韶关市将在2011—2015年期间拟新建“乐昌峡省级湿地公园”、“武江区龙湾省级湿地公园”等10个湿地公园,这些湿地公园的建立将有利于北江上游地区水生资源的有效保护和可持续利用。
2) 减少污染也是保护北江水生资源的迫切需要。北江上游有大宝山铁矿、韶关冶炼厂、韶关钢铁厂等多个大型企业和多个小型私有工矿企业。大多数大型国企在加强对污染物的处理,禁止工业废水、废渣往河道排放均已做了大量设备更新和改进,但一些小型私有企业还存在着违规排放污染物的现象,极需严格监管。
3) 水生生物的生态学研究和生态学观念的宣传教育对北江上游水生资源的保护与发展也具有重要的意义。目前,有关北江流域水生资源的研究不多,大众对北江水生植物和水生动物的认识也不多。因此,应加强对北江上游水生生态系统,特别是沉水植物及其他水生动物资源的生态学研究,为珠江上游水生资源的保护与利用提供科学依据。
-
![]()
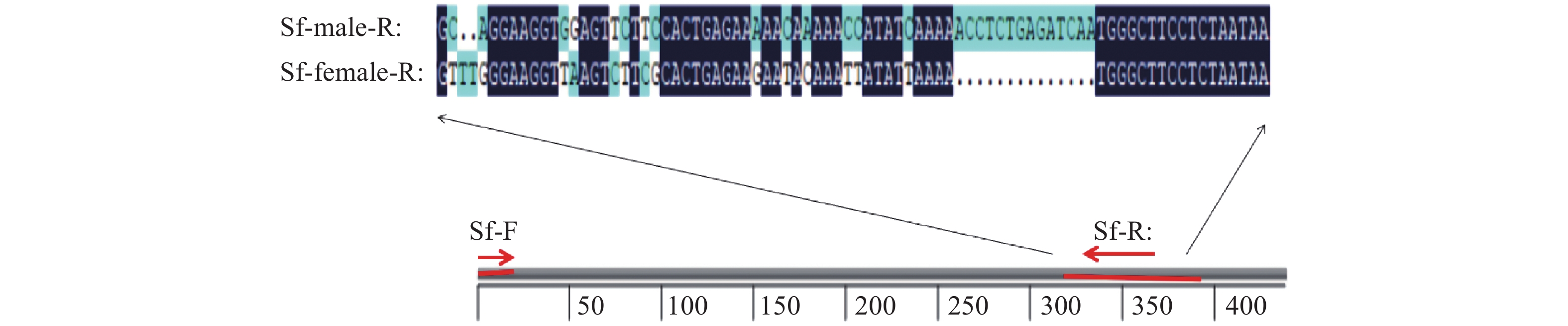
图 1 草地贪夜蛾性别鉴定引物设计
深蓝色表示相同的核苷酸序列,浅蓝色表示差异位点,黑点表示缺失位点;Sf-male-R:雄虫基因差异区段;Sf-female-R:雌虫基因差异区段;Sf-F:正向引物;Sf-R:反向引物
Figure 1. Primers design for sexual identification of Spodoptera frugiperda
Dark blue represents the same nucleotide sequence, light blue represents the differencial sites and black dots represent the missing sites; SF-male-R: Differential gene segment of male; Sf-female-R: Differential gene segment of female; Sf-F: Forward primer; Sf-R: Reverse primer
![]()
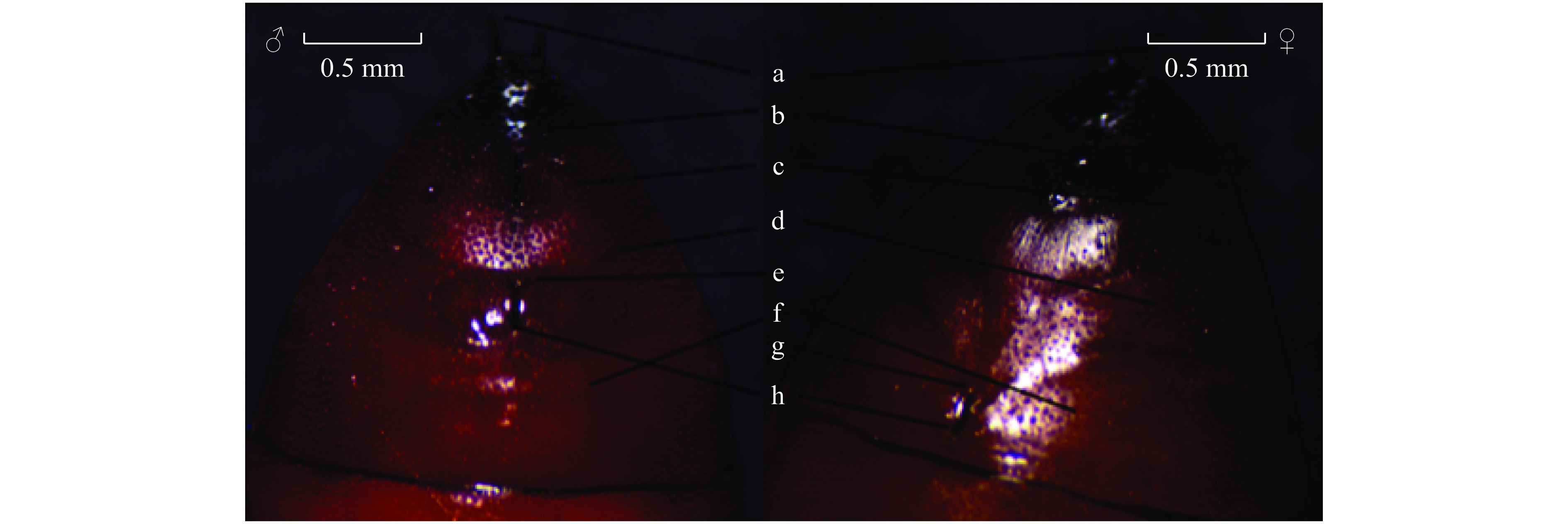
图 2 草地贪夜蛾蛹期雌、雄虫腹部末端差异对比
a:臀刺;b:肛门;c:第10腹节;d:第9腹节e:半圆形瘤状突起;f:第8腹节;g:产卵孔;h:生殖孔
Figure 2. Distinction between abdomen ends of male and female of Spodoptera frugiperda at pupal stage
a: Buttocks stab; b: Anus; c: The 10th abdominal segment; d: The 9th abdominal segment; e: Semicircular tumor-like protrusion; f: The 8th abdominal segment; g: Spawning hole; h: Genital hole
![]()
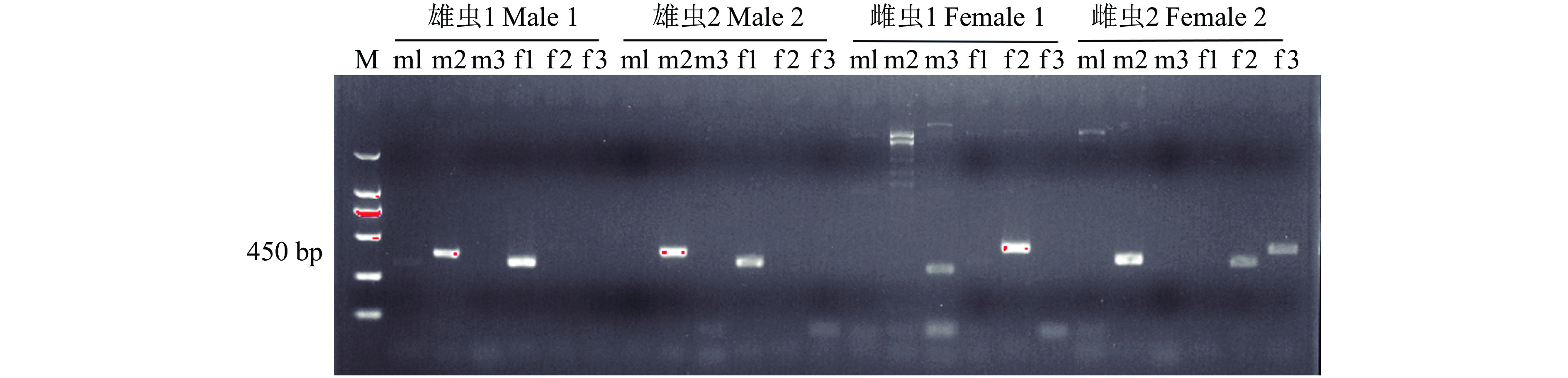
图 3 3对引物对草地贪夜蛾雌、雄虫样本的扩增
M:2000 DNA marker; m1: Sf-F/ Sf-male-R-1; m2: Sf-F/Sf-male-R-2; m3: Sf-F/Sf-male-R-3; f1: Sf-F/Sf-female-R-1; f2: Sf-F/Sf-female-R-2; f3: Sf-F/Sf-female-R-3
Figure 3. Amplification of different sexual samples of Spodoptera frugiperdaby three pairs of primers
![]()
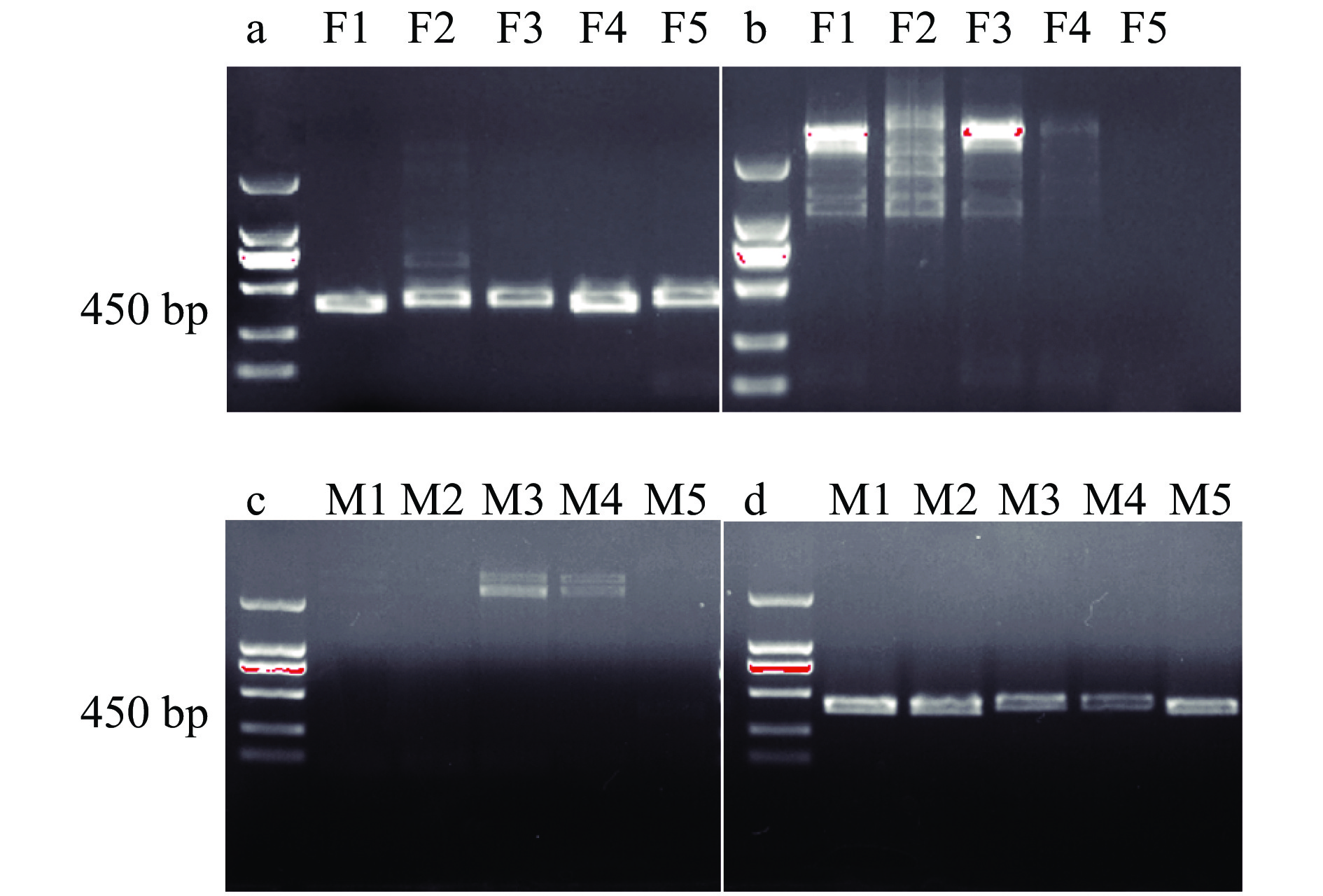
图 4 基于PCR扩增对草地贪夜蛾雌、雄虫蛹性别鉴定
F1~F5:雌虫蛹DNA;M1~M5:雄虫蛹DNA;a、c:雌虫标记引物对 Sf-F/Sf-female-R-2;b、d:雄虫标记引物对Sf- F/Sf-male-R-2
Figure 4. Sex identification of male and female pupae of Spodoptera frugiperdabased on PCR amplification
F1−F5: DNA of female pupae; M1−M5: DNA of male pupae; a and c: Pair of female marker primers of Sf-F/Sf-female-R-2; b and d: Pair of male marker primers of Sf-F/Sf-male-R-2
表 1 引物序列表
Table 1 List of primer sequence
引物名称1) Primer name 引物序列(5′→3′) Primer sequence Sf-F TAGCCGTGAGTTTGAATAGGGT Sf-female-R-1 CCTGCCAGTGCCTTATTAATTAA Sf-male-R-1 TTTTGGCAGTGCCTTATTGATTA Sf-female-R-2 CTCAGAGGTTTTTGATATGGTTT Sf-male-R-2 TGTATTCTTCTCAGTGCGAAGAC Sf-female-R-3 TTAACAACGCTCCATAATAACCT Sf-male-R-3 TAAGAACCAGTTCTTATAAACAC 1) F、R分别表示正、反向引物
1) F and R respectively represents forward and reverse primers 下载: 导出CSV
下载: 导出CSV
-
[1] TODD E L, POOLE R W. Keys and illustrations for the armyworm moths of the noctuid genus Spodoptera Guenée from the Western Hemisphere[J]. Annals of the Entomological Society of America, 1980, 73(6): 722-738. doi: 10.1093/aesa/73.6.722
[2] 姜玉英, 刘杰, 吴秋琳, 等. 我国草地贪夜蛾冬繁区和越冬区调查[J]. 植物保护, 2021, 47(1): 212-217. [3] UZAYISENGA B, WAWERU B, KAJUGA J, et al. First record of the fall armyworm, Spodoptera frugiperda(J. E. Smith, 1797) (Lepidoptera: Noctuidae), in Rwanda[J]. African Entomology, 2018, 26(1): 244-246. doi: 10.4001/003.026.0244
[4] 王磊, 陈科伟, 钟国华, 等. 重大入侵害虫草地贪夜蛾发生危害、防控研究进展及防控策略探讨[J]. 环境昆虫学报, 2019, 41(3): 479-487. [5] 林玉英, 金涛, 金启安, 等. 椰子织蛾幼虫龄数及取食量的雌雄差异[J]. 环境昆虫学报, 2017, 39(4): 912-918. [6] 覃吕高. 家蚕抗核型多角体病毒相关蛋白及雌雄差异蛋白质组学分析[D]. 镇江: 江苏大学, 2012. [7] 王华, 方妍, 王立鹏, 等. 高温对家蚕雌雄脂肪体基因表达差异的Digital gene expression(DGE)分析[C]//中国蚕学会、国家蚕桑产业技术体系. 第十二届家(柞)蚕遗传育种暨良种繁育学术研讨会论文集(摘要汇编). 中国蚕学会、国家蚕桑产业技术体系: 中国蚕学会. 2016: 41. [8] 李爽. 雌雄意大利蝗耐高温差异及其生理生化响应对策研究[D]. 乌鲁木齐: 新疆师范大学, 2016. [9] 刘振兴. 粘虫和棉铃虫趋光行为、复眼结构及光受体基因差异研究[D]. 武汉: 华中农业大学, 2019. [10] 董前进, 周金成, 朱凯辉, 等. 一种快速鉴别草地贪夜蛾蛹及成虫雌雄的简易方法[J]. 植物保护, 2019, 45(5): 96-98. [11] 冯波, 郭前爽, 王浩杰, 等. 草地贪夜蛾的准确鉴定[J]. 应用昆虫学报, 2020, 57(4): 877-888. [12] 王世英, 朱启绽, 谭煜婷, 等. 草地贪夜蛾室内人工饲料群体饲养技术[J]. 环境昆虫学报, 2019, 41(4): 742-747. [13] 刘苏, 蒋兴川, 蒋秀云, 等. 草地贪夜蛾4个性信息素结合蛋白基因的克隆及表达模式分析[J]. 环境昆虫学报, 2020, 42(3): 583-592. doi: 10.3969/j.issn.1674-0858.2020.03.9 [14] 陆雪雷, 朱诚棋, 秦文权, 等. 凤凰木夜蛾蛹和成虫的雌雄形态鉴定[J]. 中国森林病虫, 2017, 36(4): 34-36. doi: 10.3969/j.issn.1671-0886.2017.04.009 [15] 赵晓峰, 杨安頔, 张茂新, 等. 一种快速鉴别甜菜夜蛾蛹及成虫雌雄的简易方法[J]. 环境昆虫学报, 2016, 38(5): 1066-1070. [16] 张媛媛, 沈婧, 孙朝辉, 等. 桉袋蛾蛹和成虫的雌雄形态鉴定[J]. 河北林业科技, 2016(3): 8-9. doi: 10.3969/j.issn.1002-3356.2016.03.003 [17] 牛宝龙, 翁宏飚, 孟智启, 等. 棉铃虫雌性特异性分子标记的筛选及棉铃虫幼虫的性别快速鉴定[C]//浙江省人事厅、浙江省科协、浙江省蚕桑学会. “家蚕生理、病理和生物技术”学术研讨会论文选集. 浙江省人事厅、浙江省科协、浙江省蚕桑学会: 浙江省科学技术协会. 2006: 3. [18] 王慧超, 朱勇. 家蚕雌特异分子标记筛选、克隆及其序列分析[J]. 蚕业科学, 2004(1): 34-37. doi: 10.3969/j.issn.0257-4799.2004.01.008 [19] 张利娜. 长江口降海鳗鲡雌雄判别方法及雌性特异SCAR标记的建立[D]. 上海: 上海海洋大学, 2017. [20] MASARU M, TADASHI S, YOTA T, et al. Oryzias curvinotus has DMY, a gene that is required for male development in the Medaka, O. latipes[J]. Zoolog, 2003, 20(2): 159-161. doi: 10.2108/zsj.20.159
[21] 曲彬. 中国大鲵性别特异性DNA标记的鉴定及雌雄性腺差异表达基因的分析[D]. 杨凌: 西北农林科技大学, 2015. [22] 田媛. 双须骨舌鱼雌雄鉴别与性腺发育研究[D]. 上海: 上海海洋大学, 2013. [23] 田志强, 孙丽娜, 李艳艳, 等. 昆虫性信息素结合蛋白功能研究进展[J]. 农学学报, 2017, 7(9): 14-20. doi: 10.11923/j.issn.2095-4050.cjas16110038 [24] 牛小慧. 甜菜夜蛾信息素结合蛋白的表达动态及其受交配和钟基因沉默的影响[D]. 南京: 南京农业大学, 2011.