
Identification of an antagonistic strain Bacillus amyloliquefaciens E3 against Dickeya zeae and its antimicrobial activity
-
摘要:目的
纯化和鉴定从柑橘根际土壤分离得到的可抑制水稻基腐病菌Dickeya zeae EC1生长的解淀粉芽孢杆菌Bacillus amyloliquefaciens,并解析其抑菌机制。
方法采用平板扩散法从根际土壤中筛选出1株拮抗D. zeae EC1的B. amyloliquefaciens E3菌株;根据细菌菌落表型、生理生化特征结合16S rDNA序列分析等方法对E3菌株进行分类鉴定;检测无菌E3培养液对D. zeae EC1侵染水稻种子能力的影响;盐酸沉淀结合丙酮抽提法提取E3菌株脂肽粗提物;采用平板对峙法测定E3菌株脂肽粗提物对病原真菌的抑菌活性,琼脂扩散法测定E3菌株脂肽粗提物对病原细菌的抑菌活性及最小抑菌浓度(Minimum inhibitory concentration, MIC);用Durashell C18柱对脂肽粗提物进行HPLC分离纯化;通过液相色谱−质谱联用(LC-ESI-MS)分析,鉴定抑菌物质的相对分子质量并推测其化学组成。
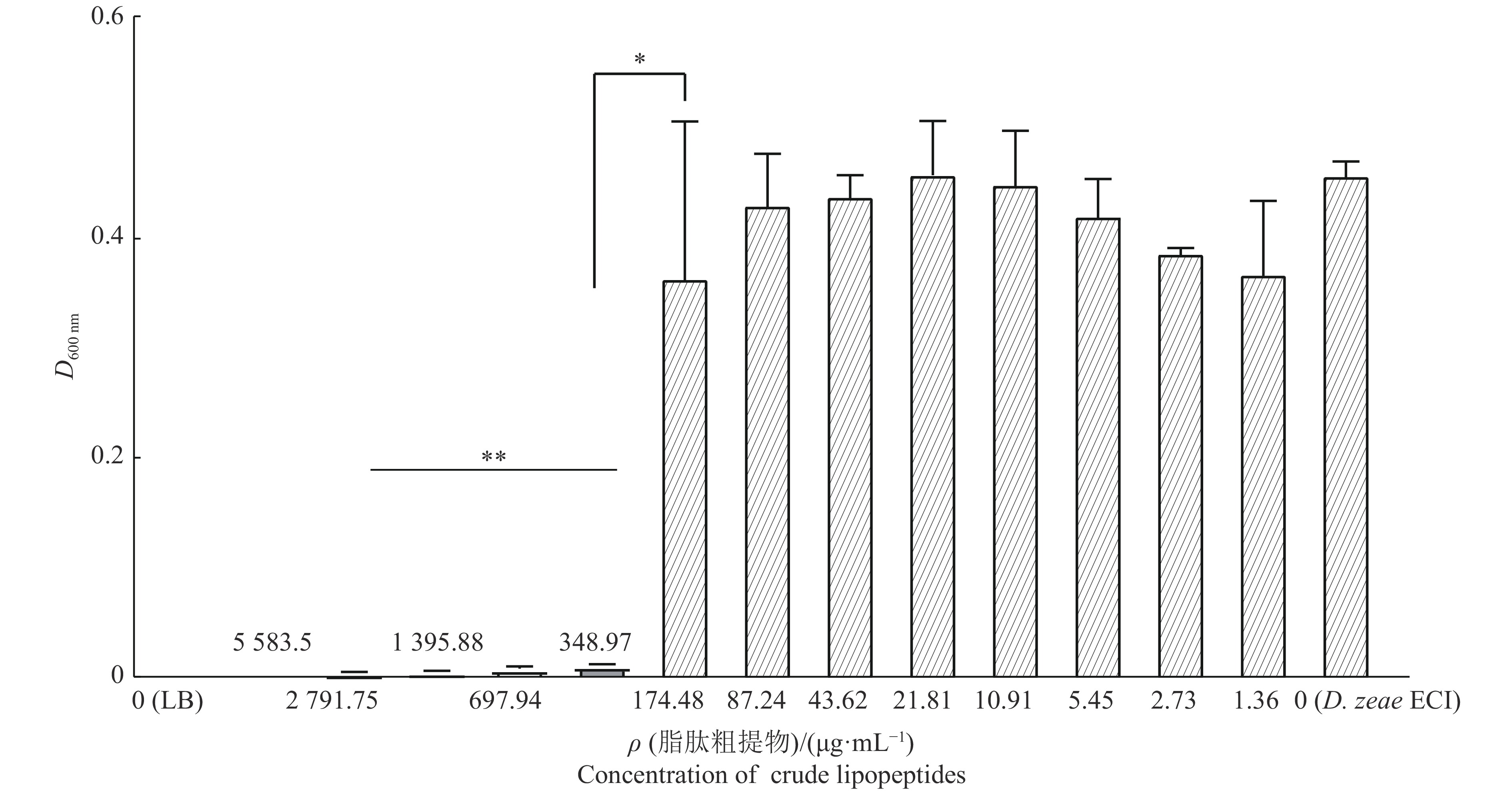
结果B. amyloliquefaciens E3菌株对茄病镰刀菌Fusarium solani、香蕉基腐病菌、茄科劳尔氏菌Ralstonia solanacearum等多种植物病原菌生长具有抑制作用,表现为广谱抗性;B. amyloliquefaciens E3菌株的无菌培养液能够抑制D. zeae EC1侵染萌发的水稻种子,显著提高水稻种子的萌芽率;B. amyloliquefaciens E3菌株的脂肽粗提物可以抑制D. zeae EC1的生长,其MIC为348.97 µg/mL;HPLC分离纯化结合LC-ESI-MS分析发现,B. amyloliquefaciens E3菌株分泌的主要抑菌活性物质包括Surfactin、Fengycin和Iturin 3类脂肽类抗生素。
结论B. amyloliquefaciens E3菌株具有作为生防菌的潜力,它可能通过分泌Surfactin、Fengycin和Iturin 3种脂肽类抑菌活性物质,拮抗水稻基腐病菌、茄科劳尔氏菌、茄病镰刀菌等多种植物病原菌。研究结果可为该菌的生物防治应用提供理论依据。
Abstract:ObjectiveTo isolate and identify Bacillus amyloliquefaciens strain which can inhibit the growth of Dickeya zeae EC1 from the rhizosphere soil of citrus plants, and analyze its antibacterial mechanism.
MethodThe B. amyloliquefaciens E3 strain antagonizing the growth of D. zeae EC1 was screened from the rhizosphere soil by plate diffusion method. The E3 strain was classified and identified based on colony phenotype, physiological and biochemical characteristics of bacteria and 16S rDNA sequence analysis. Effects of E3 sterile culture medium on the seeds germination ability of rice infected by D. zeae EC1 was detected. The crude lipopeptides of E3 strain was extracted by hydrochloric acid precipitation combined with acetone extraction. The antimicrobial activity of crude lipopeptides against pathogenic fungi were determined by plate confrontation method. The antimicrobial activity of crude lipopetides against pathogenic bacteria were determined by agar diffusion method, and the minimum inhibitory concentration (MIC) was obtained. The crude lipopeptides were separated and purified by HPLC with Durashell C18 column. Liquid chromatography-mass spectrometry (LC-ESI-MS) analysis was used to identify the molecular weights of the antimicrobial substances and speculate its chemical composition.
ResultB. amyloliquefaciens E3 strain had an inhibitory effect against Fusarium solani, D. zeae MS1 and Ralstonia solanacearum, etc, indicating that it had broad-spectrum resistance. The sterile culture medium of B. amyloliquefaciens E3 strain inhibited the infection of germinating rice seeds by D. zeae EC1 and significantly increased the seed germination rate. The crude lipopeptides of E3 strain inhibited the growth of D. zeae EC1 with the MIC of 348.97 µg/mL. HPLC combined with LC-ESI-MS analysis showed that the main antibacterial substances secreted by E3 strain included surfactin, fengycin and iturin.
ConclusionThe B. amyloliquefaciens E3 strain is potential to be a biocontrol agent. It can antigonize a variety of plant pathogens such as D. zeae EC1, R. solanacearum and F. solani by secreting three kinds of lipopeptides including surfactin, fengycin and iturin. The results provide a theoretical basis for the application of B. amyloliquefaciens E3 in biological control.
-
中药大黄来源于蓼科植物大黄的根茎,最早记载于《神农百草经》,有药中“将军”之美称,被列为中药“四大金刚”之一,已经有
2000 多年的中医临床用药史[1]。中药大黄主要由大黄酸、大黄素、大黄酚、芦荟大黄素等游离蒽醌,番泻苷A/B等二蒽酮苷类结合型蒽醌,二苯乙烯,多糖,单宁等成分组成,具有泻下攻积、抗菌、抗感染、清热泻火、凉血解毒、逐瘀通经、保肝利胆等多种药理作用[2]。其中,蒽醌是大黄的主要活性成分,含量(w)约占其成分的3%~5%[3]。大黄酸(Rhein,RH)是大黄蒽醌的主要成分之一,结构式如图1所示,具有抗感染[4]、抗肿瘤[5]、抗糖尿病[6]、降脂[7]、抗氧化、抗菌、抗病毒、抗肾毒性等[8]多种药理作用。然而,大黄酸溶解度低、脂溶性差、毒性强、生物利用度差、易引起胃肠道不适,这些特性限制了其临床应用[9]。近年来,畜禽病原菌的抗生素耐药性逐渐变得严重,导致抗生素敏感性下降,研发抗生素增效剂、复方制剂等的防控策略逐渐成为应对畜禽耐药病原菌的研究热点。大黄酸对幽门螺杆菌、金黄色葡萄球菌等多种厌氧菌有强大的抑菌作用。对大黄酸进行结构改造,通过羟基、羧基取代成酯或酰胺等修饰,改善大黄酸理化性质,降低毒性,提高抗感染、抗癌、抗菌等活性,具有重要意义,可为畜禽病原菌耐药防控提供新思路,也为大黄酸类衍生物的进一步研究提供参考依据。目前,大黄酸衍生物双醋瑞因是大黄酸最具代表性的衍生物之一,可阻滞白细胞介素−1β(Interleukin-1β,IL-1β)的作用,减少破骨细胞的形成和抑制吸收因子的合成在临床上广泛应用于治疗骨关节炎[10]。双醋瑞因的上市为后续大黄酸衍生物的研究和开发提供了理论依据;但是,双醋瑞因存在严重的毒副作用,为了控制其重度腹泻及影响肝脏的风险,对其临床使用设置了限制性条件。双醋瑞因又名1, 8−二乙酰基−3−羧基蒽醌,用乙酰氯对大黄酸第1、8位羟基进行保护。有研究表明,第1、8位羟基对大黄酸毒性的影响较小[11],因此,双醋瑞因对大黄酸毒性的改善并不明显。研究表明,大黄酸第3位羧基是大黄酸毒性的关键影响因素[11],对第3位羧基进行修饰能提高大黄酸的跨膜能力,从而可能提高大黄酸的抗感染效果[12]。据报道,游离的脂肪酸在肠道炎症反应中具有潜在的应用价值,它可以与肠道细胞膜的特异性受体结合从而发挥免疫保护作用,降低肠道促炎因子水平,以治疗肠道炎症[13]。一方面,脂肪酸作为能量物质,提供了成年人每日所需能量的30%[14];另一方面,脂肪酸参与生物膜的构成、受体激活、信号转导、神经系统发育和各种基因的调节[15]。脂肪酸衍生化可以使大黄酸具有更好的应用价值,保护大黄酸,避免其在体内快速代谢或降解,提高大黄酸稳定性[16];还可以增强大黄酸分子的亲脂性和跨膜转运能力[17];由于肿瘤组织对脂肪酸的吸收能力较强,也可以促进大黄酸分子对癌细胞的靶向积累。总之,通过脂肪酸对大黄酸进行衍生化具有广阔的应用前景。
本研究前期工作发现,大黄酸与脂肪酸具有协同抗感染作用,并且脂肪酸能在一定程度上降低大黄酸的毒性;但是,大黄酸和脂肪酸在体内代谢速率、生物利用度等因素影响下很难发挥协同作用。因此,本研究通过对大黄酸第3位羧基与羟基癸酸和癸醇进行酯化衍生化,制备出4种大黄酸衍生物,通过核磁共振氢谱(1H nuclear magnetic resonance,1H NMR)和傅里叶变换红外光谱(Fourier transforme infrared spectroscopy,FTIR)表征其结构,进一步探究其毒性、抗感染和抗氧化活性变化,筛选出具有应用前景的大黄酸衍生物,以期为大黄酸衍生物的结构设计及其生物活性等研究提供参考依据。
1. 材料与方法
1.1 仪器与材料
核磁共振波谱(德国Bruker公司);磁力搅拌器(上海龙跃仪器设备有限公司);流式细胞计数仪(美国贝克曼库尔特);傅里叶变换红外光谱仪(美国Thermo Fisher Nicolet公司);酶标定量测定仪(美国Beckman公司);CO2细胞培养箱(新加坡艺思高科技有限公司);RT-qPCR仪(美国Bio-Rad生命医学产品有限公司);旋转蒸发仪N-1200B(日本EYELA东京理化);小鼠单核巨噬细胞RAW 264.7(武汉普诺赛生命科技有限公司);一氧化氮(NO)检测试剂盒(上海碧云天生物技术股份有限公司);DMEM、特级胎牛血清、青链霉素(赛默飞生命科学产品与服务旗舰店);总RNA极速提取试剂盒(广州晖鼎生物科技有限公司);4−(二甲氨基)吡啶(4-Dimethylaminopyridine,DMAP)、N,N′−二环己基碳二亚胺(N, N′-Dicyclohexylcarbodiimide,DCC)(上海麦克林生化科技有限公司);氘代二甲基亚砜(Dimethyl sulfoxide,DMSO)(上海源叶生物科技有限公司);大黄酸(上海麦克林生化科技有限公司);1−癸醇(1-Decanol,DA)、10−羟基癸酸(10-Hydroxydecanoic acid,10-HA)、10−羟基−2−癸烯酸(10-Hydroxy-2-decenoic acid,10-HDA)、5−羟基癸酸钠(5-Hydroxydecanoate sodium,5-HD)(上海毕得医药科技股份有限公司)结构式如图2所示;双氯荧光黄乙酸乙酯(2, 7-Dichlorofluorescin diacetate,DCFH-DA)(广州市齐云生物技术有限公司)。
![]() 图 2 癸醇(A)和羟基癸酸(B、C、D)的结构式Figure 2. Structural formulas of decanol (A) and hydroxydecanoic acid (B, C, D)
图 2 癸醇(A)和羟基癸酸(B、C、D)的结构式Figure 2. Structural formulas of decanol (A) and hydroxydecanoic acid (B, C, D)1.2 试验与方法
1.2.1 大黄酸衍生物的制备与纯化
精密称取0.5 g(1.70 mmol)大黄酸,置于20 mL DMSO溶液中,于50 ℃、200 r/min搅拌1 h,完全溶解后,加入DA/10-HA/10-HDA/5-HD、DCC 3.40 mmol和DMAP 0.85 mmol,反应摩尔比按照大黄酸∶DA/10-HA/10-HDA/5-HD∶DCC∶DMAP=1.0∶1.0∶2.0∶0.5。在50 ℃条件下反应,每间隔1 h,用薄层层析法监测反应情况,确认产物生成,直至产物不再增加(展开剂为乙酸乙酯和石油醚,体积比为2∶3),停止反应。大黄酸与羟基脂肪酸的反应结构式如图3所示。反应结束后待温度降至室温,将反应混合物用饱和碳酸氢钠溶液透析1 d (透析袋截留相对分子质量为500),去离子水透析2 d,每隔12 h更换透析液,除去未反应的物质。随后,在−75 ℃、0.001 MPa条件下冷冻干燥48 h,得到冻干粗产物。将上述得到的粗产品用38~48 μm孔径的硅胶柱层析进行干法上样,使样品层与硅胶层之间的高度比为1∶20。洗脱液乙酸乙酯∶石油醚=2∶3(V/V),直至监测到产物完全洗脱,获得最终产物。用旋转蒸发器除去溶剂,于50 ℃真空烘箱干燥24 h,得到黄色粉末固体产物。
![]() 图 3 大黄酸与羟基脂肪酸的反应结构式F-A为接枝脂肪酸或癸醇,R为F-A反应掉羟基的部分结构,R-FA为接枝后的大黄酸衍生物。Figure 3. Structural formula for the reaction of rhein with hydroxy fatty acidsF-A is grafted fatty acid or decanol, R is the part of the structure where F-A reacts to drop the hydroxyl group, R-FA is the grafted rhein acid derivative.
图 3 大黄酸与羟基脂肪酸的反应结构式F-A为接枝脂肪酸或癸醇,R为F-A反应掉羟基的部分结构,R-FA为接枝后的大黄酸衍生物。Figure 3. Structural formula for the reaction of rhein with hydroxy fatty acidsF-A is grafted fatty acid or decanol, R is the part of the structure where F-A reacts to drop the hydroxyl group, R-FA is the grafted rhein acid derivative.1.2.2 大黄酸衍生物的抗感染活性测定
为验证大黄酸及其衍生物对RAW 264.7细胞存活率的影响,选用CCK-8试剂盒对RAW 264.7细胞存活率进行检测。分别用10~160 μmol/L的大黄酸及其衍生物处理RAW 264.7细胞24 h,10%(
$\varphi $ ) CCK-8试剂孵育0.5~4.0 h,酶标仪设定波长为450 nm,测定每孔光密度。为验证大黄酸衍生物的抗感染效果,构建脂多糖(Lipopolysaccharide,LPS)诱导的RAW 264.7细胞模型,测定大黄酸及其衍生物处理后RAW 264.7中NO释放量以及促炎因子TNF-α、IL-1β、IL-6表达量变化。用20 μmol/L的大黄酸、R-5HD、R-DA、R-10HA、R-10HDA在37 ℃、5%(
$\varphi $ ) CO2条件下预孵育8 h,加入LPS,终质量浓度为 1 μg/mL,在37 ℃、5%($\varphi $ ) CO2条件下孵育16 h(另外设置LPS阳性对照组,不加药物处理)。用细胞培养液上清液[DMEM+10% (φ) FBS溶液+1% (φ) 青链霉素溶液]稀释NaNO2标准品至0、1、2、5、10、20、40、60、100 μmol/L。按50 μL每孔,在96孔板中加入标准品及样品(每组设置3个复孔),用酶标仪于540 nm波长处测定光密度,计算NO含量。提取RAW 264.7的RNA,反转录,利用RT-qPCR测定肿瘤坏死因子−α (Tumour necrosis factor-α,TNF-α)、IL-1β、白细胞介素−6 (Interleukin-6,IL-6)的表达量变化,具体步骤如下。首先,投入总量为1 μg的RNA,2 μL 4× gDNA wiper Mix,用RNase-free ddH2O定容至10 μL,充分混匀,于42 ℃离心2 min后,得到去DNA的反应液。然后,在上述反应液中加入4 μL 5× Hiscript Ⅲ qRT SuperMix,用RNase-free ddH2O定容至20 μL,得到反转录体系,充分混匀,短暂离心,经37 ℃ 15 min、85 ℃ 5 s的PCR反应后,得到cDNA。最后,按照下述反应程序进行RT-qPCR:95 ℃ 30 s,循环1次;95 ℃ 10 s,循环40次;60 ℃ 30 s,循环40次;72 ℃ 30 s,循环40次。RT-qPCR反应体系中所需的引物序列见表1。表 1 RT-qPCR引物Table 1. RT-qPCR primer引物名称
Primer name引物序列(5′→3′)
Primer sequence扩增序列
Amplified sequenceGAPDH-F ACCCAGAAGACTGTGGATGG GAPDH mRNA GAPDH-R CACATTGGGGGTAGGAACAC GAPDH mRNA TNF-ɑ-F CTCTTCAAGGGACAAGGCTG TNF-ɑ mRNA TNF-ɑ-R CGGACTCCGCAAAGTCTAAG TNF-ɑ mRNA IL-6-F CCGGAGAGGAGACTTCACAG IL-6 mRNA IL-6-R TCCACGATTTCCCAGAGAAC IL-6 mRNA IL-1β-F GACCTTCCAGGATGAGGACA IL-1β mRNA IL-1β-R AGGCCACAGGTATTTTGTCG IL-1β mRNA 1.2.3 大黄酸衍生物的抗氧化活性测定
为验证大黄酸及其衍生物对LPS诱导的RAW 264.7细胞中活性氧(Reactive oxygen species,ROS)产生的影响,用20 μmol/L的大黄酸以及大黄酸衍生物处理RAW 264.7细胞。ROS荧光探针DCFH-DA可穿透活细胞膜进入细胞内,并可被细胞内的ROS氧化,生成氧化乙啶;氧化乙啶可嵌入染色体DNA中,产生红色荧光。根据此原理可以评估细胞内ROS含量及其变化情况。用DCFH-DA染色,通过流式细胞计数仪检测大黄酸及其衍生物对RAW 264.7细胞中ROS产生的影响,评估大黄酸及其衍生物抗氧化水平。
1.2.4 统计学分析
采用Orgin Pro 9.0和Graph Pad 8进行统计分析,试验分组均至少有3次重复,采用t检验比较组别之间的差异性。若P>0.05,则无统计学意义;若P<0.05、P<0.01或P<0.001,则其有显著差异;若P<0.000 1,则其有极显著差异。
2. 结果与分析
2.1 大黄酸衍生物结构表征
为了表征大黄酸衍生物是否成功合成,用FTIR和1H NMR表征其结构。R-5HD的FTIR表征结果如图4A所示。大黄酸的羟基(—OH)峰位于3 420.40 cm−1,羰基(—C=O)峰位于1 648.98 cm−1;5-HD的羟基峰位于3 410.34 cm−1,羰基峰位于1 703.44 cm−1;R-5HD的羟基峰位于3 447.58 cm−1,羰基峰位于1 740.69 cm−1,新生成的酯键为1 275.67 cm−1。大黄酸蒽醌母核上的氢在1H NMR上分别对应a1(7.40×10−6)、b1/c1(7.70×10−6)、d1(7.82×10−6)、e1(8.09×10−6),酚羟基氢对应h1/f1(11.87×10−6),第3位羧基(—COOH)氢对应g1(13.79×10−6);5-HD碳链上的氢对应b2(2.33×10−6)、c2(1.54×10−6)、d2(1.38×10−6)、e2(3.10×10−6)、g2(1.38×10−6)、h2/i2(1.25×10−6)、j2(1.28×10−6)、k2(0.88×10−6),羟基氢对应f2(3.41×10−6),羧基氢对应a2(11.87×10−6)。由大黄酸衍生物R-5HD的1H NMR表征(图4B)可以看出,大黄酸第3位羧基峰和5-HD碳链上的羟基峰消失,与5-HD对比,R-5HD中峰h3(4.02×10−6)发生明显的偏移。以上结果均表明R-5HD成功合成。
R-DA的FTIR表征结果如图4C所示,大黄酸的羟基峰位于3 420.40 cm−1,羰基峰位于1 648.98 cm−1;R-DA的羟基峰位于3 447.58 cm−1,羰基峰位于1 740.69 cm−1,新生成的酯键为1 275.67 cm−1。大黄酸蒽醌母核上的氢在1H NMR上分别对应a1(7.40×10−6)、b1/c1(7.70×10−6)、d1(7.82×10−6)、e1(8.09×10−6),酚羟基的氢对应h1/f1(11.87×10−6),第3位羧基的氢对应g1(13.79×10−6);DA碳链上的氢对应b2/d2/e2/f2(1.26×10−6)、g2(1.30×10−6)、h2(1.43×10−6)、i2(1.50×10−6)、j2(4.35×10−6),羟基氢对应k2(3.23×10−6)。由R-DA的1H NMR表征(图4D)可以看出,大黄酸第3位羧基峰和DA碳链上的羟基峰消失,与DA对比,R-DA中羟基相邻烷烃链上氢对应的h3(4.33×10−6)发生明显偏移。以上结果均表明R-DA成功合成。
R-10HA FTIR结构表征如图4E所示。大黄酸的羟基峰位于3 420.40 cm−1,羰基峰位于1 648.98 cm−1;10−HA的羟基峰位于3 410.34 cm−1,羰基峰位于
1703.44 cm−1;R-10HA的羟基峰位于3 447.58 cm−1,羰基峰位于1 740.69 cm−1,新生成的酯键为1 275.67 cm−1。大黄酸蒽醌母核上的氢在1H NMR上分别对应a1(7.40×10−6)、b1/c1(7.70×10−6)、d1(7.82×10−6)、e1(8.09×10−6),酚羟基的氢对应h1/f1(11.87×10−6),第3位羧基的氢对应g1(13.79×10−6);10-HA碳链上的氢对应c2(1.54×10−6)、d2(1.33×10−6)、e2/f2(1.26×10−6)、g2(1.30×10−6)、h2(1.43×10−6)、i2(1.58×10−6)、j2(3.62×10−6),羟基的氢对应k2(4.7×10−6),羧基的氢对应a2(11.87×10−6)。由R-10HA的1H NMR 表征(图4F)可以看出,大黄酸第3位羧基峰和10-HA碳链上的羟基峰消失,与10-HA对比,R-10HA中峰h3(4.02×10−6)发生明显的偏移。以上结果均表明R-10HA成功合成。R-10HDA FTIR表征结果如图4G所示,大黄酸的羟基峰位于3 420.40 cm−1,羰基峰位于1 648.98 cm−1;10-HDA的羟基峰位于3 410.34 cm−1,羰基峰位于1 703.44 cm−1;R-10HDA的羟基峰位于3 447.58 cm−1,羰基峰位于1 740.69 cm−1,新生成的酯键为1 275.67 cm−1。大黄酸蒽醌母核上的氢在1H NMR上分别对应a1(7.40×10−6)、b1/c1(7.70×10−6)、d1(7.82×10−6)、e1(8.09×10−6),酚羟基的氢对应h1/f1(11.87×10−6),第3位羧基的氢对应g1(13.79×10−6);10-HDA碳链上的氢对应b2(6.02×10−6)、c2(7.11×10−6)、d2(2.16×10−6)、e2(1.29×10−6)、f2(1.33×10−6)、g2(1.26×10−6)、h2(1.43×10−6)、i2(1.58×10−6),羟基的氢对应k2(4.70×10−6),羧基的氢对应a2(12.05×10−6)。由R-10HDA的1H NMR表征(图4H)可以看出,大黄酸第3位羧基峰和10-HDA碳链上的羟基峰消失,与10-HDA对比,R-10HDA中峰h3(4.02×10−6)发生明显偏移。以上结果均表明R-10HDA成功合成。
2.2 大黄酸衍生物的抗感染活性评估
2.2.1 大黄酸衍生物对RAW 264.7细胞的毒性
结果如图5所示,大黄酸浓度高于40 μmol/L时具有明显的毒性,RAW 264.7细胞存活率降低至60%;DA浓度为160 μmol/L时,RAW 264.7细胞存活率降低至60%。而R-DA、R-10HA、R-10HDA、R-5HD浓度为160 μmol/L时,RAW 264.7细胞存活率仍接近100%。由此可见,大黄酸衍生物显著降低大黄酸的细胞毒性。
![]() 图 5 不同浓度大黄酸及其衍生物对RAW 264.7细胞存活率的影响“*”“**”“***”分别表示相同处理浓度大黄酸衍生物组与大黄酸组在0.05、0.01和0.001水平差异显著(t检验)。Figure 5. Effect of different concentrations of rhein and its derivatives on survival rates of RAW 264.7 cells“*” “**” “***” indicate significant differences between rhein derivative group and rhein group at 0.05, 0.01 and 0.001 levels respectively in the same treatment concentration (t test).
图 5 不同浓度大黄酸及其衍生物对RAW 264.7细胞存活率的影响“*”“**”“***”分别表示相同处理浓度大黄酸衍生物组与大黄酸组在0.05、0.01和0.001水平差异显著(t检验)。Figure 5. Effect of different concentrations of rhein and its derivatives on survival rates of RAW 264.7 cells“*” “**” “***” indicate significant differences between rhein derivative group and rhein group at 0.05, 0.01 and 0.001 levels respectively in the same treatment concentration (t test).2.2.2 大黄酸衍生物的抗感染活性
大黄酸及其衍生物对NO释放量的影响如图6A所示。与空白对照组(CK)相比,阳性对照组(LPS模型组)NO释放量极显著增加(P<
0.0001 )。与LPS模型组相比,大黄酸、R-5HD、R-DA、R-10HA、R-10HDA均能够极显著抑制LPS诱导的RAW 264.7中NO的释放量,说明4种大黄酸衍生物均具有较强的抗感染活性,对第3位羧基进行结构修饰不会丧失大黄酸的抗感染活性。![]() 图 6 大黄酸及其衍生物的抗感染作用“*”“**”“***”“****”分别表示与LPS组(阳性对照组)在0.05、0.01、0.001和0.000 1水平差异显著(t检验)。Figure 6. Anti-inflammatory effects of rhein and its derivatives“*” “**” “***” “****” indicate significant differences differed from LPS group (positive control group) at 0.05, 0.01, 0.001 and 0.000 1 levels respectively (t test).
图 6 大黄酸及其衍生物的抗感染作用“*”“**”“***”“****”分别表示与LPS组(阳性对照组)在0.05、0.01、0.001和0.000 1水平差异显著(t检验)。Figure 6. Anti-inflammatory effects of rhein and its derivatives“*” “**” “***” “****” indicate significant differences differed from LPS group (positive control group) at 0.05, 0.01, 0.001 and 0.000 1 levels respectively (t test).大黄酸及其衍生物均能显著降低LPS诱导的RAW 264.7中促炎因子TNF-α、IL-1β、IL-6的mRNA相对表达量,结果如图6B~6D所示。与空白对照组(CK)相比,阳性对照组(LPS组)的TNF-α mRNA表达水平增加了47倍;与LPS组相比,大黄酸、R-5HD、R-DA、R-10HA、R-10HDA均显著降低了TNF-α mRNA表达水平(P<0.05)。LPS组的IL-1β mRNA表达水平比CK增加了8 813倍;与LPS组相比,大黄酸、R-5HD、R-DA、R-10HA、R-10HDA均显著降低了IL-1β mRNA表达水平。LPS组的IL-6表达水平比CK增加了4 885倍;与LPS组相比,大黄酸、R-5HD、R-DA、R-10HA、R-10HDA均显著降低了IL-6 mRNA表达水平。以上结果表明,大黄酸、R-5HD、R-DA、R-10HA和R-10HDA均能显著抑制炎症因子IL-1β、IL-6和TNF-α mRNA的表达水平,对大黄酸进行结构修饰使其抗感染效果更强,且R-DA的抗感染效果最强。
2.2.3 大黄酸衍生物的抗氧化活性
如图7所示,DCFH-DA探针孵育后,空白对照组(CK)的荧光强度为1 319.66;经过LPS刺激后细胞内ROS水平显著升高,荧光强度为60 968.00,比空白对照组增加了46倍(P<
0.0001 )。药物处理后大黄酸、R-5HD、R-DA、R-10HA以及R-10HDA组的荧光强度均显著下降,且R-5HD、R-DA、R-10HA、R-10HDA的抑制效果均优于大黄酸。以上结果表明,修饰后的R-5HD、R-DA、R-10HA、R-10HDA显著增强了大黄酸的抗氧化活性。![]() 图 7 大黄酸及其衍生物对LPS诱导RAW 264.7细胞释放ROS的影响“**”“***”“****”分别表示与LPS组(阳性对照组)在0.01、0.001和0.000 1水平差异显著(t检验)。Figure 7. Effect of rhein and its derivatives on LPS-induced ROS release from RAW 264.7 cells“**” “***” “****” indicate significant differences differed from LPS group (positive control group) at 0.01, 0.001 and 0.000 1 levels respectively (t test).
图 7 大黄酸及其衍生物对LPS诱导RAW 264.7细胞释放ROS的影响“**”“***”“****”分别表示与LPS组(阳性对照组)在0.01、0.001和0.000 1水平差异显著(t检验)。Figure 7. Effect of rhein and its derivatives on LPS-induced ROS release from RAW 264.7 cells“**” “***” “****” indicate significant differences differed from LPS group (positive control group) at 0.01, 0.001 and 0.000 1 levels respectively (t test).3. 结论与讨论
本研究以大黄酸为原料,以羟基癸酸和癸醇为修饰物,经一步酯化反应合成了4种大黄酸衍生物(R-5HD、R-DA、R-10HA、R-10HDA),通过FTIR以及1H NMR对合成的化合物进行结构鉴定,证实4种衍生物合成成功,经对大黄酸及其衍生物进行毒性研究以及抗感染、抗氧化活性研究,主要结论及其分析如下。
通过大黄酸及其衍生物对RAW 264.7细胞存活率影响的分析,经过羟基癸酸和癸醇修饰之后的大黄酸衍生物对RAW 264.7细胞的毒性降低,安全性增加。羟基癸酸和癸醇与药物分子的化学偶联可能会引起它们在体内的药效学/药代动力学变化,从而降低它们的毒性。经羟基癸酸和癸醇修饰之后的大黄酸衍生物具有更强的抗感染效果,能显著降低LPS诱导的NO释放水平和促炎因子TNF-α、IL-1β、IL-6的转录水平。综合来讲,在体外炎症模型下,R-DA的抗感染效果最为突出。
ROS作为LPS诱导炎症的重要中介,可引起DNA、蛋白质和脂质的损伤[18]。此外,过量的ROS产生和积累可诱导细胞损伤和氧化应激[19]。本研究中,LPS刺激后,ROS水平显著升高,经大黄酸以及4种衍生物孵育后,LPS诱导的RAW 264.7细胞中ROS水平显著降低,R-DA抑制ROS效果与大黄酸有显著差异,相比其他3种衍生物有更好的ROS抑制效果。有研究发现,大黄酸可以通过抑制诱导型一氧化氮合酶的表达起到抗感染作用,经修饰后的大黄酸衍生物作用后,细胞NO释放水平显著降低[20],这与Ge等[21]研究结果一致。
在大黄酸构效关系研究中,大黄酸分子上第3位碳上的取代基是羧基,增大了大黄酸的极性,是溶解性的关键基团,因此,本研究采用酯化反应的方法,将第3位碳上的羧基转化为酯键,以制备具有良好脂溶性和抗感染活性的大黄酸衍生物。据调查,对蒽醌类药物的修饰改性主要是通过在羟基和羧基引入侧链基团增加药物的吸收,引入脂肪酸可以克服母体药物的脂溶性、吸收率、生物利用度等限制。此外,多数脂肪酸具有抗菌、抗感染、抗氧化活性,例如Omega-3和Omega-6等,在人体内发挥至关重要的作用。通过引入脂肪酸可以增强大黄酸的抗感染活性,降低毒性,同时也为机体的正常运转和代谢提供必需的化学物质。前期工作中经对大黄酸衍生物的抗菌活性进行初步验证,发现其能显著增强大黄酸对金黄色葡萄球菌和肠球菌的抗菌活性,相关研究后续将继续展开,为畜禽耐药性防控提供思路,也为大黄酸进一步的研究提供基础数据。
在体外抗感染过程中,R-DA可能通过抑制NF-κB途径减少LPS刺激的巨噬细胞中促炎细胞因子的产生,也可能是由于R-DA中羟基癸醇的脂溶性作用,与细胞膜上的蛋白受体靶向结合,增加细胞膜的通透性,使药物进入细胞内发挥作用,从而起到抗感染、抗氧化的作用。
本研究中,虽然制备的大黄酸衍生物具有更低的毒性以及更优的抗感染效果,但大黄酸衍生物的纯化方法、合成产率需要进一步优化和提高。因此,在本文的基础上,仍有许多工作值得深入探究,并且要对衍生物的抗感染机制做深入性研究,如衍生物的抗感染机制是否与大黄酸的作用机制一致,以及通过结构修饰后衍生物的生物利用度是否有所增加,以期为新药的开发利用提供参考价值。
-
![]()
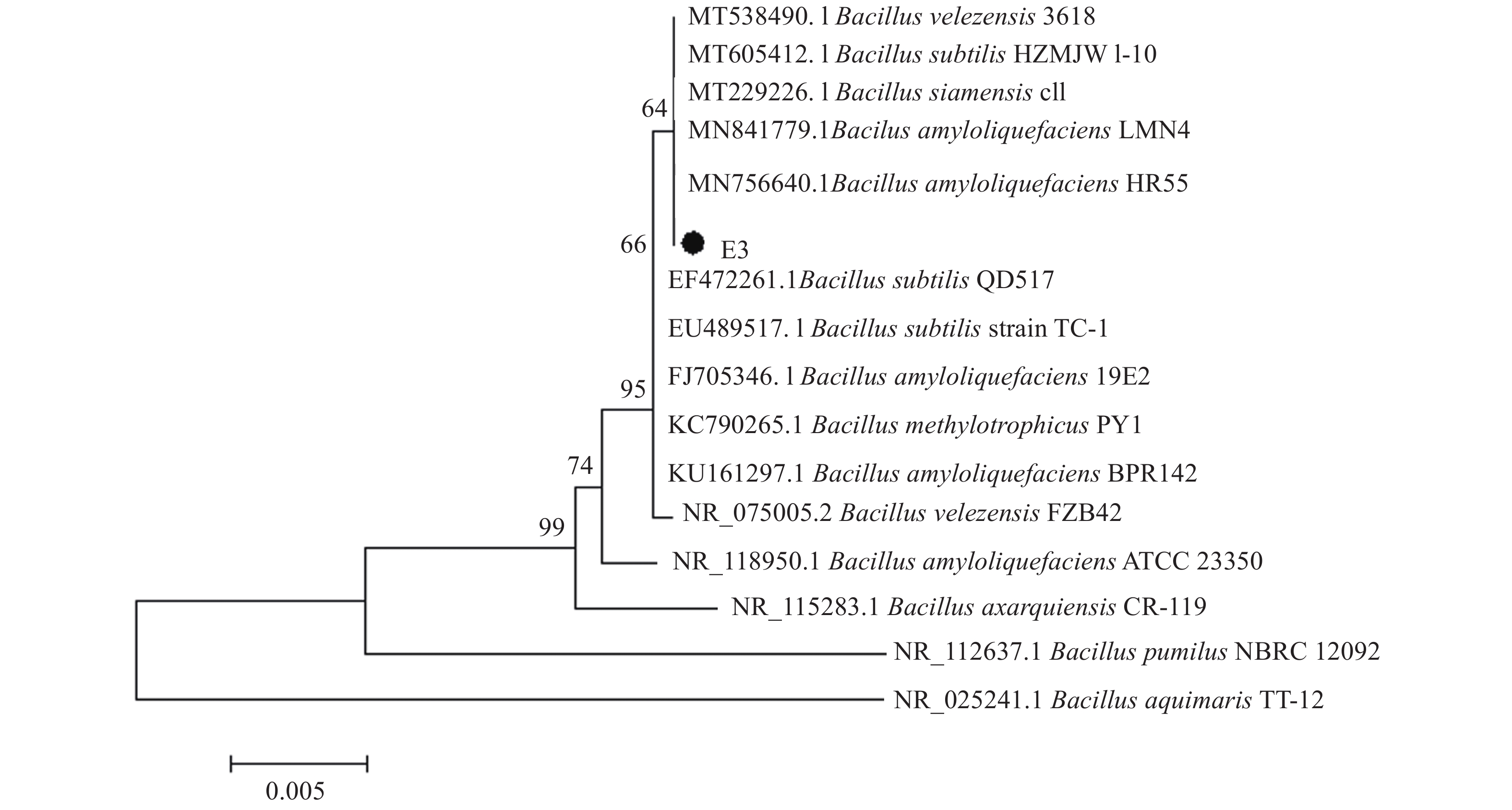
图 2 依据16S rDNA序列构建的E3菌株系统发育树
Figure 2. The phylogenetic tree of E3 strain based on 16S rDNA sequence
![]()
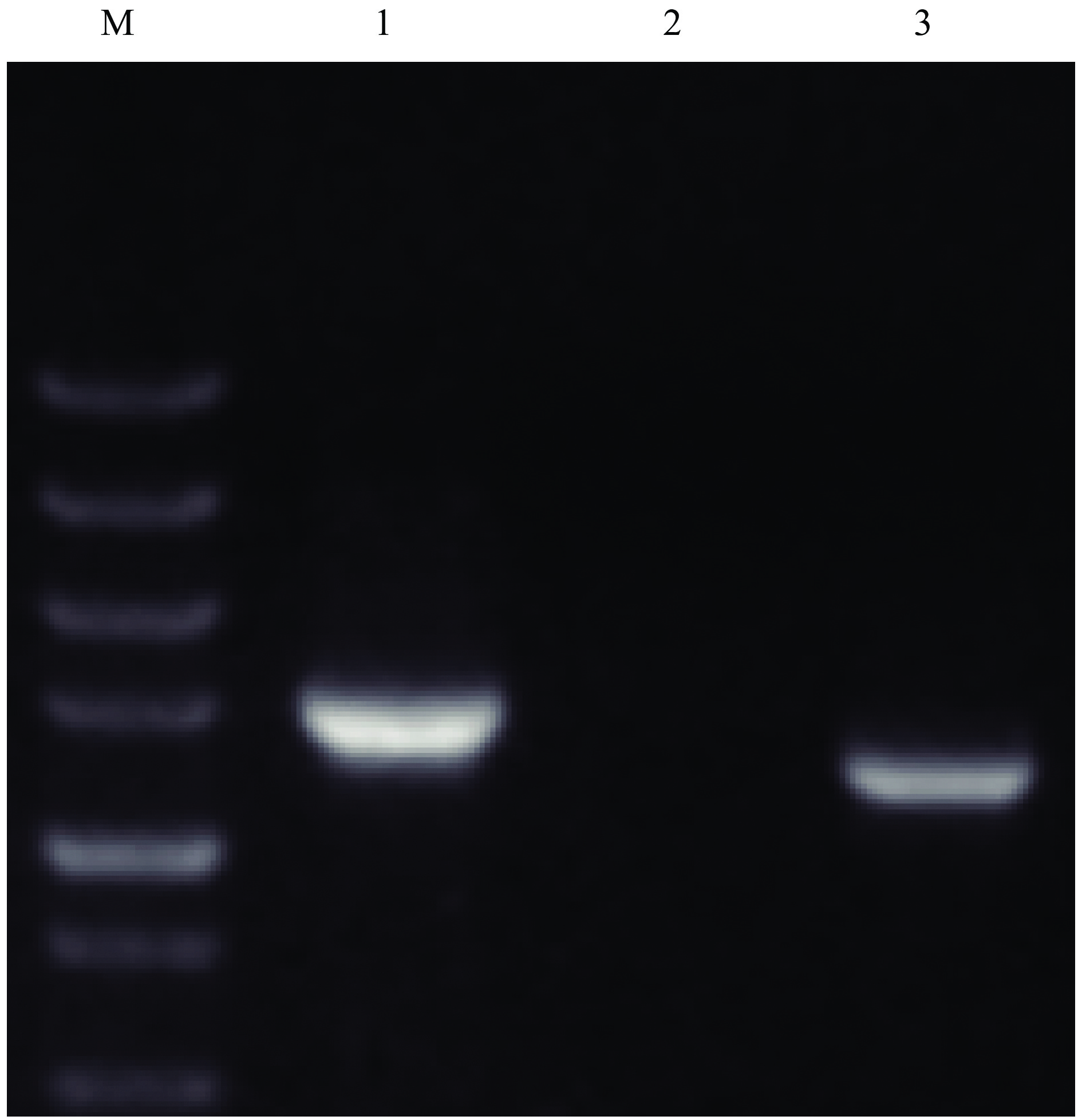
图 3 β-甘露聚糖酶基因序列特异性扩增电泳结果
M:DS 5000 marker;1:16S rDNA;2:枯草芽孢杆菌β-甘露聚糖酶基因;3:解淀粉芽孢杆菌β-甘露聚糖酶基因
Figure 3. The electrophoresis results of specific amplification of β-mannanase gene sequence
M:DS 5000 marker;1:16S rDNA;2:β-mannanase gene from Bacillus subtilis;3:β-mannanase gene from B. amyloliquefaciens
![]()
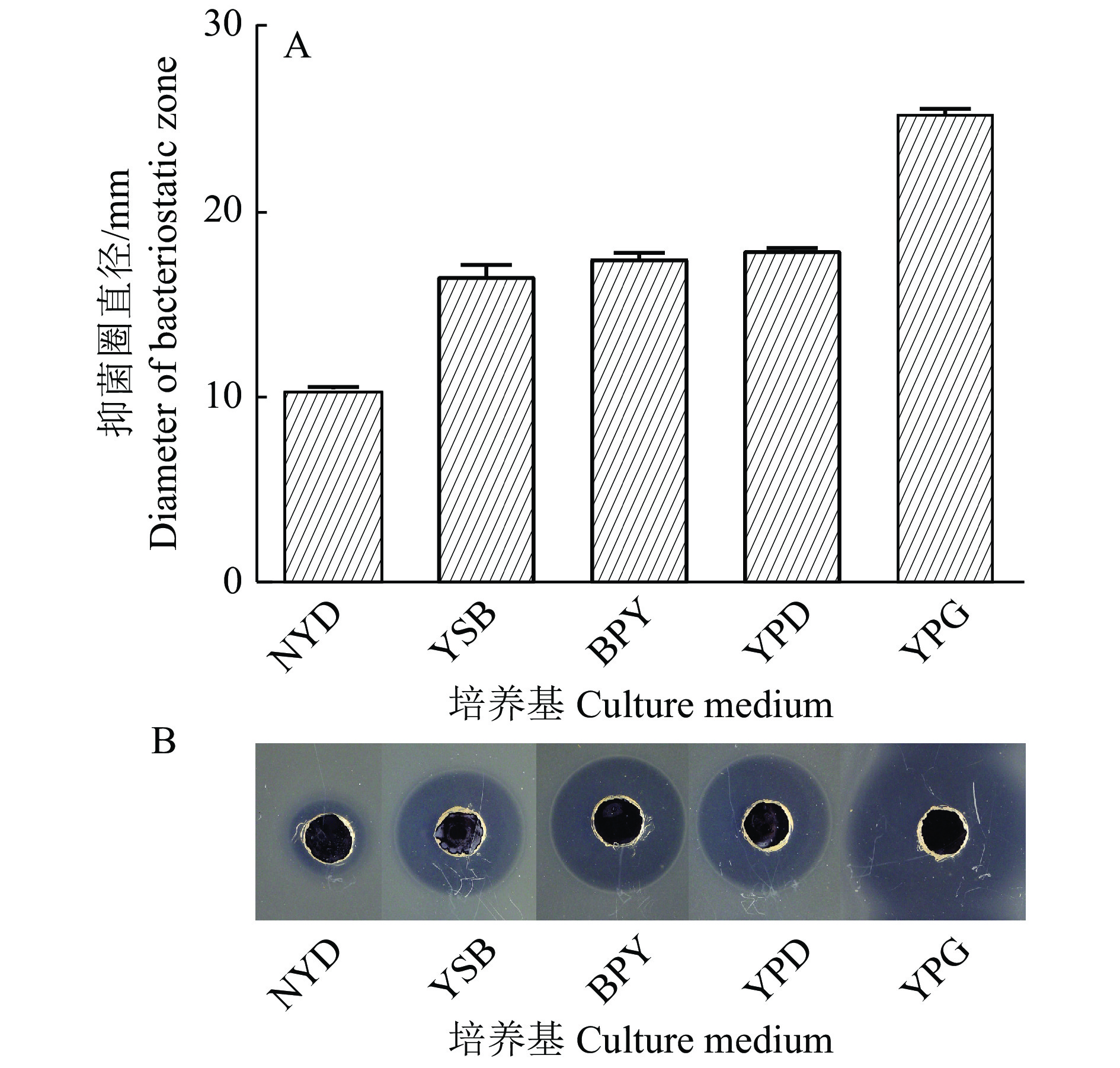
图 4 以无细胞上清液的抗菌活性为指标的培养基优化结果
Figure 4. Results of medium optimization based on the antibacterial activity of cell-free supernatant
![]()
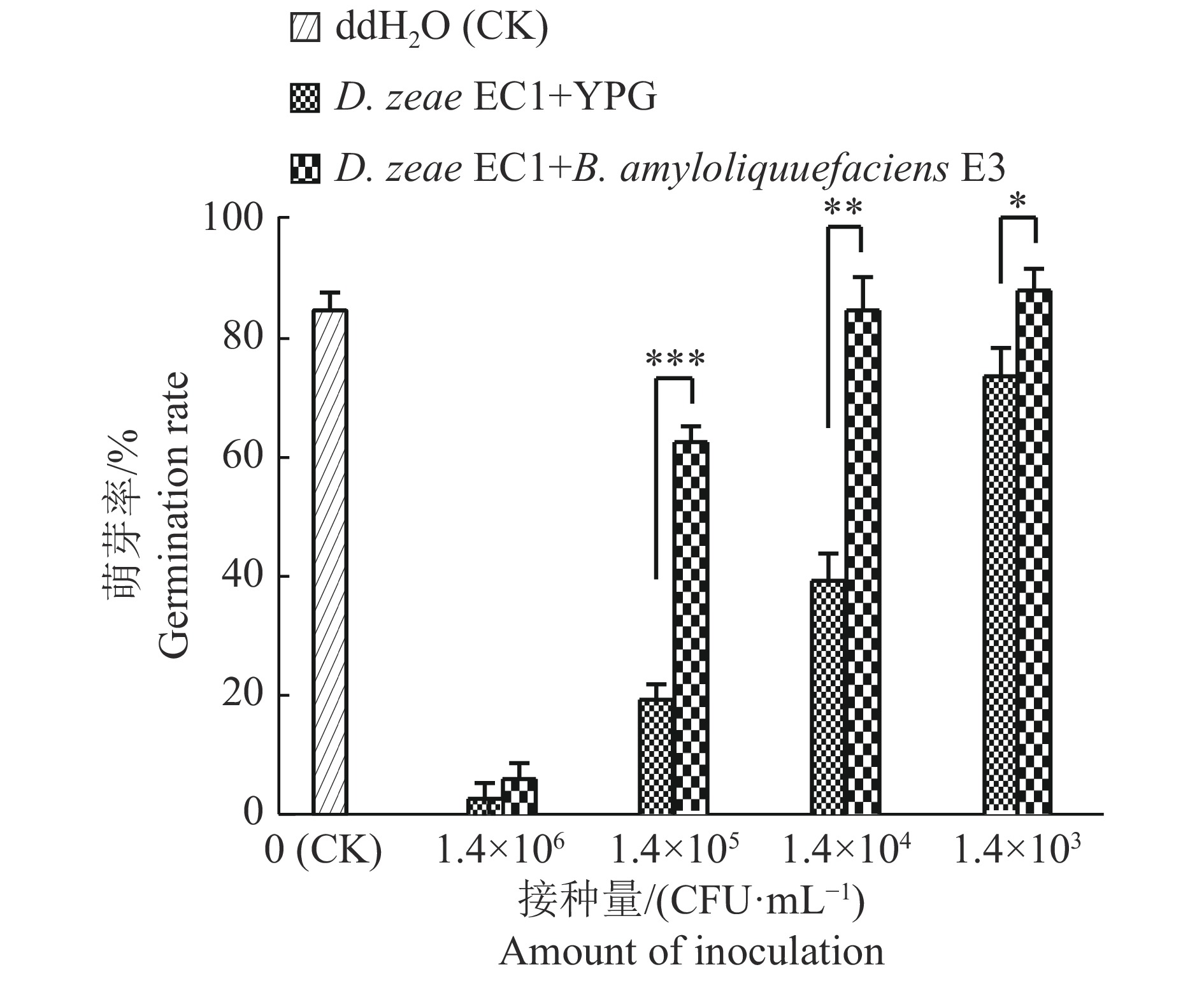
图 5 Bacillus amyloliquefacions E3菌株无菌培养液对Dickeya zeae EC1抑制水稻种子萌芽率的拮抗效果
“*”、“**”、“***”分别表示0.05、0.01和0.000 1水平的差异显著性(t检验)
Figure 5. Antagonism effect of the sterile culture medium of Bacillus amyloliquefaciens E3 strain on the inhibition of rice seeds germination by Dickeya zeae EC1
“*”,“**”,“***” indicate significance differences at the levels of 0.05,0.01 and 0.000 1 respectively (t test)
![]()
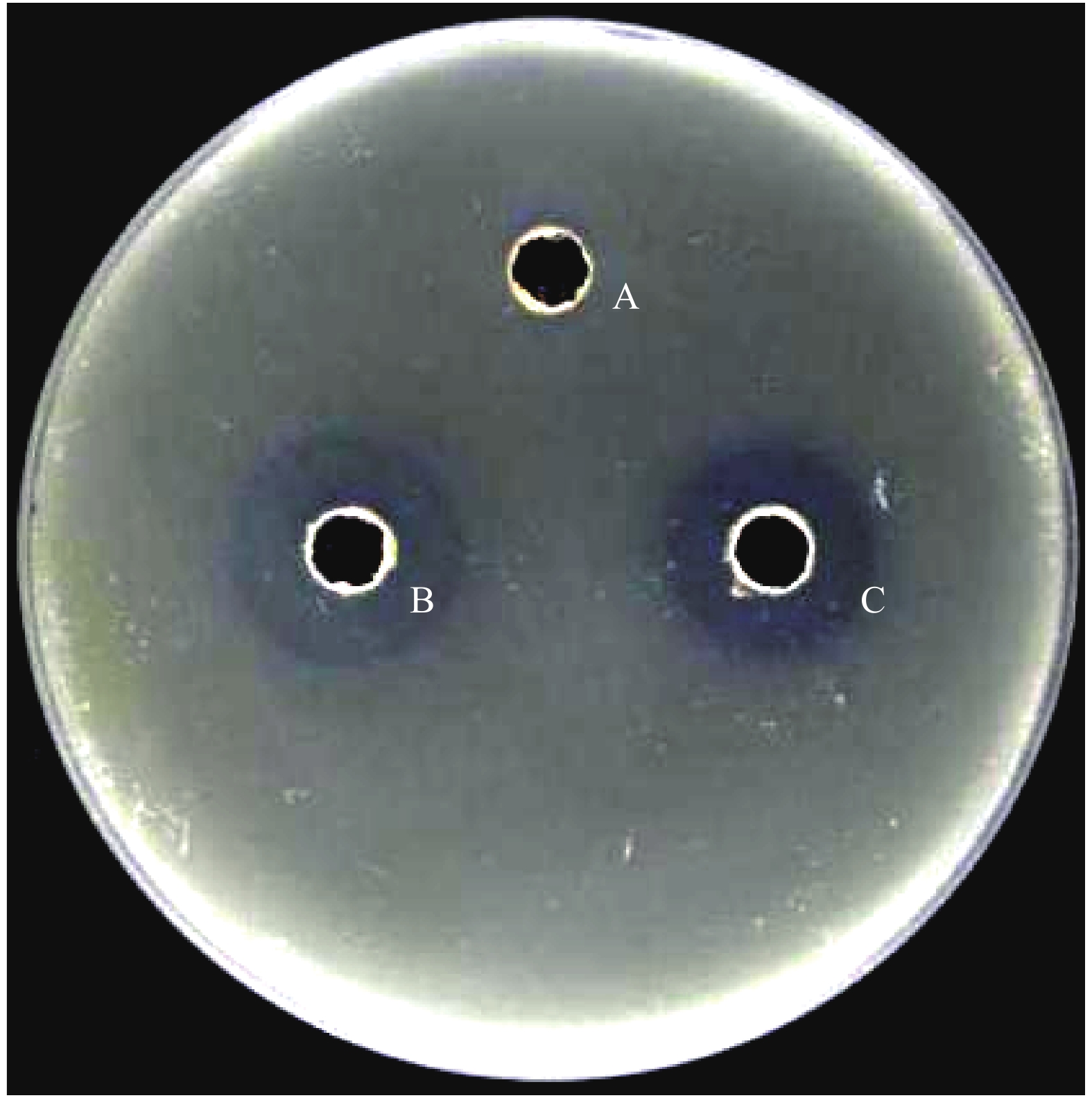
图 6 Bacillus amyloliquefaciens E3菌株粗提脂肽的抑菌活性
A:甲醇;B:无菌上清液;C:粗提脂肽类物质
Figure 6. Antimicrobial activity of crude lipopeptides extracted from Bacillus amyloliquefaciens E3 strain
A:Methanol;B:Sterile supernatant;C:The crude lipopeptides
![]()
图 7 Bacillus amyloliquefaciens E3菌株脂肽粗提物对Dickeya zeae EC1的抑制作用
“*”、“**”分别表示0.05和0.000 1水平的差异显著性(t检验)
Figure 7. Inhibitory effect of crude lipopeptides extracted from Bacillus amyloliquefaciens E3 strain against Dickeya zeae EC1
“*”,“**” indicate significance differences at the levels of 0.05 and 0.000 1 respectively (t test)
![]()
图 8 Bacillus amyloliquefaciens E3菌株的抗真菌谱
A:茄病镰刀菌Fusarium solani;B:黄曲霉Aspergillus flavus;C:黑曲霉Aspergillus niger;D:雪腐镰刀菌Fusarium nival;E:大肠埃希菌Escherichia coli;F:柑橘溃疡菌Xanthomonas citri pv. citri;G:水稻基腐病菌 Dickeya zeae EC1;H:茄科劳尔氏菌Ralstonia solanacearum;I:洋葱伯克氏菌Burkholderia cenocepacia;J:香蕉基腐病菌Dickeya zeae MS1
Figure 8. The antimicrobial spectrum of strain Bacillus amyloliquefaciens E3 strain
![]()
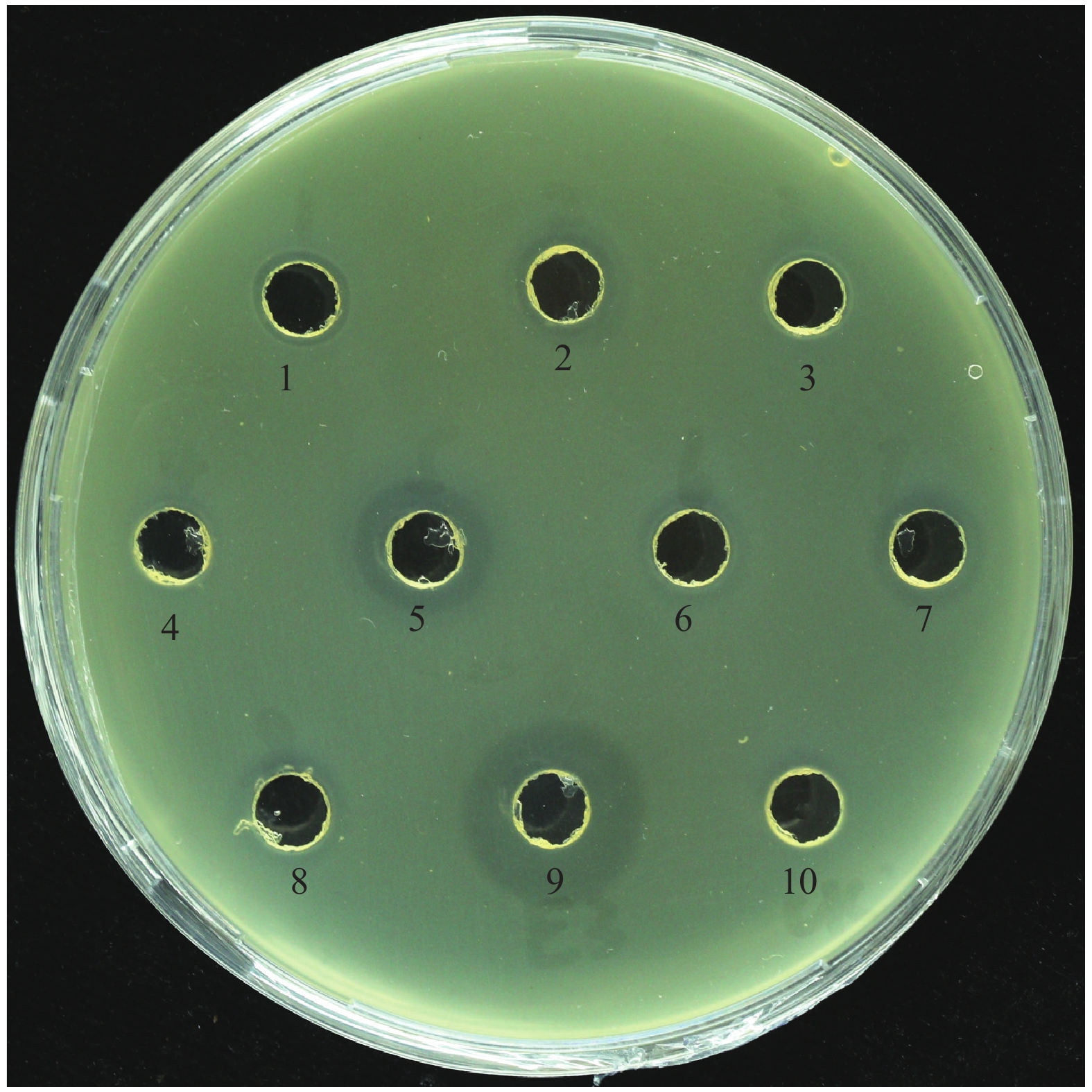
图 9 HPLC 纯化分离抑菌物质各馏分活性检测图
1~8:HPLC收集的馏分;9:脂肽粗提物;10:甲醇
Figure 9. Determination of antibacterial activity of each fraction isolated and purified by HPLC
1−8: Fractions collected by HPLC; 9: Crude lipopeptides; 10: Methanol
表 1 水稻基腐病菌拮抗菌 E3菌株的主要生理生化特性
Table 1 The main physiological and biochemical characteristics of Bacillus amyloliquefaciens E3 strain
测定指标
Detected index生化反应
Biochemical
reaction测定指标
Detected index生化反应
Biochemical
reaction接触酶 Contact enzyme 阳性 Positive 苯丙氨酸脱氢酶 Phenylalanine dehydrogenase 阴性 Negative 氧化酶 Oxidase 阴性 Negative 2%(w) NaCl耐盐试验 Salt tolerance test with 2% NaCl 阳性 Positive 明胶水解 Gelatin hydrolysis 阴性 Negative 5%(w) NaCl耐盐试验 Salt tolerance test with 5% NaCl 阳性 Positive V-P试验 V-P test 阳性 Positive 7%(w) NaCl耐盐试验 Salt tolerance test with 7% NaCl 阳性 Positive 丙酸盐利用 Propionate utilization 阴性 Negative 10%(w) NaCl耐盐试验 Salt tolerance test with 10% NaCl 阳性 Positive 脲酶试验 Urease test 阴性 Negative 淀粉水解 Starch hydrolysis 阳性 Positive 吲哚试验 Indole test 阴性 Negative 葡萄糖产酸 Glucose acidogenesis 阳性 Positive 硝酸盐还原 Nitrate reduction 阳性 Positive 柠檬酸盐利用 Citrate utilization 阳性 Positive 酪氨酸水解 Tyrosine hydrolysis 阴性 Negative 酪素水解 Casein hydrolysis 阳性 Positive pH5.7肉汤 pH5.7 Broth 阳性 Positive pH6.8肉汤 pH6.8 Broth 阳性 Positive  下载: 导出CSV
下载: 导出CSV
-
[1] 冯成玉. 水稻细菌性基腐病发生情况与研究进展[J]. 中国稻米, 2009(4): 21-23. doi: 10.3969/j.issn.1006-8082.2009.04.007 [2] 刘琼光, 张庆, 魏楚丹. 水稻细菌性基腐病研究进展[J]. 中国农业科学, 2013, 46(14): 2923-2931. doi: 10.3864/j.issn.0578-1752.2013.14.008 [3] PU X M, ZHOU J N, LIN B R, et al. First report of bacterial foot rot of rice caused by a Dickeya zeae in China[J]. Plant Disease, 2012, 96(12): 1818.
[4] 张蝶. 浙江省水稻细菌性基腐病发生因素及化学防治研究[D]. 金华: 浙江师范大学, 2016. [5] SANSINENEA E, VACA J, ELENA ROJAS N, et al. A wide spectrum of antibacterial activity of secondary metabolites from Bacillus amyloliquefaciens ELI149[J]. Bioscience Journal, 2020, 36(1): 235-244.
[6] BEN ABDALLAH D, TOUNSI S, GHARSALLAH H, et al. Lipopeptides from Bacillus amyloliquefaciens strain 32a as promising biocontrol compounds against the plant pathogen Agrobacterium tumefaciens[J]. Environmental Science and Pollution Research, 2018, 25(36): 36518-36529.
[7] STEIN T. Bacillus subtilis antibiotics: Structures, syntheses and specific functions[J]. Molecular Microbiology, 2005, 56(4): 845-857. doi: 10.1111/j.1365-2958.2005.04587.x
[8] WU L M, WU H J, CHEN L N, et al. Difficidin and bacilysin from Bacillus amyloliquefaciens FZB42 have antibacterial activity against Xanthomonas oryzae rice pathogens[J]. Scientific Reports, 2015, 5(1). doi: http://doi.org/10.1038/srep12975.
[9] LIU Y F, CHEN Z Y, NG T B, et al. Bacisubin, an antifungal protein with ribonuclease and hemagglutinating activities from Bacillus subtilis strain B-916[J]. Peptides, 2007, 28(3): 553-559. doi: 10.1016/j.peptides.2006.10.009
[10] TAO Y, BIE X M, LV F X, et al. Antifungal activity and mechanism of fengycin in the presence and absence of commercial surfactin against Rhizopus stolonifer[J]. Journal of Microbiology, 2011, 49(1): 146-150. doi: 10.1007/s12275-011-0171-9
[11] CHO S J, LEE S K, CHA B J, et al. Detection and characterization of the Gloeosporium gloeosporioides growth inhibitory compound iturin A from Bacillus subtilis strain KS03[J]. FEMS Microbiology Letters, 2003, 223(1): 47-51. doi: 10.1016/S0378-1097(03)00329-X
[12] CAO H Y, JIAO Y, YIN N, et al. Analysis of the activity and biological control efficacy of the Bacillus subtilis strain Bs-1 against Meloidogyne incognita[J]. Crop Protection, 2019, 122: 125-135. doi: 10.1016/j.cropro.2019.04.021
[13] ZHAO P, QUAN C, WANG Y, et al. Bacillus amyloliquefaciens Q-426 as a potential biocontrol agent against Fusarium oxysporum f. sp spinaciae[J]. Journal of Basic Microbiology, 2014, 54(5): 448-456. doi: 10.1002/jobm.201200414
[14] ARGUELLES-ARIAS A, ONGENA M, HALIMI B, et al. Bacillus amyloliquefaciens GA1 as a source of potent antibiotics and other secondary metabolites for biocontrol of plant pathogens[J]. Microbial Cell Factories, 2009, 8: 63.
[15] ONGENA M, JACQUES P. Bacillus lipopeptides: Versatile weapons for plant disease biocontrol[J]. Trends in Microbiology, 2008, 16(3): 115-125. doi: 10.1016/j.tim.2007.12.009
[16] CAWOY H, MARIUTTO M, HENRY G, et al. Plant defense stimulation by natural isolates of Bacillus depends on efficient surfactin production[J]. Molecular Plant-Microbe Interactions, 2014, 27(2): 87-100. doi: 10.1094/MPMI-09-13-0262-R
[17] ZHOU J, CHENG Y, LV M, et al. The complete genome sequence of Dickeya zeae EC1 reveals substantial divergence from other Dickeya strains and species[J]. BMC Genomics, 2015, 16(1): 571. doi: 10.1186/s12864-015-1545-x
[18] 东秀珠, 蔡妙英. 常见细菌系统鉴定手册[M]. 北京: 科学出版社, 2001: 267-294. [19] 布坎南, 等. 伯杰细菌鉴定手册[M]. 北京: 科学出版社, 1984: 729-758. [20] REDBURN A C, PATEL B K. Phylogenetic analysis of Desulfotomaculum thermobenzoicum using polymerase chain reaction-amplified 16S rRNA-specific DNA[J]. FEMS Microbiology Letters, 1993, 113(1): 81-86. doi: 10.1111/j.1574-6968.1993.tb06492.x
[21] 刘勇, 毛爱军, 李辉, 等. 枯草芽孢杆菌群β-甘露聚糖酶基因克隆及同源性分析[J]. 基因组学与应用生物学, 2009, 28(5): 845-850. doi: 10.3969/gab.028.000845 [22] 韩玉竹, 邓钊, 张宝, 等. 解淀粉芽孢杆菌H15产抗菌肽的发酵条件优化和提取方法比较研究[J]. 食品科学, 2015, 36(15): 135-141. doi: 10.7506/spkx1002-6630-201515025 [23] 张睿佳, 胡杰, 陆建忠, 等. 不同成熟度对水稻种子萌发的影响[J]. 上海农业科技, 2017(5): 62-63. doi: 10.3969/j.issn.1001-0106.2017.05.029 [24] LI S B, XU S R, ZHANG R N, et al. The antibiosis action and rice-induced resistance, mediated by a lipopeptide from Bacillus amyloliquefaciens B014, in controlling rice disease caused by Xanthomonas oryzae pv. oryzae[J]. Journal of Microbiology and Biotechnology, 2016, 26(4): 748-756. doi: 10.4014/jmb.1510.10072
[25] 朱弘元, 康健, 范昕, 等. 解淀粉芽孢杆菌B15产脂肽的分离鉴定及抑菌机理[J]. 江苏农业科学, 2016, 44(5): 186-189. [26] 张玉胜, 谭亚男. 油菜核盘菌拮抗菌的筛选及鉴定[J]. 生命科学仪器, 2009, 7(8): 27-28. [27] 陈华, 王丽, 袁成凌, 等. 高效液相色谱-电喷雾质谱法分离和鉴别枯草芽孢杆菌产生的脂肽类化合物[J]. 色谱, 2008(3): 343-347. doi: 10.3321/j.issn:1000-8713.2008.03.016 [28] CHEN M C, WANG J P, ZHU Y J, et al. Antibacterial activity against Ralstonia solanacearum of the lipopeptides secreted from the Bacillus amyloliquefaciens strain FJAT-2349[J]. Journal of General and Applied Microbiology, 2019, 126(5): 1519-1529. doi: 10.1111/jam.14213
[29] 周海亮. 水稻细菌性基腐病病原学及检测技术的研究[D]. 武汉: 华中农业大学, 2012. [30] VATER J, GAO X, HITZEROTH G, et al. "Whole cell"--matrix-assisted laser desorption ionization-time of flight-mass spectrometry, an emerging technique for efficient screening of biocombinatorial libraries of natural compounds-present state of research[J]. Combinatorial Chemistry& High Throughput Screening, 2003, 6(6): 557-567.
[31] AERON A, KHARE E, JHA C K, et al. Revisiting the plant growth-promoting rhizobacteria: Lessons from the past and objectives for the future[J]. Archives of Microbiology, 2020, 202(4): 665-676. doi: 10.1007/s00203-019-01779-w
[32] GOUDA S, KERRY R G, DAS G, et al. Revitalization of plant growth promoting rhizobacteria for sustainable development in agriculture[J]. Research in Microbiology, 2018, 206: 131-140. doi: 10.1016/j.micres.2017.08.016
[33] KHAYALETHU N, LESIBA K L, MOLEMI E R, et al. The mode of action of bacillus species against Fusarium graminearum, tools for investigation, and future prospects[J]. Toxins, 2019, 11(10). doi: http://doi.org/10.3390/toxins11100606.
[34] 严婉荣, 肖敏, 陈圆, 等. 解淀粉芽孢杆菌抗菌脂肽研究进展[J]. 北方园艺, 2018(7): 162-167. [35] FAN B, WANG C, SONG X, et al. Bacillus velezensis FZB42 in 2018: The gram-positive model strain for plant growth promotion and biocontrol[J]. Frontiers in Microbiology, 2018, 9: 2491. doi: 10.3389/fmicb.2018.02491
[36] CHOWDHURY S P, UHL J, GROSCH R, et al. Cyclic lipopeptides of Bacillus amyloliquefaciens subsp. plantarum colonizing the lettuce rhizosphere enhance plant defense responses toward the bottom rot pathogen Rhizoctonia solani[J]. Molecular Plant-Microbe Interactions, 2015, 28(9): 984-995. doi: 10.1094/MPMI-03-15-0066-R
[37] 向亚萍, 周华飞, 刘永锋, 等. 解淀粉芽孢杆菌B1619脂肽类抗生素的分离鉴定及其对番茄枯萎病菌的抑制作用[J]. 中国农业科学, 2016, 49(15): 2935-2944. doi: 10.3864/j.issn.0578-1752.2016.15.008 [38] FIRA D, DIMKIC I, BERIC T, et al. Biological control of plant pathogens byBacillus species[J]. Biotechnology Journal, 2018, 285: 44-55. doi: 10.1016/j.jbiotec.2018.07.044
[39] ZERIOUH H, ROMERO D, GARCIA-GUTIERREZ L, et al. The iturin-like lipopeptides are essential components in the biological control arsenal of Bacillus subtilis against bacterial diseases of cucurbits[J]. Molecular Plant-Microbe Interactions, 2011, 24(12): 1540-1552. doi: 10.1094/MPMI-06-11-0162
[40] CAO Y, PI H L, CHANDRANGSU P, et al. Antagonism of two plant-growth promoting Bacillus velezensis isolates against Ralstonia solanacearum and Fusarium oxysporum[J]. Scientific Reports, 2018, 8(1). doi: http://doi.org/10.1038/S41598-018-22782-Z.
[41] HADIZADEH I, PEIVASTEGAN B, HANNUKKALA A, et al. Biological control of potato soft rot caused by Dickeya solani and the survival of bacterial antagonists under cold storage conditions[J]. Plant Pathology, 2019, 68(2): 297-311. doi: 10.1111/ppa.12956
[42] CHEN J, LIU T, WEI M, et al. Macrolactin a is the key antibacterial substance of Bacillus amyloliquefaciens D2WM against the pathogen Dickeya chrysanthemi[J]. European Journal of Plant Pathology, 2019, 155(2): 393-404. doi: 10.1007/s10658-019-01774-3
[43] LI J, HU M, XUE Y, et al. Screening, identification and efficacy evaluation of antagonistic bacteria for biocontrol of soft rot disease caused by Dickeya zeae[J]. Microorganisms, 2020, 8(6975). doi: 10.3390/microorganisms8050697.












计量
- 文章访问数: 566
- HTML全文浏览量: 41
- PDF下载量: 859
