
Traversal path planning of agricultural robot based on memory simulated annealing and A* algorithm
-
摘要:目的
解决农业机器人大田作业时遍历路径规划的问题。
方法提出一种记忆模拟退火与A*算法相结合的遍历算法。首先通过记忆模拟退火算法搜索出任务最优目标点行走顺序,然后使用A*算法进行跨区域衔接路径规划。
结果仿真试验结果表明,该算法规划的遍历路径曼哈顿距离比传统模拟退火算法减少了9.4%,遍历路径覆盖率能达到100%,重复率控制为4.2%。
结论记忆模拟退火通过为传统模拟退火算法增加记忆器,增强了跳出局部最优陷阱的能力,提高了算法所得解的质量。该研究结果可为农业机器人遍历路径规划提供理论基础。
Abstract:ObjectiveTo solve the problem of traversal path planning of agricultural robot in field operation.
MethodA memory simulated annealing algorithm combined with A* algorithm was proposed. Firstly, the optimal walking sequence of target points in task was found by memory simulated annealing algorithm, and then A* algorithm was used for crossing regional linking of path planning.
ResultThe simulation experiments showed that the Manhattan distance of traversal path planned by this algorithm was reduced by 9.4% compared with the traditional simulated annealing algorithm, the coverage of traversal path could reach 100%, and the repetition rate could be controlled at 4.2%.
ConclusionMemory simulated annealing algorithm enhances the ability to jump out of the local optimal trap, and improves the quality of the solution obtained by adding memory device to the traditional simulated annealing algorithm. The research results can provide a theoretical basis for the path planning of agricultural robot.
-
自噬是真核生物体内一种重要的分解代谢过程。错误折叠的蛋白或受损的细胞器被自噬体包裹运输至液泡,在液泡腔中被水解酶降解[1]。分解产物会被储存于液泡,或被运输回细胞质中重复利用[2]。在模式植物拟南芥Arabidopsis thaliana中,现已鉴定到大约40个自噬相关基因ATGs(Autophagy-related genes),且大部分ATGs对自噬体的形成是必需的[3-4]。
研究发现,过表达ATGs可以增强植物的环境适应性。自噬作为物质循环途径之一,对氮素的再利用和营养饥饿的抵抗至关重要。过表达AtATG8a、AtATG8e、AtATG8f和AtATG8g的拟南芥植株氮素再活化能力增强,转基因植株中有更多的氮素从衰老叶片中活化并运输到种子中,从而使种子的蛋白质含量显著提高[5]。拟南芥中异源过表达水稻OsATG8b或苹果MdATG18a均可以增强拟南芥对氮饥饿或碳饥饿的耐受性[6-8]。自噬过程也是植物抗旱、抗盐的关键途径之一。在干旱条件下,过表达MdATG18a和MdATG8i的苹果植株,以及异源过表达香蕉MaATG8f的拟南芥植株,都表现出更强的耐干旱能力,而且在这些过表达植株中,光合效率提高,可溶性糖和氨基酸的积累量明显增加[9-11]。在高盐环境下,过表达MdATG10苹果植株的根系具有更强的水分吸收能力,植株能够保持较高的光合效率从而维持生长[12]。除此之外,过表达MdATG18a的苹果植株不仅对碱性和高温等不良环境的耐受力增强,而且对斑褐病的免疫力也明显提高[13-15]。
自噬相关蛋白ATG7通过催化ATG8-ATG3偶联物的形成激活ATG8的脂化作用,被认为是自噬过程中的关键限速因子之一[16]。atg7缺失突变体不能形成自噬体,在碳/氮饥饿条件下生长受阻,叶片早衰,繁殖能力下降[2, 17]。过表达AtATG7的拟南芥植株自噬活性显著提高,转基因植株不仅具有衰老延缓、生物量和产量提高、种子含油量增加等优势,而且对病原菌的抗性也明显增强[16]。
苜蓿Medicago spp.是一种广泛种植的饲料作物,同时也是豆科模式植物之一[18]。通过过表达苜蓿ATGs使自噬活性水平提高,是改善苜蓿农艺性状的一种潜在手段。然而苜蓿ATGs的生物学功能是否保守,以及其对植物的抗逆能力是否有促进作用在很大程度上仍然未知。本文以蒺藜苜蓿Medicago truncatula中关键的自噬相关基因MtATG7为研究对象,构建拟南芥异源过表达MtATG7的稳定遗传植株,并在碳/氮饥饿条件下进行表型分析。本研究旨在为利用MtATG7基因改良苜蓿和其他植物的农艺性状提供理论依据。
1. 材料与方法
1.1 植物材料与培养条件
拟南芥生态型为Columbia,拟南芥T-DNA插入的自噬功能缺陷突变体atg7-3(SAIL_11_H07)和GFP-ATG8e植株为Chen等保存[19],通过杂交得到atg7-3×GFP-ATG8e纯和植株,构建35S::MtATG7-NOS过表达载体,通过农杆菌Agrobacterium tumefaciens介导侵染Col、atg7-3、atg7-3×GFP-ATG8e的过表达株系获得MtATG7-ox、atg7/MtATG7、MtATG7/atg7-3/GFP-ATG8e转基因植株。
蒺藜苜蓿生态型为A17。
培养条件:光暗周期为16 h光照/8 h黑暗,温度为22 ℃,光照强度为120 µmol·m−2·s−1。
1.2 植物表达载体构建及拟南芥遗传转化
以蒺藜苜蓿cDNA为模板,利用常规PCR方法扩增MtATG7基因的编码序列(2097 bp)。分别在上、下游引物中引入BamHI和PstI的酶切位点,通过BamHI/PstI双酶切插入到双元表达载体pCambia1300,使用无缝克隆试剂盒(金沙生物,SC612)进行连接,完成携带35S::MtATG7的植物表达载体构建。使用农杆菌GV3101介导的花序浸泡法[20]转化拟南芥Col-0、atg7-3和atg7-3×GFP-ATG8e植株。将转基因拟南芥T0代种子在含有25 mg/L潮霉素的筛选培养基上经过3代筛选获得6个纯合株系,选取2个代表株系用于后续试验。所有构建均经测序验证正确,构建引物信息见表1。
表 1 PCR特异性引物序列Table 1. Specific primers used for PCR引物类型
Primer type引物名称
Primer name引物序列(5′→3′)
Primer sequence目的基因引物
Target gene primerMtATG7-F gaacacgggggactGGATCCATGGCTTTGCTCCAATTTAT MtATG7-R AATGTTTGAACGATCTGCAGTCATATTTCAAAACAATCTT 自噬突变体鉴定引物
Autophagy mutant identification primeratg7-F TCTCTTGTTGGTCAAGCCTC atg7-R CATTGTTGGTCTAGAGTTCG LB1 GCCTTTTCAGAAATGGATAAATAGCCTTGCTT 1.3 MtATG7基因的生物信息学分析
通过Phytozomev13网站(https://phytozome-next.jgi.doe.gov/)查询不同物种中ATG7基因的氨基酸序列,利用Pfam数据库(http://pfam.xfam.org/search/sequence)分析不同物种ATG7基因的保守基序;利用MEGA11软件构建系统发育树。
1.4 缺碳胁迫处理和存活率测定
缺碳处理:将在1/2MS培养基平板上生长11 d的拟南芥幼苗,转移至无糖1/2MS培养基平板上,并放置于黑暗环境下培养使植物不能进行光合作用,继续培养9 d后进行表型观察和拍照。缺碳存活率的测定:将土中生长3周的拟南芥植株转移至黑暗条件下培养7 d,随后转移到正常生长条件下恢复8 d,统计植株存活率。试验经3次生物学重复。
1.5 缺氮胁迫处理
拟南芥幼苗在正常1/2MS培养基平板上生长11 d后,转移至缺氮1/2MS培养基平板上,继续培养7 d后进行表型观察和拍照。
1.6 叶绿素含量的测定
取适量拟南芥幼苗称量鲜质量,加入2 mL N,N−二甲基甲酰胺(N,N-Dimethylformamide, DMF),4 ℃遮光提取2 d,每个样品设3个生物学重复。按以下公式计算叶绿素含量:叶绿素浓度= 20.21×D645 nm+8.02×D663 nm,叶绿素含量=叶绿素浓度×提取液体积×稀释倍数/样品鲜质量。
1.7 蛋白提取及Western blot检测自噬活性
取适量2周龄的拟南芥幼苗,整株取样至2 mL离心管中,经液氮速冻后,用均质破碎仪研磨样品。加入200 μL RIPA Buffer并充分振荡混匀,4 ℃放置10 min;4 ℃、12000 r·min−1离心 30 min,吸取上清。蛋白溶液经95 ℃加热5 min后,使用变性聚丙烯酰胺凝胶(SDS-PAGE)进行电泳分离。使用PVDF膜,在4 ℃条件下恒电流200 mA转膜120 min。将膜置于质量分数为5%的脱脂奶粉中封闭1 h,然后使用Anti-GFP(Invitrogen A11122)抗体4 ℃孵育过夜。1×TBST洗膜3次后孵育二抗2 h。1×TBST溶液重复洗膜3次,将PVDF膜放至干净的培养皿中,用显色液试剂盒进行显色后,使用化学发光成像仪拍照保存。
1.8 数据统计分析
试验所得数据使用统计软件SPSS 19.0进行分析,其中叶绿素含量测定数据n=8,存活率数据n=3,样品之间差异采用单因素方差分析和LSD多重比较方法进行统计检验,P<0.05视为差异显著,数据用平均值±标准差表示。
2. 结果与分析
2.1 ATG7蛋白结构在植物中高度保守
为了解蒺藜苜蓿MtATG7的进化特征和生物学功能,本研究对MtATG7的氨基酸序列与拟南芥等其他植物的同源基因构建了系统发育树,并分析了MtATG7及其同源基因的序列相似性。结果表明,在分析的15种植物中,蒺藜苜蓿的ATG7与红车轴草Trifolium pratense、鹰嘴豆Cicer arietinum的ATG7亲缘关系最近,与模式植物拟南芥的亲缘关系相对较远。利用Pfam数据库中对ATG7的氨基酸序列进行结构域分析,发现在所有15种植物中都具有典型的ATG7特征结构域,说明ATG7的功能很可能是高度保守的(图1)。
![]() 图 1 不同植物ATG7基因的系统发育树分析和保守结构域预测Figure 1. Phylogenetic tree analysis and domain prediction of ATG7 gene from different species
图 1 不同植物ATG7基因的系统发育树分析和保守结构域预测Figure 1. Phylogenetic tree analysis and domain prediction of ATG7 gene from different species2.2 MtATG7基因的克隆及异源过表达植株的遗传转化
根据MtATG7的DNA序列设计PCR引物,以蒺藜苜蓿的cDNA为模板进行PCR扩增(表1)。PCR产物经琼脂糖凝胶电泳后得到约2000 bp的特异条带,与2097 bp的MtATG7编码序列长度吻合,说明MtATG7基因扩增成功(图2A)。进一步使用无缝克隆方法,将MtATG7序列连接到表达载体上,构建了35S::MtATG7双元过表达载体(图2B)。在农杆菌介导下,通过花序浸泡法分别侵染拟南芥野生型(Col)、atg7-3和atg7-3/GFP-ATG8e植株,获得了多个独立的过表达株系,包括MtATG7-OX、atg7/MtATG7和atg7-3/MtATG7/GFP-ATG8e植株。每个基因型植株挑选两个纯合的株系进行后续试验。
![]() 图 2 MtATG7过表达载体的构建A:苜蓿MtATG7基因的克隆;B:35S::MtATG7过表达载体的菌落PCR结果,M:Trans2K Plus II DNA Marker,1~10:菌落;目标条带大小为2097 bp左右Figure 2. Construction of MtATG7 overexpression vectorA: Cloning of MtATG7 gene in Medicago; B: Colony PCR results of 35S::MtATG7 overexpression vector, M: Trans2KPlus II DNA Marker, 1−10: Colony; The target band size is about 2097 bp
图 2 MtATG7过表达载体的构建A:苜蓿MtATG7基因的克隆;B:35S::MtATG7过表达载体的菌落PCR结果,M:Trans2K Plus II DNA Marker,1~10:菌落;目标条带大小为2097 bp左右Figure 2. Construction of MtATG7 overexpression vectorA: Cloning of MtATG7 gene in Medicago; B: Colony PCR results of 35S::MtATG7 overexpression vector, M: Trans2KPlus II DNA Marker, 1−10: Colony; The target band size is about 2097 bp2.3 过表达MtATG7减缓拟南芥叶片在碳饥饿胁迫下的衰老速率
将苗龄11 d的转基因植株转移至无糖培养基上,并放置在黑暗环境下培养,使植物光合作用停止并产生碳饥饿。在缺碳胁迫条件下,atg7幼苗的叶片明显变黄并伴随白化现象,但野生型Col与互补植株atg7/MtATG7#1,atg7/MtATG7#2的叶片仍呈现绿色,说明MtATG7能够挽救atg7突变体的生长缺陷表型(图3A、3B)。过表达植株MtATG7-OX#1和MtATG7-OX#2和野生型的叶片一样都保持相对嫩绿。对缺碳胁迫前后的幼苗进行叶绿素含量的测定,结果显示缺碳胁迫后,atg7突变体的叶绿素含量和野生型相比显著下降,但异源转基因的atg7/MtATG7植株和atg7突变体相比叶绿素含量明显提高(图4)。该结果表明MtATG7能够互补AtATG7抵抗碳饥饿胁迫的生物学功能。
![]() 图 3 MtATG7促进转基因拟南芥抵抗碳胁迫A:将苗龄11 d的野生型Col、自噬突变体atg7以及转基因atg7/MtATG7#1、atg7/MtATG#2、MtATG7-OX#1、MtATG7-OX#2拟南芥幼苗移到不含糖的1/2MS培养基中,黑暗条件下培养9 d后的表型;B:在碳胁迫9 d后分别对每个株系取2株幼苗拍照;比例尺=5 mmFigure 3. MtATG7 promotes resistance to carbon starvation in transgenic ArabidopsisA: Phenotypes for 11-day-old wild-type Col, autophagy mutant atg7, and transgenic atg7/MtATG7#1, atg7/MtATG#2, MtATG7-OX#1, MtATG7-OX#2 Arabidopsis seedlings after transferred to sugar-free 1/2MS medium and cultured under dark conditions for 9 days; B: After 9 days of carbon starvation stress, two seedlings from each line were photographed; Scale bar = 5 mm
图 3 MtATG7促进转基因拟南芥抵抗碳胁迫A:将苗龄11 d的野生型Col、自噬突变体atg7以及转基因atg7/MtATG7#1、atg7/MtATG#2、MtATG7-OX#1、MtATG7-OX#2拟南芥幼苗移到不含糖的1/2MS培养基中,黑暗条件下培养9 d后的表型;B:在碳胁迫9 d后分别对每个株系取2株幼苗拍照;比例尺=5 mmFigure 3. MtATG7 promotes resistance to carbon starvation in transgenic ArabidopsisA: Phenotypes for 11-day-old wild-type Col, autophagy mutant atg7, and transgenic atg7/MtATG7#1, atg7/MtATG#2, MtATG7-OX#1, MtATG7-OX#2 Arabidopsis seedlings after transferred to sugar-free 1/2MS medium and cultured under dark conditions for 9 days; B: After 9 days of carbon starvation stress, two seedlings from each line were photographed; Scale bar = 5 mm![]() 图 4 碳胁迫前后的拟南芥幼苗叶绿素含量数据为平均值±标准差, n=8;相同处理柱子上方的不同小写字母表示各株系之间差异显著(P<0.05,LSD法)Figure 4. Chlorophyll content of Arabidopsis seedling before and after carbon stressData are means ± SDs, n=8; Different lowercase letters on bars of the same treatment indicate significant differences among strains (P<0.05,LSD test)
图 4 碳胁迫前后的拟南芥幼苗叶绿素含量数据为平均值±标准差, n=8;相同处理柱子上方的不同小写字母表示各株系之间差异显著(P<0.05,LSD法)Figure 4. Chlorophyll content of Arabidopsis seedling before and after carbon stressData are means ± SDs, n=8; Different lowercase letters on bars of the same treatment indicate significant differences among strains (P<0.05,LSD test)2.4 过表达MtATG7基因提高拟南芥在碳饥饿胁迫下的存活率
将苗龄3周的拟南芥转移到黑暗条件下处理7 d,探究过表达MtATG7对拟南芥碳饥饿胁迫存活率的影响。研究发现,在碳饥饿胁迫下,atg7因自噬功能缺陷导致生长受到抑制,即使恢复光照也无法存活,但自噬功能正常的植株都有一定比例植株能够存活下来(图5)。值得注意的是,多个过表达MtATG7的植株,包括atg7/MtATG7#1、MtATG7-OX#1和MtATG7-OX#2,存活率和野生型Col相比都明显提高(图6)。上述结果表明过表达MtATG7基因可以提高拟南芥对碳饥饿胁迫的耐受能力。
![]() 图 5 MtATG7促进转基因拟南芥在碳胁迫下的生存图中为将苗龄3周的野生型Col、自噬突变体atg7以及转基因atg7/MtATG7#1、atg7/MtATG#2、MtATG7-OX#1、MtATG7-OX#2拟南芥植株黑暗处理7 d,恢复光照8 d后的表型;比例尺=1 cmFigure 5. MtATG7 promotes survival of transgenic Arabidopsis under carbon starvation3-week-old Arabidopsis seedlings of wild type Col, autophagy mutant atg7, and transgenic lines atg7/MtATG7#1, atg7/MtATG#2, MtATG7-OX#1, and MtATG7-OX#2 were treated in the dark for 7 days,and the phenotypes after 8 days of recovery under light are shown in the figure; Scale bar = 1 cm
图 5 MtATG7促进转基因拟南芥在碳胁迫下的生存图中为将苗龄3周的野生型Col、自噬突变体atg7以及转基因atg7/MtATG7#1、atg7/MtATG#2、MtATG7-OX#1、MtATG7-OX#2拟南芥植株黑暗处理7 d,恢复光照8 d后的表型;比例尺=1 cmFigure 5. MtATG7 promotes survival of transgenic Arabidopsis under carbon starvation3-week-old Arabidopsis seedlings of wild type Col, autophagy mutant atg7, and transgenic lines atg7/MtATG7#1, atg7/MtATG#2, MtATG7-OX#1, and MtATG7-OX#2 were treated in the dark for 7 days,and the phenotypes after 8 days of recovery under light are shown in the figure; Scale bar = 1 cm![]() 图 6 碳胁迫后的拟南芥幼苗的存活率统计各株系在恢复光照8 d后的存活率;柱子上方的不同小写字母表示各株系之间差异显著(P<0.05,LSD法)Figure 6. Survival of Arabidopsis seedlings after carbon stressThe survival rate of each strain after 8 days of light recovery are calculated; Different lowercase letters on bars indicate significant differences among strains (P<0.05,LSD test)
图 6 碳胁迫后的拟南芥幼苗的存活率统计各株系在恢复光照8 d后的存活率;柱子上方的不同小写字母表示各株系之间差异显著(P<0.05,LSD法)Figure 6. Survival of Arabidopsis seedlings after carbon stressThe survival rate of each strain after 8 days of light recovery are calculated; Different lowercase letters on bars indicate significant differences among strains (P<0.05,LSD test)2.5 MtATG7基因能够恢复拟南芥atg7突变体的自噬活性
GFP-ATG8e重组蛋白能够被自噬体降解,并释放游离的GFP,因此GFP/GFP-ATG8e比值是植物自噬活性的重要指标之一。为了探究MtATG7过表达植株是否通过自噬途径增强碳饥饿胁迫耐受性,将GFP-ATG8e、atg7/GFP-ATG8e以及转基因的atg7/MtATG7/GFP-ATG8e置于碳饥饿胁迫下,并检测GFP-ATG8e蛋白的剪切活性。结果显示atg7突变体在碳饥饿胁迫下,不能产生正常的游离的GFP条带,而转基因的atg7/MtATG7/GFP-ATG8e植株的GFP-ATG8剪切活性能够恢复到接近Col的水平(图7)。以上结果表明,MtATG7能够互补拟南芥内源AtATG7的功能,自噬活性的恢复很可能是碳饥饿胁迫耐受性提高的原因。
![]() 图 7 MtATG7使拟南芥atg7突变体的自噬活性恢复免疫印迹试验检测拟南芥GFP-ATG8e、atg7-3/GFP-ATG8e以及转基因MtATG7/atg7-3/GFP-ATG8e的GFP剪切活性;上样量对照为丽春红染色的Rubisco蛋白Figure 7. MtATG7 restores autophagy activity of atg7 mutant in ArabidopsisWestern blot was used to detect the GFP cleavage activities of GFP-ATG8e, atg7-3/ ATG8e and MtATG7/atg7-3/ATG8e; Rubisco protein with ponceau staining was used as the loading control
图 7 MtATG7使拟南芥atg7突变体的自噬活性恢复免疫印迹试验检测拟南芥GFP-ATG8e、atg7-3/GFP-ATG8e以及转基因MtATG7/atg7-3/GFP-ATG8e的GFP剪切活性;上样量对照为丽春红染色的Rubisco蛋白Figure 7. MtATG7 restores autophagy activity of atg7 mutant in ArabidopsisWestern blot was used to detect the GFP cleavage activities of GFP-ATG8e, atg7-3/ ATG8e and MtATG7/atg7-3/ATG8e; Rubisco protein with ponceau staining was used as the loading control2.6 过表达MtATG7抑制氮饥饿导致的叶片变黄
将苗龄11 d的MtATG7过表达植株转移至缺氮培养基上培养,结果表明,在缺氮处理后atg7和野生型相比叶片明显变黄,叶绿素含量下降(图8、9)。而互补植株atg7/MtATG7的叶绿素含量和野生型相近,说明MtATG7能够挽救atg7突变体的缺陷表型(图9)。其中atg7/MtATG7#1和MtATG7-OX#1两个株系在缺氮处理后,几乎所有植株的叶片仍然保持绿色,而野生型已有少量植株开始发紫(图8A、8B)。叶绿素测量结果也表明,atg7/MtATG7#1和MtATG7-OX#1两个株系的叶绿素含量和野生型相比有轻微增加(图9)。
![]() 图 8 MtATG7促进转基因拟南芥抵抗氮胁迫A:将苗龄11 d的野生型Col、自噬突变体atg7以及转基因atg7/MtATG7#1、atg7/MtATG#2、MtATG7-OX#1、MtATG7-OX#2拟南芥幼苗移入缺氮的1/2MS培养基中培养7 d后的表型;B:在氮胁迫7 d后分别对每个株系取2株幼苗拍照;比例尺=5 mmFigure 8. MtATG7 promotes resistance to nitrogen starvation in transgenic ArabidopsisA: Phenotypes for 11-day-old wild-type Col, autophagy mutant atg7, and transgenic atg7/MtATG7#1, atg7/MtATG#2, MtATG7-OX#1, MtATG7-OX#2 Arabidopsis seedlings after transferred to nitrogen-deficient 1/2MS medium and cultured for 7 days; B: After 7 days of nitrogen starvation stress, two seedlings from each line were photographed; Scale bar = 5 mm
图 8 MtATG7促进转基因拟南芥抵抗氮胁迫A:将苗龄11 d的野生型Col、自噬突变体atg7以及转基因atg7/MtATG7#1、atg7/MtATG#2、MtATG7-OX#1、MtATG7-OX#2拟南芥幼苗移入缺氮的1/2MS培养基中培养7 d后的表型;B:在氮胁迫7 d后分别对每个株系取2株幼苗拍照;比例尺=5 mmFigure 8. MtATG7 promotes resistance to nitrogen starvation in transgenic ArabidopsisA: Phenotypes for 11-day-old wild-type Col, autophagy mutant atg7, and transgenic atg7/MtATG7#1, atg7/MtATG#2, MtATG7-OX#1, MtATG7-OX#2 Arabidopsis seedlings after transferred to nitrogen-deficient 1/2MS medium and cultured for 7 days; B: After 7 days of nitrogen starvation stress, two seedlings from each line were photographed; Scale bar = 5 mm![]() 图 9 氮胁迫前后的拟南芥幼苗叶绿素含量数据为平均值±标准差, n=8;相同处理柱子上方的不同小写字母表示各株系之间差异显著(P<0.05,LSD法)Figure 9. Chlorophyll content of Arabidopsis seedlings before and after N stressData are means ± SDs, n=8; Different lowercase letters on bars of the same treatment indicate significant differences among strains (P<0.05,LSD test)
图 9 氮胁迫前后的拟南芥幼苗叶绿素含量数据为平均值±标准差, n=8;相同处理柱子上方的不同小写字母表示各株系之间差异显著(P<0.05,LSD法)Figure 9. Chlorophyll content of Arabidopsis seedlings before and after N stressData are means ± SDs, n=8; Different lowercase letters on bars of the same treatment indicate significant differences among strains (P<0.05,LSD test)3. 讨论与结论
碳元素是糖类物质和蛋白质分子的基础,对植物体内能量和物质的稳态至关重要。一方面,植物遭受短期碳饥饿时,细胞自噬能够降解储存于叶片中的瞬时淀粉,为细胞提供额外的能量以渡过逆境[21];另一方面,在长期碳饥饿时,细胞自噬能够降解叶绿体及其中的蛋白质,维持重要组织的氨基酸供应,从而提高植物的生存能力[22]。氮元素是氨基酸、核苷酸、叶绿素的重要组成成分,也是植物生存所必需的元素之一。在低氮条件下,细胞自噬能够将衰老叶片中的氮素分解再活化,使之能够被运输至新生叶片,实现生长发育效率的最大化;同时氮素的再活化也是种子成熟的关键步骤之一[23]。
本研究发现MtATG7蛋白具有保守的ATG7特征结构域,异源过表达MtATG7能够改善atg7突变体在碳/氮饥饿条件下的叶片早衰性状,而且在缺碳胁迫下MtATG7过表达植株的存活率显著提高,从野生型的55.5%提高至85.2%~92.6%。缺碳条件下,MtATG7能够使atg7突变体的自噬剪切活性恢复,说明MtATG7能够参与拟南芥自噬途径的调控。ATG7是植物自噬过程的限速酶之一,在自噬过程中分别和ATG12、ATG8结合,激活ATG8的脂化过程[24],因此MtATG7很可能通过提高植物自噬水平,使碳/氮饥饿的耐受能力提高。在拟南芥中,ATG7和许多叶片衰老标记基因,例如MYB2、NAP、SAG12、NYE1的表达存在明显关联性,表明ATG7也是植物叶片衰老调控的枢纽之一[25]。在碳/氮饥饿条件下,MtATG7过表达植株叶绿素含量下降的速度减慢,该结果暗示MtATG7还可能通过抑制衰老相关基因的表达,使植物叶片的衰老延迟。
虽然MtATG7基因在参与自噬过程当中具有保守性,但是在不同植物中,自噬途径所影响的生理过程仍然存在特殊性。例如在正常生长条件下,自噬途径的缺失对拟南芥生长和繁殖几乎没有影响;但是在水稻和烟草中,自噬途径受阻会导致其育性明显降低,结实率下降,同时水稻的抽穗时间明显延迟[26-27]。因此过表达MtATG7基因在苜蓿和拟南芥中对抗逆性状的影响是否存在差异,是今后值得探究的课题。
-
![]()
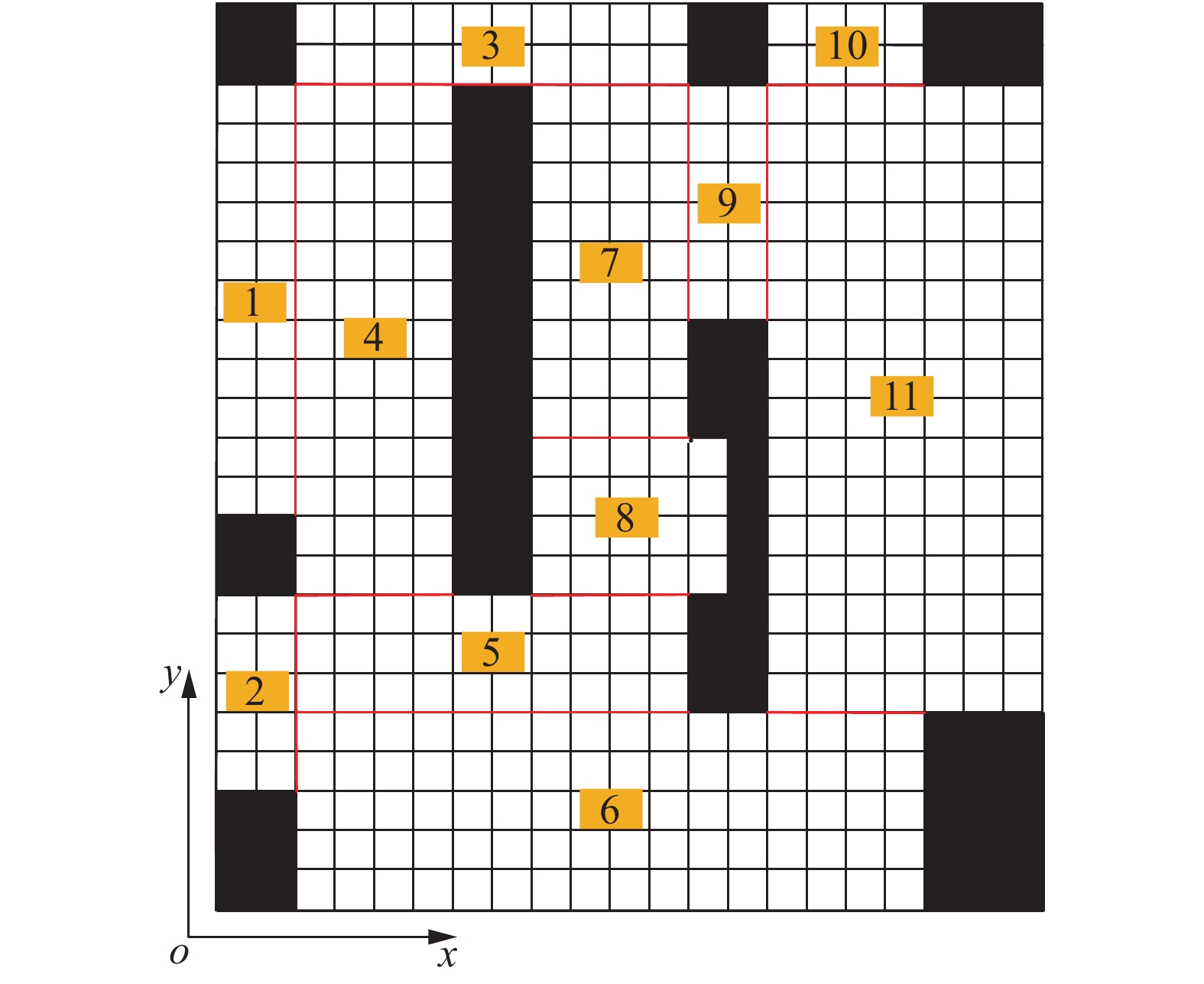
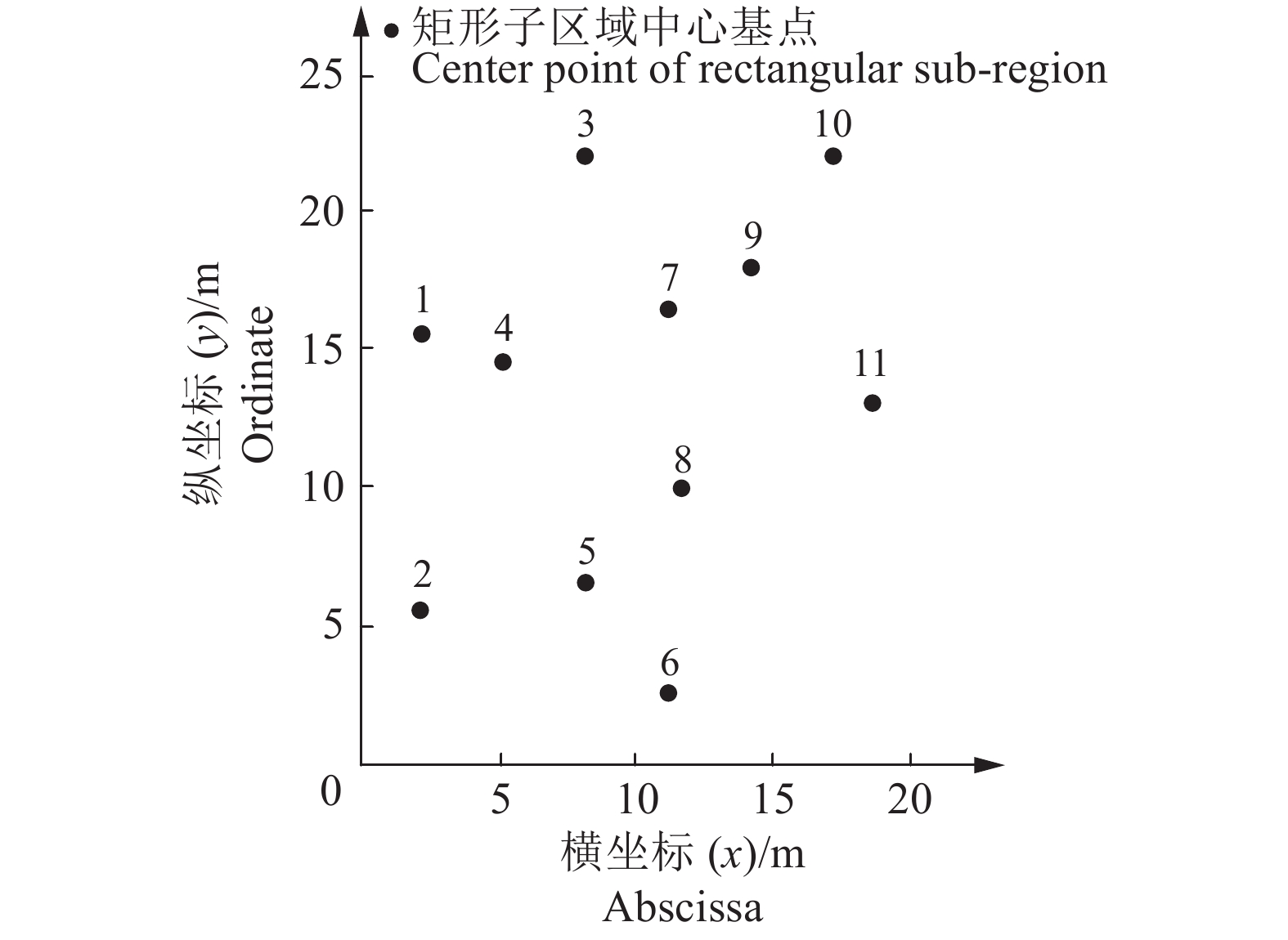
图 3 矩形子区域划分结果
图中数字为标号
Figure 3. Result of rectangular sub-region division
The figures in the diagram are labels
![]()
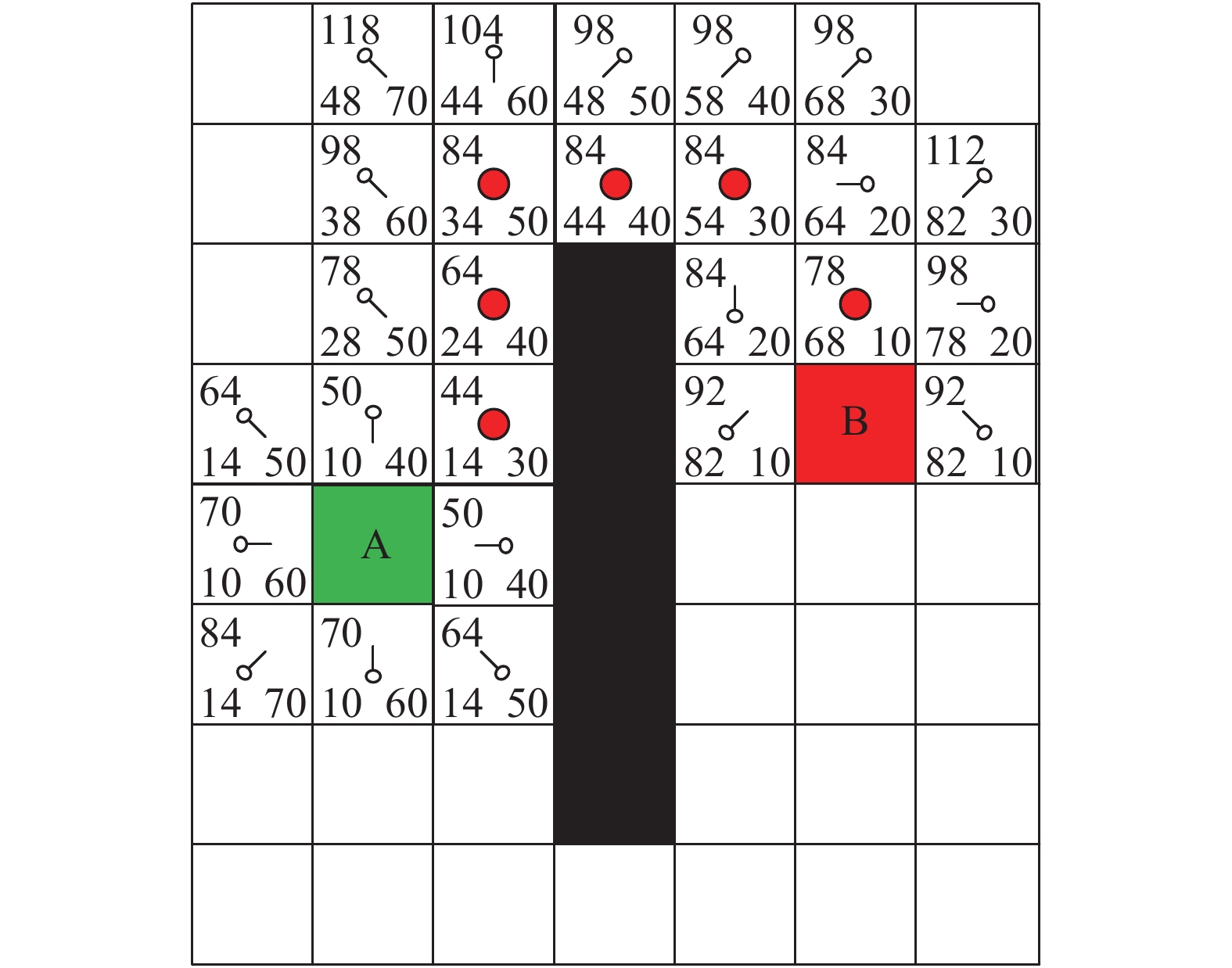
图 4 农业机器人A*算法路径搜索
红色圆点即从起点A到目标点B的最短路径;方格中左上角数字为 f 值,代表由起点A到目标点B的代价值;右下角数字为 h 值,代表从指定节点到终点B的估算成本;左下角数字为 g 值,代表农业机器人从起点A移动到指定节点的移动代价
Figure 4. Path search of agricultural robot by A* algorithm
The red dots are the shortest path from starting point A to target point B; The upper left number is f value, representing the cost value for moving from starting point A to target point B; The bottom right number is h value, representing the estimated cost for moving from an assigned node to target point B; The bottom left number is g value, representing the movement cost for moving from starting point A to an assigned node
![]()
图 7 遍历路径规划图
绿色圆点和红色五角星分别为整个遍历路径规划的起点和终点,蓝色线条为遍历规划路线
Figure 7. Traversal path planning
The green dot is the starting point and the red pentagram is the ending point of the whole planned traversal path, the blue line is the planned traversal path
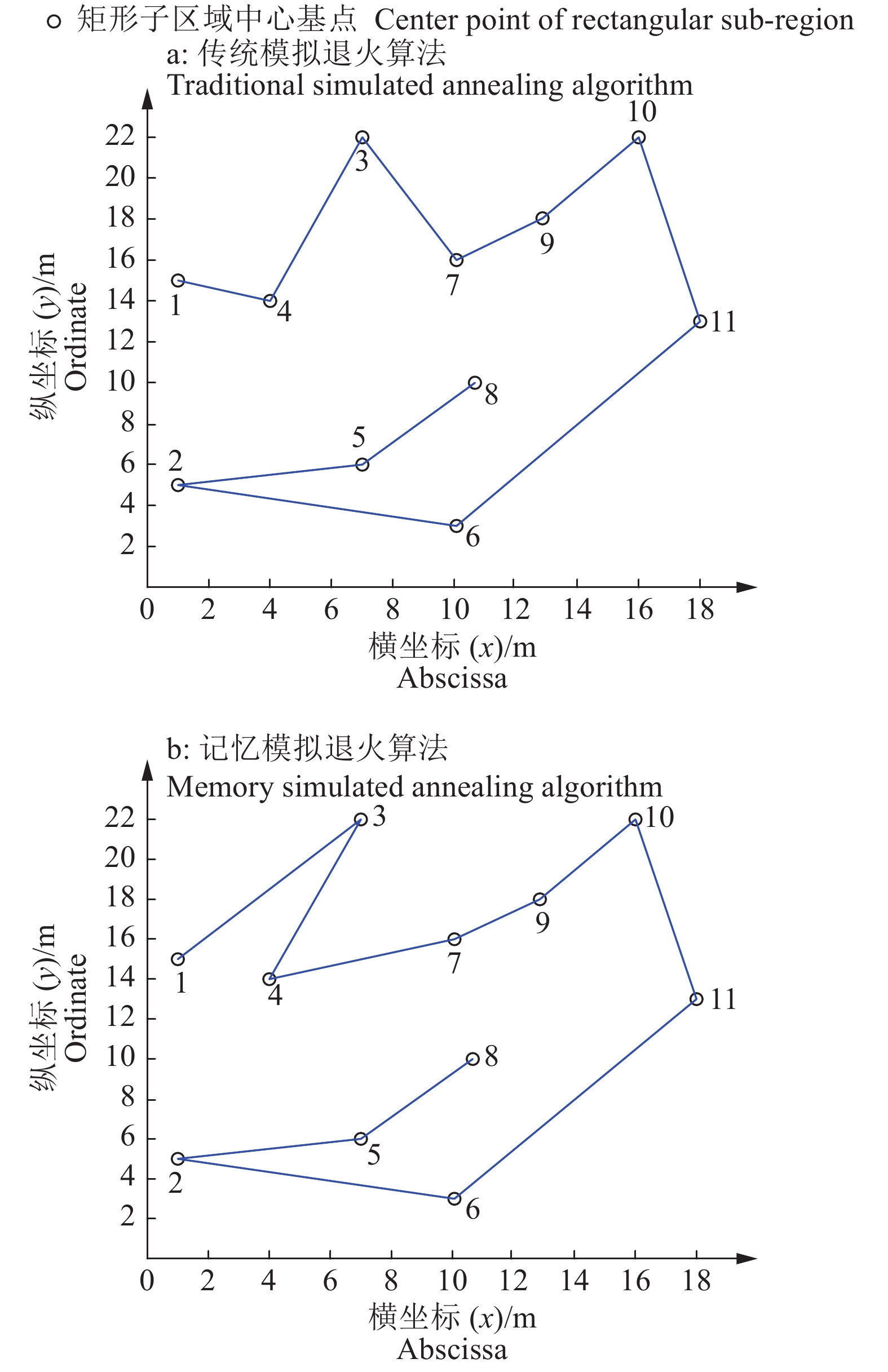
表 1 算法遍历顺序规划对比
Table 1 Comparison of traversal sequence planning in different algorithm
算法名称 Algorithm name 遍历顺序1) Traversal order 曼哈顿距离/m Manhattan distance 传统模拟退火算法
Traditional simulated annealing algorithm1-4-3-7-9-10-11-6-2-5-8 29.6 记忆模拟退火算法
Memory simulated annealing algorithm1-3-4-7-9-10-11-6-2-5-8 26.8 1) 数字表示矩形子区域中心基点
1)The figures are center points of rectangular sub-region 下载: 导出CSV
下载: 导出CSV
-
[1] 朱铁欣, 董桂菊, 颜丙学, 等. 基于改进蚁群算法的农业机器人路径规划研究[J]. 农机化研究, 2016, 38(9): 48-52. doi: 10.3969/j.issn.1003-188X.2016.09.009 [2] 邱雪娜, 刘士荣, 宋加涛, 等. 不确定动态环境下移动机器人的完全遍历路径规划[J]. 机器人, 2006(6): 586-592. doi: 10.3321/j.issn:1002-0446.2006.06.007 [3] 邱雪娜, 刘士荣, 俞金寿, 等. 移动机器人的完全遍历路径规划: 生物激励与启发式模板方法[J]. 模式识别与人工智能, 2006, 19(1): 122-128. doi: 10.3969/j.issn.1003-6059.2006.01.022 [4] 谢斌, 刘士荣, 俞金寿. 基于在线图搜索的移动机器人遍历运动规划[J]. 华东理工大学学报(自然科学版), 2007(4): 551-557. doi: 10.3969/j.issn.1006-3080.2007.04.021 [5] ZOU D X, WANG G G, PAN G, et al. A modified simulated annealing algorithm and an excessive area model for floorplanning using fixed-outline constraints[J]. Front Inform Tech El, 2016, 17(11): 1228-1244. doi: 10.1631/FITEE.1500386
[6] 路鹏, 周东岱, 钟绍春, 等. 基于模拟退火算法的计算机自适应测试项目选择方法研究[J]. 计算机应用与软件, 2012, 29(10): 175-179. [7] 杜利超, 钱桦, 肖爱平. 路径规划技术及其在大棚作业机器人中的应用[J]. 湖北农业科学, 2010, 49(5): 1205-1208. doi: 10.3969/j.issn.0439-8114.2010.05.058 [8] 史兵, 段锁林, 李菊, 等. 温室移动机器人复合栅格地图构建方法研究[J]. 计算机应用研究, 2019, 36(3): 824-828. [9] 张堂凯. 己知环境下智能清洁机器人路径规划研究[D]. 南京: 南京邮电大学, 2017. [10] QIAN J Y, ZHOU Z D, ZHAO L Z, et al. Accelerating reconfiguration for VLSI arrays with A‐star algorithm[J]. IEEJ T Electr Electr , 2018, 13(10): 1511-1519. doi: 10.1002/tee.22716
[11] 王维, 裴东, 冯璋. 改进A*算法的移动机器人最短路径规划[J]. 计算机应用, 2018, 38(5): 1523-1526. [12] 张欣欣, 薛金林. 基于云模型的农业移动机器人人机合作路径规划[J]. 华南农业大学学报, 2017, 38(6): 105-111. doi: 10.7671/j.issn.1001-411X.2017.06.016 [13] 张文, 刘勇, 张超凡, 等. 基于方向A*算法的温室机器人实时路径规划[J]. 农业机械学报, 2017, 48(7): 22-28. doi: 10.6041/j.issn.1000-1298.2017.07.003 [14] 孙秀巧, 王健, 巫威眺. 基于改进遗传退火算法的高速公路巡逻车路径优化调度[J]. 科学技术与工程, 2019, 19(21): 296-302. doi: 10.3969/j.issn.1671-1815.2019.21.045 [15] MIAO Z W, YANG F, FU K, et al. Transshipment service through crossdocks with both soft and hard time windows[J]. Ann Oper Res, 2012, 192(1): 21-47. doi: 10.1007/s10479-010-0780-4












计量
- 文章访问数: 1289
- HTML全文浏览量: 13
- PDF下载量: 1446
