Antibacterial activity and mechanism of garlic organic sulfide against Bacillus amyloliquefaciens
-
摘要:目的
明确大蒜有机硫化物(GOS)对豆制品优势腐败解淀粉芽孢杆菌Bacillus amyloliquefaciens的抑菌活性及作用机理。
方法通过抑菌圈法和二倍稀释法测定GOS对菌株的抑菌活性、最低抑菌浓度(MIC)和最低杀菌浓度(MBC),并分析其对细菌生长曲线及胞外蛋白酶活性的影响;分析GOS处理后解淀粉芽孢杆菌代谢活性、碱性磷酸酶活性、β–半乳糖苷酶活性及胞外溶液电导率的变化,并结合扫描电镜观察菌体形态变化,探讨GOS对解淀粉芽孢杆菌的抑菌机理。
结果3 g/mL 的GOS对解淀粉芽孢杆菌DY1a和DY1b的抑菌圈直径分别为31.7和25.7 mm,MIC均为30 mg/mL,MBC分别为240和480 mg/mL。30 mg/mL GOS对菌株DY1a和DY1b胞外蛋白酶活性的抑制率分别为47.3%和15.6%,使细菌细胞代谢活性(D490 nm)分别下降了2.771和4.091,胞外碱性磷酸酶活性分别提高了0.029和0.036 U/mL,胞外β−半乳糖苷酶活性(D420 nm)分别提高了0.047与0.016,胞外溶液电导率分别提高了0.060和0.031 mS/cm。扫描电镜结果显示,GOS处理后的菌株细胞表面出现皱缩、孔洞式塌陷和破裂。
结论大蒜有机硫化物对解淀粉芽孢杆菌抑制作用明显,其抑菌机理与影响细菌细胞壁、细胞膜的通透性和完整性,干扰细菌生理代谢活动有关。
Abstract:ObjectiveTo explore antibacterial activity and mechanism of garlic organic sulfide (GOS) against Bacillus amyloliquefaciens.
MethodThe antimicrobial activity, minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) of GOS against B. amyloliquefaciens strain were determined using the inhibition zone method and the double dilution method, and its inhibitory effects on bacterial growth curve and extracellular protease activity were evaluated. Metabolic activity, alkaline phosphatase activity, β-galactosidase activity, extracellular conductivity and thallus morphological changes observed through scanning electron microscope of B. amyloliquefaciens treated with GOS were investigated for elucidating antibacterial mechanism of GOS against B. amyloliquefaciens.
ResultDiameter of inhibitory zone of GOS against B. amyloliquefaciens DY1a and DY1b were 31.7 and 25.7 mm respectively, MIC of both strains were 30 mg/mL, MBC were 240 and 480 mg/mL, respectively. The inhibition rate of 30 mg/mL GOS on DY1a and DY1b extracellular protease activity were 47.3% and 15.6% respectively. After treatment with 30 mg/mL GOS, metabolic activity (D490 nm) decreased by 2.771 and 4.091 respectively, extracellular alkaline phosphatase activity increased by 0.029 and 0.036 U/mL, extracellular β-galactosidase activity (D420 nm) increased by 0.047 and 0.016 respectively and the extracellular electrical conductivity increased by 0.060 and 0.031 mS/cm respectively. The results of SEM showed that the surface of bacteria cells treated with GOS had shrinkage, cavity collapse and rupture.
ConclusionThe excellent antibacterial effect of GOS against B. amyloliquefaciens may be attributed to irreversible damage to cell wall and cell membrane and alterations of cell physiological metabolism.
-
番茄Solanum lycopersicum因含有丰富的营养物质而成为我国重要的蔬菜作物,也是研究果实发育、成熟的重要模式植物[1-2]。番茄种植过程中易受病原菌侵害,造成减产,细菌性斑点病、细菌性溃疡病、细菌性枯萎病等细菌性病害是造成番茄减产的重要原因[3]。丁香假单胞菌番茄致病变种(Pseudomonas syringae pv. tomato DC3000,Pst DC3000)是番茄细菌性斑点病的致病变种之一,也是研究植物−病原菌互作的模式菌株[4]。Pst DC3000吸附到番茄叶片表面,从气孔或伤口侵入,引起局部坏死、周围弥漫性黄化,危害番茄叶片、果实、花、茎秆等,给番茄产业造成严重的损失。
茉莉酸(Jasmonic acid,JA)是一类广泛存在于植物中的内源激素,1971年在一种真菌培养过滤液中获得[5]。研究发现,外源JA具有促进植物衰老和调控生长的作用,且昆虫啃咬和真菌侵染均可以促进JA和茉莉酸甲酯(Methyl jasmonate,MeJA)的积累[5-6]。植物在逆境条件下,积累不同形式的JA,最终生成活性的茉莉酸−异亮氨酸(Jasmonic acid-isoleucine,JA-Ile),JA-Ile诱导JA受体F-box CORONATINE INSENSITIVE1(COI1)与Jasmonate ZIM-domain(JAZ)蛋白形成共受体复合体,促使JAZ蛋白被泛素化降解,激活转录因子下游抗病反应基因的表达[7-8]。例如SlJAZ25基因在番茄对灰叶斑病的抗性反应中起负调控作用[9]。通过病毒沉默番茄JAZ3基因发现番茄对叶霉病的抵抗能力降低,同时,植株内源JA含量降低,水杨酸(Salicylic acid,SA)含量升高[10]。AtJAZ7基因的突变增加了拟南芥对尖孢镰刀菌和Pst DC3000的感病性[11]。SlJAZ2、SlJAZ6、SlJAZ7瞬时沉默植株对Pst DC3000的敏感性增加[12]。Pst DC3000的致病因子冠菌素与JA-Ile在结构上存在很大的相似性,冠菌素通过模仿JA-Ile,激活JA信号传导,抑制SA介导的防御信号[12]。拟南芥ANAC032转录因子与MYC2、NIMIN1和PDF1.2启动子结合,激活SA介导的防御反应但抑制JA反应基因的表达,提高对Pst DC3000的抗性[13],说明JA和SA之间拮抗或协同的关系可帮助植物抵御外界不良环境[14]。
植物TGA转录因子(TGACG-binding transcription factor)是SA抗病途径的重要调节因子,也是JA/乙烯抗病信号途径中的重要组成部分,发挥重要作用[15-16]。拟南芥TGA转录因子家族共有10个基因,分为5组,每组都具有不同的功能。TGA与病程相关(Pathogenesis related,PR)蛋白基因启动子上TGACG(as-1)元件结合调节下游病程相关基因的转录水平,从而提高植物的抗病性[17-19]。研究表明TGA转录因子I、II、III组与SA受体NON-EXPRESSOR of PR-1(NPR1)均存在不同强度的互作[17-19]。在JA/ETH诱导下tga2/5/6 3份突变体中PDF1.2和b-CHI表达被抑制,使得番茄对坏死性番茄灰霉病更易感[20]。从大豆基因组中鉴定出的GmTGA8和GmTGA19明显抑制大豆花叶病毒的增殖,对大豆花叶病具有明显基础抗性[21]。在猕猴桃中瞬时过表达AcTGA01、AcTGA06、AcTGA07、AcTGA09有利于增强猕猴桃对细菌性溃疡病的抗病性,其中瞬时过表达AcTGA7的猕猴桃叶片中细菌积累量最少[22]。JAZ和TGA转录因子分别是JA和SA途径的重要调控因子,JAZ3基因瞬时沉默可以同时影响JA和SA的含量,推测JAZ和TGA间可能有一定关联[10]。
番茄13个JAZ家族成员中,SlJAZ1~12基因均受到Pst DC3000的诱导,其中SlJAZ7在Pst DC3000病原菌诱导3 h的表达量显著上调,瞬时沉默SlJAZ7基因后植株感病性增加[12],然而其中的分子机制尚未阐明。为进一步探索SlJAZ7在番茄抗病性中的调控机制,本研究首先通过转基因技术获得过表达SlJAZ7基因的转基因番茄,接种Pst DC3000验证其抗病功能,通过RT-qPCR明确SlJAZ7响应Pst DC3000、JA及SA处理,通过Y2H等试验探索SlJAZ7与SlTGA7的关系,这将为进一步研究SlJAZ7基因的作用机理和防治番茄细菌性斑点病提供理论基础。
1. 材料与方法
1.1 材料种植与试剂
野生小果型番茄‘Micro-Tom’(MT)和本氏烟草由海南大学海南省热带生物资源可持续利用重点实验室、热带作物生物育种国家重点实验室(本实验室)保存,种植于26 ℃温室中,培养条件为16 h光照/8 h黑暗。
2× Taq PCR试剂(目录号:KT201-02)、多糖多酚植物总RNA提取试剂盒(目录号:DP441)购自天根生化科技(北京)有限公司;反转录试剂盒(目录号:D2639A)和SYBR Premix Ex Taq Kit(目录号:DRR081A)、Clontech酵母双杂交试剂盒(目录号:630489)及酵母培养基购自宝生物有限公司;PCR产物回收纯化试剂盒(目录号:DR02)、质粒提取试剂盒(目录号:PL03)购自艾德莱生物有限公司;大肠埃希菌感受态DH5α、农杆菌感受态LBA4404和酵母Y2H Gold感受态细胞购自上海唯地生物技术有限公司;植物表达载体pGAMBIA1300-eGFP由本实验室改造保存。
1.2 SlJAZ7基因克隆
在Phytozome 13(https://phytozome-next.jgi.doe.gov/)数据库中选择番茄数据库S. lycopersicum ITAG4.0,根据基因序列号Solyc11g011030搜索得到SlJAZ7的基因序列,设计全长引物(表1),利用PCR扩增得到SlJAZ7全长片段。
表 1 本研究所用引物Table 1. Primers used in this study引物名称
Primer name引物序列1) (5′→3′)
Primer sequence目的
PurposeSlJAZ7-NC-F agtggtctctgtccagtcctATGGATTCAAGAATGGAGATAGATT 基因克隆 SlJAZ7-NC-R ggtctcagcagaccacaagtGTTTTCCCAATGAACGCTTGAC 基因克隆 SlJAZ7-EcoRI-F ccggaattcATGGATTCAAGAATGGAGATAGATT 互作关系验证 SlJAZ7-BamHI-R cgggatccGTTTTCCCAATGAACGCTTGAC 互作关系验证 SlTGA7-EcoRI-F ccggaattcATGAACATGAGTTCTACATCAACTC 互作关系验证 SlTGA7-BamHI-R cgggatccGGTAGGTTCACGAGGACGAG 互作关系验证 SlJAZ7-SalI-F acgcgtcgacATGGATTCAAGAATGGAGATAGATT BiFC试验 SlTGA7-SalI-F acgcgtcgacATGAACATGAGTTCTACATCAACTC BiFC试验 SlJAZ7-Q-F ATTATGCTAAAGGAGCACTTGCTAT RT-qPCR SlJAZ7-Q-R CGCTTGACGACGCCG RT-qPCR Sltublin-Q-F TTGGTTTTGCACCACTGACTTC RT-qPCR Sltublin-Q-R AAGCTCTGGCACTCTCAAAGC RT-qPCR 1)带下划线的碱基代表酶切位点序列
1) The underlined bases represent the sequences of enzyme cleavage site1.3 SlJAZ7过表达番茄植株创制及鉴定
将MT番茄种子消毒后培养7~10 d长出2片完整子叶,切下子叶,放入D600 nm=0.35~0.40的农杆菌悬浮液中侵染15 min,然后于20~22 ℃下避光共培养2 d。将外植体转移到筛选培养基诱导愈伤,一定时间后转入再生培养基中,待长出4~5 cm不定芽,转入生根培养基中。将再生苗移栽至装有基质的营养钵中培养成苗。
为鉴定SlJAZ7的转基因阳性植株,采集野生型植株(WT)及转基因植株(OE)的叶片,利用RT-qPCR,以SlTublin (Solyc10g086760)为内参基因,参考宝生物公司SYBR Premix Ex Taq II Kit说明书操作完成RT-qPCR反应,检测SlJAZ7在转基因植株的表达量。
1.4 Pst DC3000处理对照和转基因番茄
Pst DC3000菌液于KB平板(25 µg/mL利福平)上划线纯化,挑取单菌落,使用10 mmol/L氯化镁溶液稀释,使悬浮液D600 nm为0.2,加入0.025%(φ)的表面活性剂,均匀喷洒至野生型植株(WT)叶片和过表达SlJAZ7植株(SlJAZ7-OE)叶片正反面,23 ℃暗培养1 d后转入26 ℃培养房中正常培养3 d,拍照保存。
1.5 DAB染色
使用纯水溶解DAB粉末,配置终质量浓度为1 mg/mL的DAB染色液,将叶片浸泡至染色液中,于锡箔纸中避光染色4 h。按照无水乙醇、甘油、冰醋酸的体积比为3∶1∶1的比例配制成脱色液。将染色后的叶片放入脱色液中煮沸15 min,至叶片底色变白,弃脱色液,超纯水清洗多次,拍照保存。
1.6 番茄样本处理与采集
为了研究SlJAZ7和SlTGA7基因的组织表达模式,采集种植45 d MT番茄的根、茎、叶、花和不同时期果实的样品,液氮保存,备用。为了研究SlJAZ7和SlTGA7基因对不同胁迫的响应模式,生长期45 d左右、生长势一致的MT幼苗被用于胁迫处理试验,将2 mmol/L SA、100 µmol/L MeJA、D600 nm为0.2的Pst DC3000菌液喷施在叶片背面,于0、0.25、0.5、1、2、4、6、24 h采集植株叶片。提取RNA并反转录获得cDNA作为模板,以SlTublin (Solyc10g086760)为内参基因,参考宝生物公司SYBR Premix Ex Taq II Kit说明书操作完成RT-qPCR反应,仪器为Step-One Plus (ABI,USA),利用2−ΔΔCt计算相对表达量,使用GraphPad Prism 8.0.1软件绘图。
1.7 亚细胞定位分析
将去掉终止密码子的SlJAZ7和SlTGA7基因的CDS序列重组至pGAMBIA1300:eGFP载体中,构建35S:SlJAZ7:eGFP:pGAMBIA1300、35S:SlTGA7:eGFP:pGAMBIA1300植物表达载体。以pGAMBIA1300:eGFP为对照,将测序正确的质粒转化农杆菌感受态细胞LBA4404,挑取单菌落活化至D600 nm为0.4,浓缩至D600 nm在0.8左右,加入终浓度200 µmol/L的乙酰丁香酮,混匀后室温避光静置3 h,注射烟草叶片下表皮细胞,48 h后观察绿色荧光蛋白的亚细胞定位。
1.8 Y2H验证SlJAZ7和SlTGA7蛋白互作关系
将SlTGA7-pGBKT7质粒与SlJAZ7-pGADT7质粒共转至酵母感受态Y2H Gold中,涂布酵母培养基DDO(SD/-Trp-Leu)上,挑取单菌落,用pGADT7和pGBKT7的通用引物进行PCR鉴定,得到阳性菌株后,用9 g/L氯化钠溶液重悬并调整菌液D600 nm为0.002,9 g/L氯化钠溶液稀释不同的浓度梯度,点至DDO/X/A(SD/-Trp-Leu/X-α-Gal/AbA)、QDO/X/A(SD/-Trp-Leu-His-Ade/X-α-Gal/AbA)平板上培养3~5 d,观察菌斑是否变蓝,以验证两者是否存在互作。
1.9 BiFC验证互作关系
通过Sal I和BamH I内切酶将已去掉终止密码子的SlJAZ7和SlTGA7基因CDS片段分别重组得到SlJAZ7:nYFP、SlTGA7:cYFP载体,转化至农杆菌感受态GV3101(pSoup-p19),菌液经PCR鉴定正确后,将两者与核定位标记基因的农杆菌按照1∶1∶1的体积比混合均匀注射至烟草叶背面。3 d后在共聚焦显微镜下观察荧光。
1.10 pull down试验验证蛋白互作关系
将番茄SlJAZ7基因重组至pet28(a)-sumo-flag、SlTGA7重组至pet28(a)-sumo-strep构建原核表达载体(原核表达载体由本实验室改造保存),经测序正确后,转入大肠埃希菌感受态Rosetta (DE3)中大量表达蛋白,使用IPTG诱导提取细菌蛋白,BCA法测定蛋白浓度[23],经Western blot和考马斯亮蓝染色后检测蛋白质表达量。Input组A、B设置为等比例混合的SlJAZ7: pet28(a)-sumo-flag和SlTGA7: pet28(a)-sumo-strep、SlJAZ7: pet28(a)-sumo-flag和pet28(a)-sumo-strep空载蛋白,IP组设置为使用带有flag抗体的磁珠下拉处理过的上述蛋白,经SDS-PAGE验证后分别使用flag和strep抗体孵育,检测flag和strep标签的存在。
1.11 数据统计方法
定量数据均设计3次技术重复,数据以平均值±标准误表示,使用Excel表格对数据进行整理分析,使用公式2−ΔΔCt计算相对表达量,以t检验作为差异显著性分析方法,使用GraphPad Prism 8.0.1软件绘图。
2. 结果与分析
2.1 过表达SlJAZ7番茄的鉴定及表型分析
本研究利用农杆菌介导的番茄遗传转化体系,创制稳定遗传的转基因番茄植株,并通过RT-qPCR鉴定获得6株阳性株系。所有过表达株系SlJAZ7-OE中,SlJAZ7的表达量均显著高于野生型(WT),其中SlJAZ7基因在OE5株系表达量最高(图1)。为研究SlJAZ7基因在防御番茄细菌性斑点病中的功能,对WT和SlJAZ7-OE接种Pst DC3000,结果显示,接种4 d后,WT和SlJAZ7-OE叶片均产生黄色病斑,但是,WT病斑的面积更大(图2A)。
![图 1 转基因阳性植株SlJAZ7相对表达量]() 图 1 转基因阳性植株SlJAZ7相对表达量WT:野生型;OE1、5、6、7、12、14:过表达株系;“*”“**”分别表示过表达株系与野生型在P < 0.05和P < 0.01水平差异显著(t检验)Figure 1. Relative expression level of SlJAZ7 in transgenic positive plantsWT: Wild type; OE1, 5, 6, 7, 12, 14: Overexpressed lines; “*” and “**” indicate signidicant differences between overexpressed lines and wild type at P < 0.05 and P < 0.01 levels (t test)
图 1 转基因阳性植株SlJAZ7相对表达量WT:野生型;OE1、5、6、7、12、14:过表达株系;“*”“**”分别表示过表达株系与野生型在P < 0.05和P < 0.01水平差异显著(t检验)Figure 1. Relative expression level of SlJAZ7 in transgenic positive plantsWT: Wild type; OE1, 5, 6, 7, 12, 14: Overexpressed lines; “*” and “**” indicate signidicant differences between overexpressed lines and wild type at P < 0.05 and P < 0.01 levels (t test)由于Pst DC3000侵染会引起活性氧的积累,导致植物细胞膜系统损伤。本研究使用DAB染色检测接种4 d后番茄叶片中活性氧的积累。结果发现,DAB染色后WT叶片的黄褐色沉淀比SlJAZ7-OE多而深,说明WT较SlJAZ7-OE积累了更多的活性氧(图2B)。以上结果表明过表达SlJAZ7基因减少了Pst DC3000接种后番茄的活性氧积累,减轻了转基因番茄的病症。
![图 2 Pst DC3000病原菌侵染野生型(WT)和过表达株系(OE)后的表型(A)及DAB染色结果(B)]() 图 2 Pst DC3000病原菌侵染野生型(WT)和过表达株系(OE)后的表型(A)及DAB染色结果(B)Figure 2. Phenotype (A) and DAB staining results (B) after Pst DC3000 pathogens infection on wild type (WT) and overexpressed line (OE)
图 2 Pst DC3000病原菌侵染野生型(WT)和过表达株系(OE)后的表型(A)及DAB染色结果(B)Figure 2. Phenotype (A) and DAB staining results (B) after Pst DC3000 pathogens infection on wild type (WT) and overexpressed line (OE)2.2 番茄SlTGA7和SlJAZ7基因表达模式的比较
为了探究TGA家族与JAZ家族基因间的关系,前期通过RT-qPCR研究了番茄TGA转录因子家族13个成员对Pst DC3000、SA和JA处理的响应模式,结果显示SlTGA7 (Solyc12g056860)对Pst DC3000、SA和MeJA都有响应。在Pst DC3000和MeJA处理下,SlTGA7表达被显著抑制,而在SA处理下,SlTGA7表达显著上调,在6 h时表达量达到最高(图3A~3C)。与SlTGA基因的表达模式不同,在Pst DC3000处理0~24 h内,SlJAZ7基因的表达量显著上调。在MeJA处理过程中SlJAZ7基因表达量除6 h显著下调外,其余时间点均显著上调。在SA处理0.5、4 h时,SlJAZ7基因表达量显著上调,2、6~24 h时,SlJAZ7表达量显著下调(图3D~3F)。
![图 3 Pst DC3000、MeJA和SA处理下SlTGA7 (A、B、C)和SlJAZ7 (D、E、F)的表达模式]() 图 3 Pst DC3000、MeJA和SA处理下SlTGA7 (A、B、C)和SlJAZ7 (D、E、F)的表达模式“*”“**”分别表示其他时间点与0 h在P < 0.05和P < 0.01水平差异显著(t检验)Figure 3. Expression patterns of SlTGA7 (A, B, C) and SlJAZ7 (D, E, F) under Pst DC3000, MeJA and SA treatments“*” and “**” indicate significant differences between other time points and 0 h at P < 0.05 and P < 0.01 respectively (t test)
图 3 Pst DC3000、MeJA和SA处理下SlTGA7 (A、B、C)和SlJAZ7 (D、E、F)的表达模式“*”“**”分别表示其他时间点与0 h在P < 0.05和P < 0.01水平差异显著(t检验)Figure 3. Expression patterns of SlTGA7 (A, B, C) and SlJAZ7 (D, E, F) under Pst DC3000, MeJA and SA treatments“*” and “**” indicate significant differences between other time points and 0 h at P < 0.05 and P < 0.01 respectively (t test)两者的组织特异性表达分析结果显示,SlTGA7基因在成熟红果中的表达量最高,在根、茎和老叶等营养器官中也高表达(图4A);说明SlTGA7可能参与番茄生殖和营养生长。SlJAZ7在茎中的表达量最高,在老叶、幼叶、根中次之,在果和花中表达量最低,在营养器官中总体表达水平要高于生殖器官(图4B),这表明SlJAZ7可能主要在营养器官中发挥功能。利用RT-qPCR检测SlTGA7基因在SlJAZ7过表达株系中的表达量,结果发现SlJAZ7过表达后,SlTGA7基因的表达量均显著高于野生型(图5);说明SlJAZ7在转录水平的变化可以影响SlTGA7的转录水平。
![图 4 SlTGA7和SlJAZ7在不同器官中的表达模式]() 图 4 SlTGA7和SlJAZ7在不同器官中的表达模式1:青熟期果实,2:绿熟期果实,3:破色期果实,4:成熟红果,5:根,6:茎,7:幼叶,8:成熟叶,9:老叶,10:未完全展开的花;“*”“**”分别表示其他器官与青熟期果实在P < 0.05、P < 0.01水平差异显著(t检验)Figure 4. Tissue specific expression profile of SlTGA7 and SlJAZ71: Fruit at cyan ripening stage, 2: Fruit at green ripening stage, 3: Fruit at green colour breaking stage, 4: Mature red fruit, 5: Root, 6: Stem, 7: Young leaf, 8: Mature leaf, 9: Old leaf, 10: Not fully unfolded flower; “*” and “**” indicate significant differences between other organs and fruits at cyan ripening stage at P < 0.05 and P < 0.01 levels respectively (t test)
图 4 SlTGA7和SlJAZ7在不同器官中的表达模式1:青熟期果实,2:绿熟期果实,3:破色期果实,4:成熟红果,5:根,6:茎,7:幼叶,8:成熟叶,9:老叶,10:未完全展开的花;“*”“**”分别表示其他器官与青熟期果实在P < 0.05、P < 0.01水平差异显著(t检验)Figure 4. Tissue specific expression profile of SlTGA7 and SlJAZ71: Fruit at cyan ripening stage, 2: Fruit at green ripening stage, 3: Fruit at green colour breaking stage, 4: Mature red fruit, 5: Root, 6: Stem, 7: Young leaf, 8: Mature leaf, 9: Old leaf, 10: Not fully unfolded flower; “*” and “**” indicate significant differences between other organs and fruits at cyan ripening stage at P < 0.05 and P < 0.01 levels respectively (t test)![图 5 SlJAZ7过表达株系叶片中SlTGA7的表达模式]() 图 5 SlJAZ7过表达株系叶片中SlTGA7的表达模式WT:野生型,OE1、5、6、7、12、14:过表达株系;“*”“**”分别表示过表达株系与野生型在P < 0.05和P < 0.01水平差异显著(t检验)Figure 5. Expression pattern of SlTGA7 in the leaves of SlJAZ7 overexpressed strainsWT: Wild type, OE1, 5, 6, 7, 12, 14: Overexpressed lines; “*” and “**” indicate signidicant differences between overexpressed lines and wild type at P < 0.05 and P < 0.01 levels (t test)
图 5 SlJAZ7过表达株系叶片中SlTGA7的表达模式WT:野生型,OE1、5、6、7、12、14:过表达株系;“*”“**”分别表示过表达株系与野生型在P < 0.05和P < 0.01水平差异显著(t检验)Figure 5. Expression pattern of SlTGA7 in the leaves of SlJAZ7 overexpressed strainsWT: Wild type, OE1, 5, 6, 7, 12, 14: Overexpressed lines; “*” and “**” indicate signidicant differences between overexpressed lines and wild type at P < 0.05 and P < 0.01 levels (t test)2.3 SlJAZ7和SlTGA7蛋白亚细胞定位
通过注射烟草下表皮细胞,使用共聚焦显微镜观察。35S:eGFP:pGAMBIA1300蛋白是质膜共定位,而SlJAZ7和SlTGA7蛋白均在细胞核中呈现绿色荧光信号(图6),与核定位Marker的红色荧光信号重叠后呈现黄色荧光,表明SlJAZ7和SlTGA7蛋白定位于细胞核中,主要在细胞核中行使功能。
![图 6 番茄SlJAZ7和SlTGA7的亚细胞定位]() 图 6 番茄SlJAZ7和SlTGA7的亚细胞定位Figure 6. Subcellular localization of SlJAZ7 and SlTGA7 in tomato
图 6 番茄SlJAZ7和SlTGA7的亚细胞定位Figure 6. Subcellular localization of SlJAZ7 and SlTGA7 in tomato2.4 Y2H验证SlJAZ7与SlTGA7的互作关系
通过Y2H点对点试验验证SlJAZ7蛋白与SlTGA7蛋白的互作关系。如图7所示,pGADT7-SlJAZ7与pGBKT7-SlTGA7在DDO/X/A和QDO/X/A平板上和阳性对照一样,正常生长且变蓝,而阴性对照没有生长、变蓝,表明SlJAZ7和SlTGA7蛋白可能存在互作。
![图 7 Y2H试验验证番茄SlJAZ7和SlTGA7蛋白的互作关系]() 图 7 Y2H试验验证番茄SlJAZ7和SlTGA7蛋白的互作关系pGADT7-T/pGBKT7-Lam为阴性对照;pGADT7-T/pGBKT7-53为阳性对照;0、10−1、10−2、10−3表示菌液的稀释梯度Figure 7. Interaction between SlJAZ7 and SlTGA7 proteins verified by Y2HpGADT7-T/pGBKT7-Lam is negative control; pGADT7-T/pGBKT7-53 is positive control; 0, 10−1, 10−2, 10−3 represent the dilution gradients of bacterial solution
图 7 Y2H试验验证番茄SlJAZ7和SlTGA7蛋白的互作关系pGADT7-T/pGBKT7-Lam为阴性对照;pGADT7-T/pGBKT7-53为阳性对照;0、10−1、10−2、10−3表示菌液的稀释梯度Figure 7. Interaction between SlJAZ7 and SlTGA7 proteins verified by Y2HpGADT7-T/pGBKT7-Lam is negative control; pGADT7-T/pGBKT7-53 is positive control; 0, 10−1, 10−2, 10−3 represent the dilution gradients of bacterial solution2.5 BiFC验证SlJAZ7与SlTGA7的互作关系
分别将SlJAZ7:nYFP、SlTGA7:cYFP菌液与核定位Marker等比例混合后共注射至烟草中,于共聚焦显微镜的叠加场下观察到细胞核中存在黄色荧光信号,与阳性对照nYFP、cYFP载体一致。但是,SlJAZ7:nYFP/cYFP、SlTGA7:cYF/nYFP无绿色荧光,这进一步证明SlJAZ7和SlTGA7蛋白可能在细胞核中共同发挥功能(图8)。
![图 8 BiFC验证SlJAZ7和SlTGA7蛋白的互作关系(标尺:20 μm)]() 图 8 BiFC验证SlJAZ7和SlTGA7蛋白的互作关系(标尺:20 μm)Figure 8. Interaction between SlJAZ7 and SlTGA7 verified by BiFC (scale: 20 μm)
图 8 BiFC验证SlJAZ7和SlTGA7蛋白的互作关系(标尺:20 μm)Figure 8. Interaction between SlJAZ7 and SlTGA7 verified by BiFC (scale: 20 μm)2.6 pull down验证SlJAZ7与SlTGA7互作关系
将构建的原核表达载体SlJAZ7-pet28(a)-sumo-flag和SlTGA7-pet28(a)-sumo-strep通过细菌表达蛋白后,经SDS-PAGE电泳检测蛋白相对分子质量均正确,分别为41 280、51 590(图9)。使用Flag磁珠处理IP组,一抗分别使用flag和strep抗体进行孵育,结果发现,在试验组和阴性对照组的Input组和IP组中均检测到flag标签蛋白,同时试验组的Input和IP组也检测到strep标签,但在阴性对照IP组中不存在strep标签蛋白,进一步证明SlJAZ7和SlTGA7蛋白存在互作关系(图10)。
![图 9 考马斯亮蓝染色检测SlJAZ7与SlTGA7蛋白的分子量]() 图 9 考马斯亮蓝染色检测SlJAZ7与SlTGA7蛋白的分子量Figure 9. Relative molecular masses of SlJAZ7 and SlTGA7 determined by Kaumas bright blue staining
图 9 考马斯亮蓝染色检测SlJAZ7与SlTGA7蛋白的分子量Figure 9. Relative molecular masses of SlJAZ7 and SlTGA7 determined by Kaumas bright blue staining![图 10 Pull down试验验证SlJAZ7与SlTGA7的互作关系]() 图 10 Pull down试验验证SlJAZ7与SlTGA7的互作关系Figure 10. Interaction between SlJAZ7与SlTGA7 verified by pull down experiment
图 10 Pull down试验验证SlJAZ7与SlTGA7的互作关系Figure 10. Interaction between SlJAZ7与SlTGA7 verified by pull down experiment3. 讨论与结论
3.1 讨论
本研究通过转基因番茄的抗病表型试验,发现Pst DC3000处理后SlJAZ7-OE番茄叶片积累的H2O2更少,说明过表达SlJAZ7后番茄的抗病性增强。这与前期瞬时沉默SlJAZ7基因后植株感病性增强的研究结果一致[12],再次验证了SlJAZ7基因正调控番茄对细菌性斑点病抗性的结论。本研究发现SlTGA7基因在番茄SlJAZ7过表达株系中均上调表达,证实了SlJAZ7和SlTGA7基因可能存在协同作用的推测。番茄TGA转录因子通过与SA受体及参与植物免疫的主要调节因子NPR1相互作用,调节防御相关基因的表达来发挥作用[24]。拟南芥AtTGA1和AtTGA4与乙烯响应因子ERF互作调控植物抗逆性[25]。拟南芥AtTGA7负调控下游AtBG1基因的表达影响植物体内ABA含量,从而响应干旱胁迫[26-27]。番茄SlTGA7与拟南芥中广泛参与SA调控的抗病反应基因AtTGA4、AtTGA7聚在同一分支[24, 28-29],猜测SlTGA7可能也具有相似的功能。根据上述结果推测SlJAZ7基因可能通过提高Pst DC3000病原菌侵染时SlTGA7基因的表达量,启动SlTGA7下游抗病基因的表达,从而提高SlJAZ7转基因番茄的抗病性。
MeJA比JA更稳定,但与JA作用相同,常被用来代替JA[30]。在对番茄JAZ基因的表达分析中,发现几乎所有SlJAZs的表达都受到JA和SA的调节,因此,Sun等[9]推测SlJAZs参与抗病的途径可能与JA和SA激素密切相关。本研究发现MeJA处理0.5 h时,SlJAZ7基因表达量显著上调,在1 h时达到最大值,说明SlJAZ7在JA诱导的抗病反应过程中属于早期基因,与番茄SlJAZ11基因的研究结果[30]一致。在SA处理过程中,SlJAZ7表达响应SA的模式与JA并不相同,大多处于相反状态,SlTGA7基因对2种激素的响应模式与SlJAZ7基因相反,这与JA与SA信号之间存在拮抗的报道[31-32]一致。JA与SA之间的协同作用表现在植物受到胁迫时,先激活JA介导的防御基因,随后激活SA信号介导的防御基因的表达[33]。已知JAZ基因家族在JA信号通路起作用,TGA基因家族在SA信号通路起作用,正是因为JA与SA信号之间存在串扰,因此,我们猜测SlJAZ7基因和TGA基因家族成员之间可能存在互作,共同调控番茄对Pst DC3000的抗病性。
本研究通过组织特异性表达分析发现SlJAZ7和SlTGA7在营养器官中都高表达,猜测两者都在营养生长中发挥功能,SlTGA7可能也参与果实发育。亚细胞定位显示SlJAZ7和SlTGA7蛋白均定位于细胞核,两者主要在细胞核中行使功能,这与SlJAZ7和SlTGA7基因家族的众多同源基因的研究结果类似,如SlJAZ11[30]、SlJAZ25[9]、GmJAZ3[34]、SlTGA2[28]、AtTGA7[27]均定位于细胞核中。JAZ蛋白作为JA反应的抑制子,已报道可通过与蛋白结合形成特异性的JAZ/TF复合物调控许多生理过程[35],通常涉及到丝裂原活化蛋白激酶(MAPK)、氧化还原调节谷胱甘肽(GRX)、MYC2、PDF1.2、WRKY70等[36]。植物体内JA含量高时,JAZ抑制因子被泛素化降解,释放转录因子,启动JA应答基因的表达。植物体内缺少JA时,JAZ蛋白抑制转录因子MYC2、MYC3、MYC4,从而抑制JA应答基因的表达[11]。本研究证明SlJAZ7蛋白可以与SlTGA7蛋白结合,猜测在SlJAZ7过表达的番茄株系中,SlJAZ7可能通过与SlTGA7蛋白结合,影响SlJAZ7与MYC2等转录因子形成复合物的功能,继而影响JA应答基因的表达。后期将细化转基因番茄的抗病试验,如通过在番茄中瞬时过表达及瞬时沉默SlTGA7后接种Pst DC3000,检测抗病相关基因如PR基因的表达量,验证SlTGA7的抗病功能,以及对SlJAZ7转基因植株接种Pst DC3000,利用定量PCR检测SlTGA7下游抗病相关基因的表达,检测SlJAZ7是否抑制SlTGA7的转录调控。
3.2 结论
为研究SlJAZ7的抗病机理,从番茄叶片中克隆得到SlJAZ7基因。利用农杆菌介导的遗传转化体系创制番茄过表达SlJAZ7转基因株系,接种Pst DC3000后,发现过表达SlJAZ7转基因株系积累了更少的活性氧,表明过表达SlJAZ7基因增强番茄对细菌性斑点病的抗性。表达模式分析发现SlJAZ7和SlTGA7都响应Pst DC3000、JA和SA处理,SlTGA7在SlJAZ7过表达的番茄株系中也高表达,说明SlJAZ7在转录水平正调控SlTGA7的表达。SlJAZ7和SlTGA7均定位于细胞核中,说明其主要在细胞核中行使功能。Y2H、BiFC和pull down试验证明SlJAZ7和SlTGA7蛋白存在互作,猜测SlJAZ7可能通过与SlTGA7互作,在蛋白水平参与番茄对细菌性斑点病的调控。
-
![]()
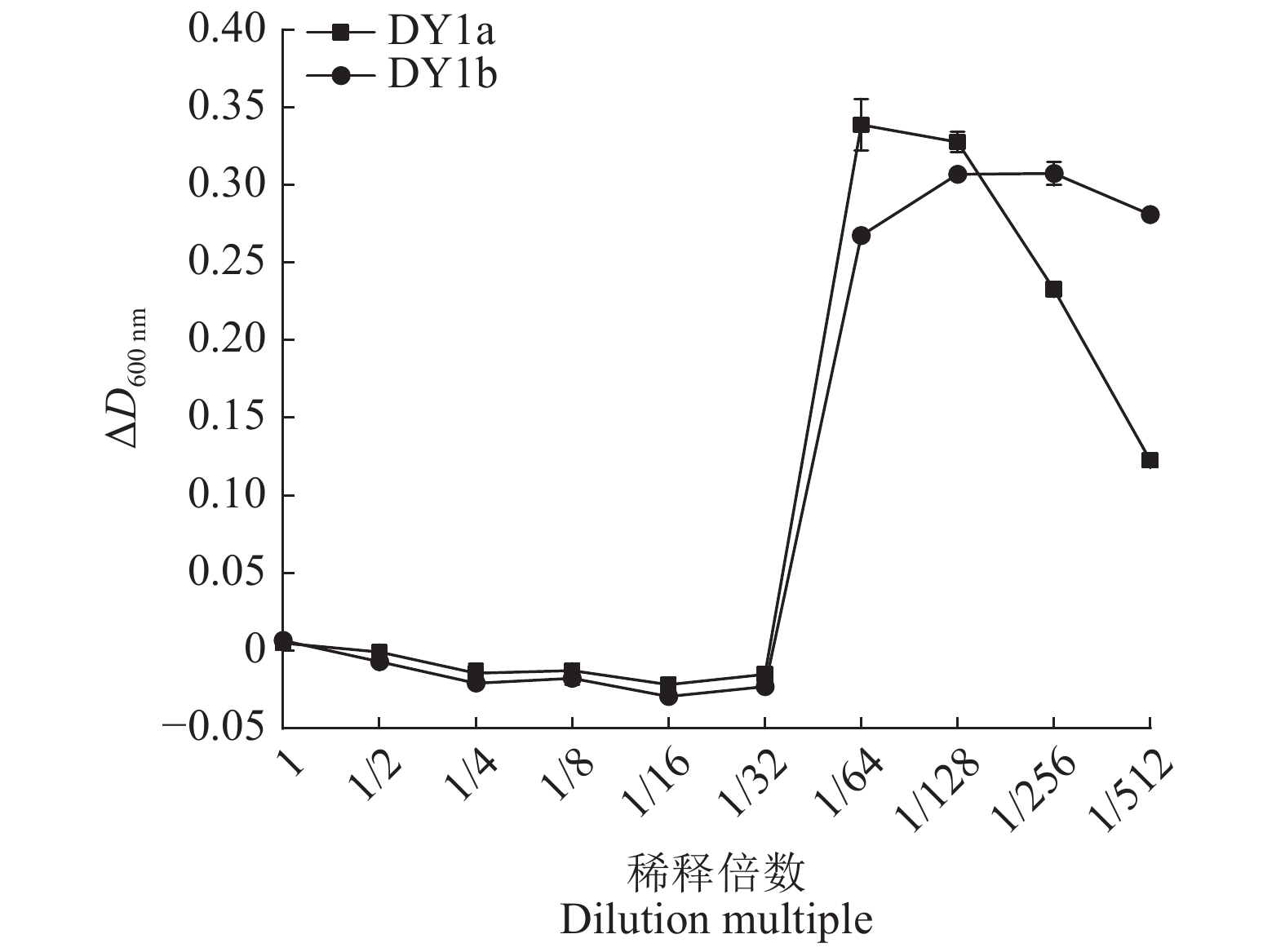
图 1 不同浓度大蒜有机硫化物(GOS)对2株解淀粉芽孢杆菌的抑菌作用
Figure 1. Antibacterial effects of different concentrations of garlic organic sulfide (GOS) on two strains of Bacillus amyloliquefaciens
![]()
图 2 不同浓度大蒜有机硫化物(GOS)对2株解淀粉芽孢杆菌生长的影响
GOS初始质量浓度为960 mg/mL
Figure 2. Effects of different concentrations of garlic organic sulfide (GOS) on the growth of two strains of Bacillus amyloliquefaciens
GOS initial concentration is 960 mg/mL
![]()
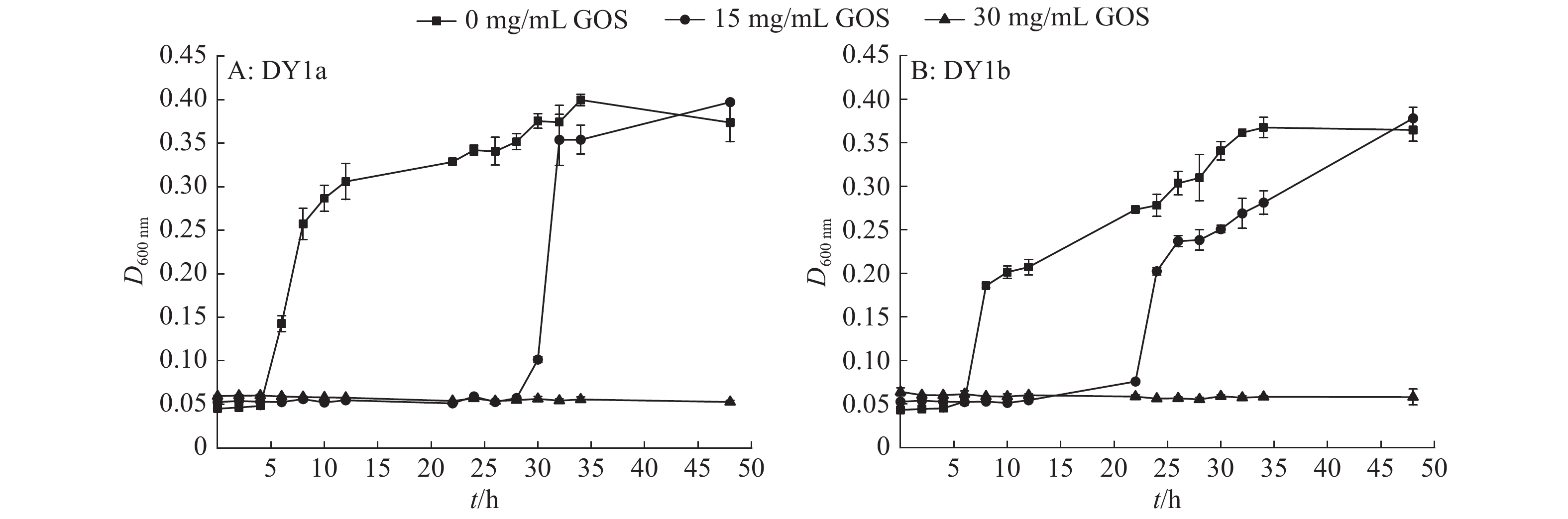
图 3 不同浓度大蒜有机硫化物(GOS)对2株解淀粉芽孢杆菌生长曲线的影响
Figure 3. Effects of different concentrations of garlic organic sulfide (GOS) on the growth curves of two strains of Bacillus amyloliquefaciens
![]()
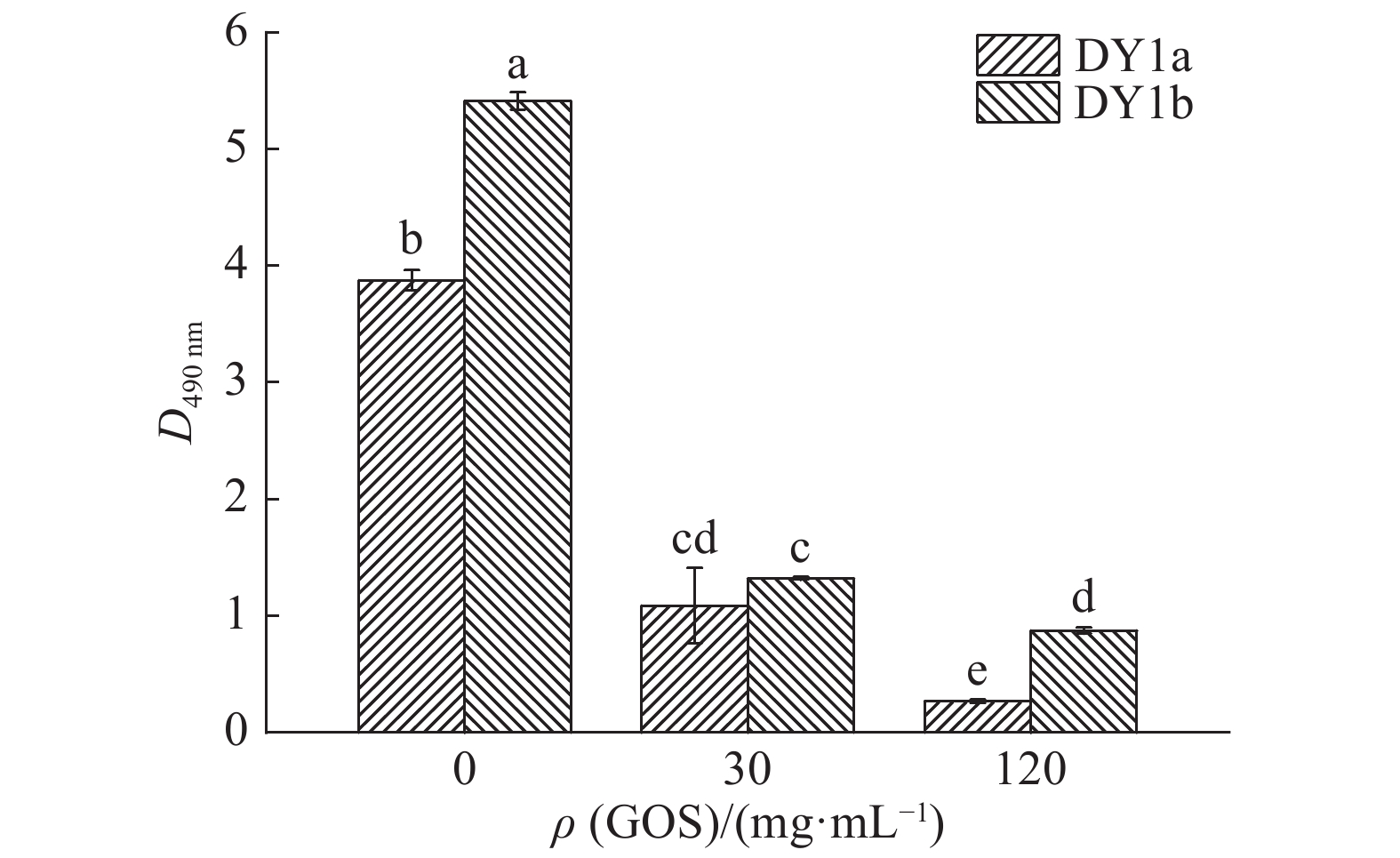
图 4 不同浓度大蒜有机硫化物(GOS)对2株解淀粉芽孢杆菌细胞代谢活性的影响
柱子上方的不同小写字母表示差异显著(P<0.05, Duncan’s法)
Figure 4. Effects of different concentrations of garlic organic sulfide (GOS) on metabolic activities of two strains of Bacillus amyloliquefaciens
Different lowercase letters above the column indicate significant difference (P<0.05, Duncan’s test)
![]()
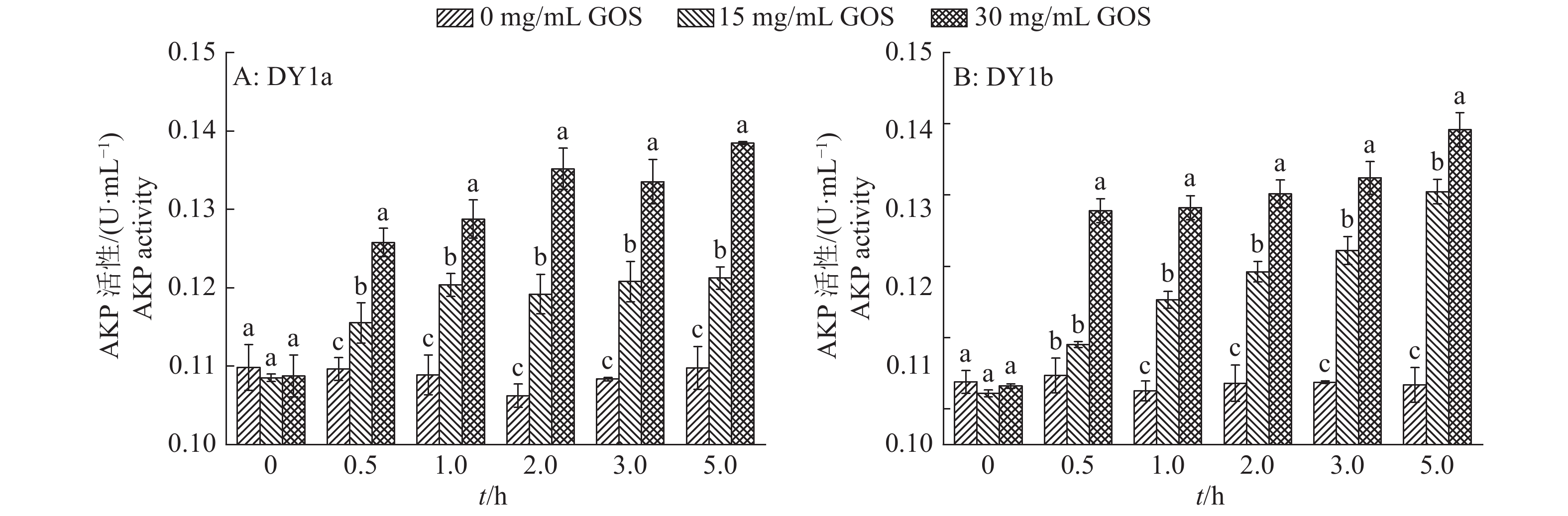
图 5 不同浓度大蒜有机硫化物(GOS)处理2株解淀粉芽孢杆菌的胞外碱性磷酸酶(AKP)活性的变化
各图中相同时间柱子上方的不同小写字母表示差异显著(P<0.05, Duncan’s法)
Figure 5. Changes of extracellular AKP activity in two strains of Bacillus amyloliquefaciens treated with different concentrations of garlic organic sulfide (GOS)
Different lowercase letters above columns of the same time in each graph indicate significant difference (P<0.05, Duncan’s test)
![]()
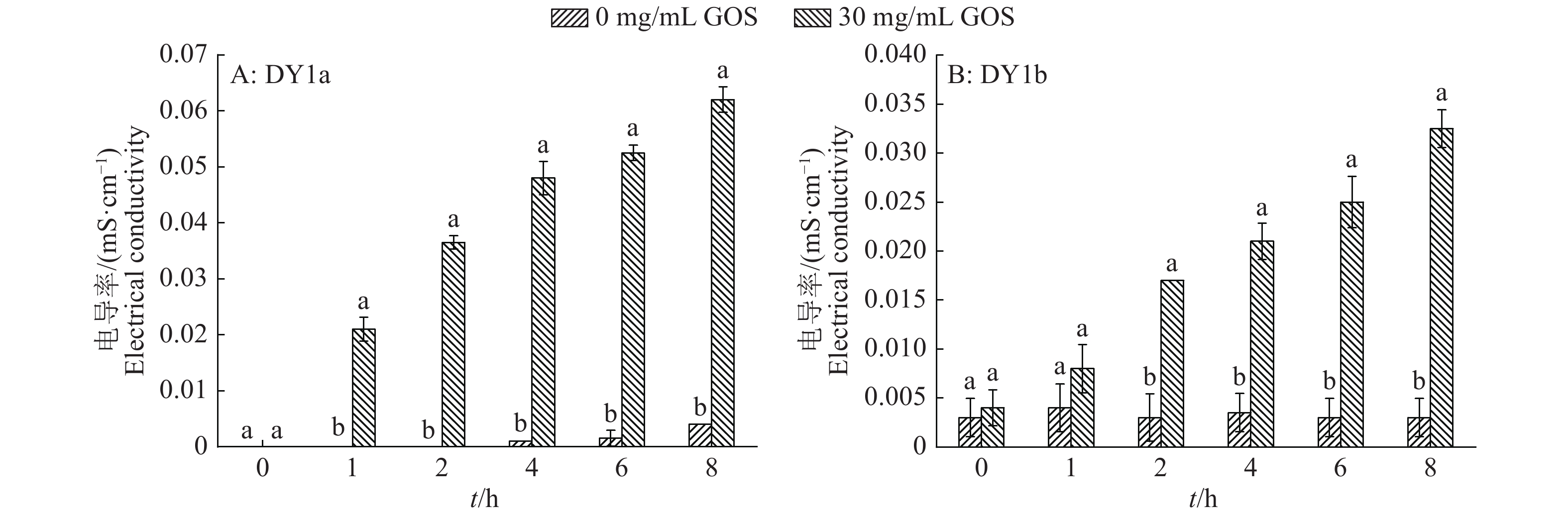
图 6 大蒜有机硫化物(GOS)处理2株解淀粉芽孢杆菌的胞外电导率变化
各图中相同时间柱子上方的不同小写字母表示差异显著(P<0.05, Duncan’s法)
Figure 6. Changes of extracellular electrical conductivity in two strains of Bacillus amyloliquefaciens treated with garlic organic sulfide (GOS)
Different lowercase letters above columns of the same time in each graph indicate significant difference (P<0.05, Duncan’s test)
![]()
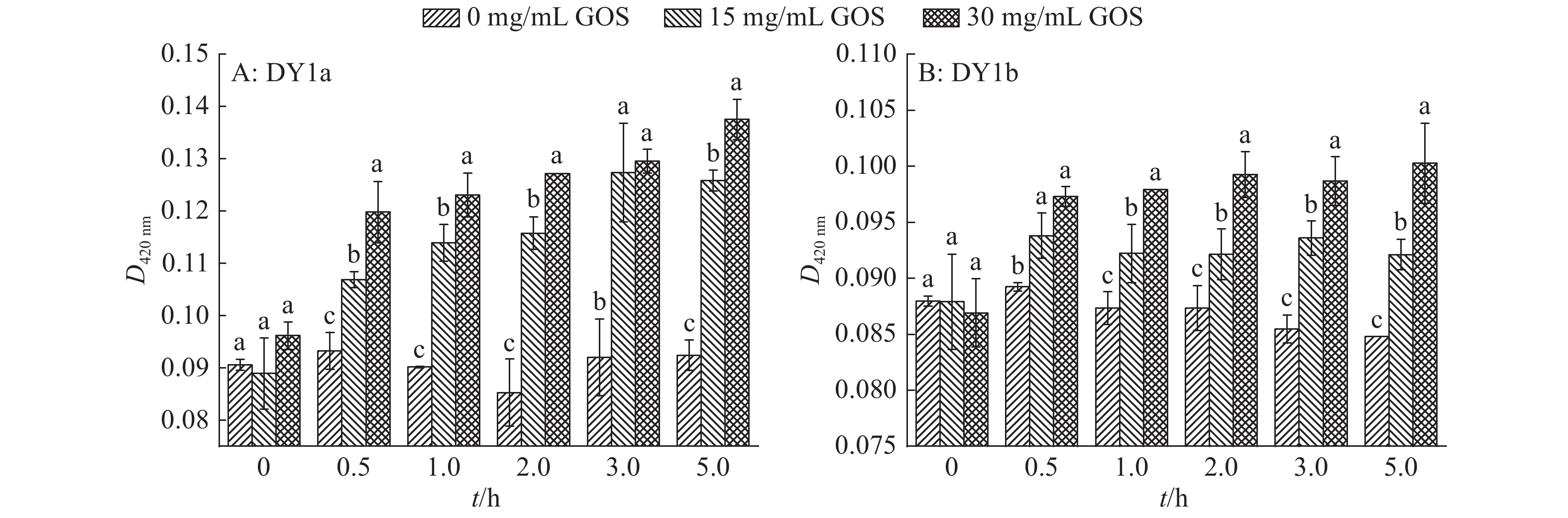
图 7 不同浓度大蒜有机硫化物(GOS)处理2株解淀粉芽孢杆菌的胞外β–半乳糖苷酶活性变化
各图中相同时间柱子上方的不同小写字母表示差异显著(P<0.05, Duncan’s法)
Figure 7. Changes of extracellular β-galactosidase activity in two strains of Bacillus amyloliquefaciens treated with different concentrations of garlic organic sulfide (GOS)
Different lowercase letters above columns of the same time in each graph indicate significant difference (P<0.05, Duncan’s test)
![]()
图 8 不同浓度大蒜有机硫化物(GOS)处理2株解淀粉芽孢杆菌细胞表面形态
A:未处理菌株DY1a;B:30 mg/mL GOS处理菌株DY1a;C:480 mg/mL GOS处理菌株DY1a;D:未处理菌株DY1b;E:30 mg/mL GOS处理菌株DY1b;F:480 mg/mL GOS处理菌株DY1b
Figure 8. Cell surface morphology of two strains of Bacillus amyloliquefaciens treated by different concentrations of garlic organic sulfide (GOS)
A: Strain DY1a untreated; B: Strain DY1a treated with GOS at 30 mg/mL; C: Strain DY1a treated with GOS at 480 mg/mL; D: Strain DY1b untreated; E: Strain DY1b treated with GOS at 30 mg/mL; F: Strain DY1b treated with GOS at 480 mg/mL
表 1 不同浓度大蒜有机硫化物(GOS)对2株解淀粉芽孢杆菌的杀菌作用
Table 1 Bactericidal effect of different concentrations of garlic organic sulfide (GOS) against two strains of Bacillus amyloliquefaciens
ρ(GOS)/
(mg·mL−1)菌落数/ (CFU·mL−1)
Colony countDY1a DY1b 480 0 0 240 0 50 120 20 290 60 300 >300  下载: 导出CSV
下载: 导出CSV
表 2 不同浓度大蒜有机硫化物(GOS)对2株解淀粉芽孢杆菌胞外蛋白酶活性的影响1)
Table 2 Effects of different concentrations of garlic organic sulfide (GOS) on extracellular protease activities of two
strains of Bacillus amyloliquefaciens ρ(GOS)/
(mg·mL−1)DY1a DY1b 水解圈直径/mm
Diameter of
hydrolysis zone蛋白酶活性抑制率/%
Inhibitory rate of
protease activity水解圈直径/mm
Diameter of
hydrolysis zone蛋白酶活性抑制率/%
Inhibitory rate of
protease activity0 17.41±0.85a 0 16.37±0.04a 0 15 16.17±1.36a 13.7 16.68±0.43a 0 30 12.64±0.71b 47.3 15.04±0.42a 15.6 45 7.00±0.00c 83.8 10.98±0.46b 55.0 1)同列数据后的不同小写字母表示差异显著(P<0.05,Duncan’s法)
1) Different lowercase letters in the same column indicate significant difference (P<0.05, Duncan’s test)
下载: 导出CSV
-
[1] 李永吉, 曾茂茂, 何志勇, 等. 腐竹加工技术及品质影响因素的研究进展[J]. 食品科学, 2013, 34(23): 333-337. [2] 郑丽君, 申光辉, 张志清, 等. 真空包装免泡豆杆优势腐败细菌分离鉴定及其致腐能力分析[J]. 食品科学, 2018, 39(2): 177-184. doi: 10.7506/spkx1002-6630-201802028 [3] 郑丽君, 申光辉, 张志清, 等. 免泡豆杆优势腐败菌腐败能力及产生物胺特性分析[J]. 食品与发酵工业, 2017, 43(10): 56-62. [4] 夏硕, 郑丽君, 申光辉, 等. 高温杀菌对预浸泡豆杆的杀菌效果及品质影响[J]. 基因组学与应用生物学, 2019, 38(5): 2086-2093. [5] 唐桂宇, 唐怡淋, 代博仁, 等. 免泡豆杆腐败菌复配抑菌剂的优化及其保鲜效果[J]. 食品与发酵工业, 2019, 45(4): 110-116. [6] 周建新. 植物源天然食品防腐剂的研究现状、存在问题及前景[J]. 食品科学, 2006, 27(1): 263-268. doi: 10.3321/j.issn:1002-6630.2006.01.063 [7] LEE Y S, LEE Y J, PARK S N. Synergistic antimicrobial effect of Lonicera japonica and Magnolia obovata extracts and potential as a plant-derived natural preservative[J]. J Microbiol Biotechnol, 2018, 28(11): 1814-1822. doi: 10.4014/jmb.1807.07042
[8] 陈宁. 蒜氨酸降解产物的成分分析和抑菌机理研究[D]. 天津: 天津科技大学, 2016: 1-7. [9] ANKRI S, MIRELMAN D. Antimicrobial properties of allicin from garlic[J]. Microbes Infect, 1999, 1(2): 125-129. doi: 10.1016/S1286-4579(99)80003-3
[10] XUE R, FENG J S, MA L N, et al. Whole transcriptome sequencing analysis of the synergistic antimicrobial effect of metal oxide nanoparticles and ajoene on Campylobacter jejuni[J]. Front Microbiol, 2018, 9: 2074. doi: 10.3389/fmicb.2018.02074
[11] REITER J, HUBBERS A M, ALBRECHT F, et al. Allicin, a natural antimicrobial defence substance from garlic, inhibits DNA gyrase activity in bacteria[J/OL]. Int J Med Microbiol, 2020, 310(1).[2019-10-11]. https://doi.org/10.1016/j.ijmm.2019. 151359.
[12] 冯少龙. 大蒜提取物中有机硫化物对阪崎克罗诺杆菌的抑菌活性与抑菌机理的研究[D]. 天津: 天津科技大学, 2014: 26-47. [13] 黄笠原, 毛顺, 李蕊婷, 等. 大蒜精油对熏马肠中德氏乳杆菌产腐胺的影响机制[J]. 食品科学, 2019, 40(5): 17-23. doi: 10.7506/spkx1002-6630-20180226-208 [14] 穆可云. 大蒜提取物对蜡样芽孢杆菌的抑制研究[D]. 广州: 华南理工大学, 2013: 36-47. [15] 彭光华, 韩月峰, 马荣池. 大蒜有机硫化物提取条件的优化及其成分的分析[J]. 食品科学, 2008, 29(7): 226-230. doi: 10.3321/j.issn:1002-6630.2008.07.047 [16] 陶文卿. 10-HDA对大肠杆菌抑制机理的研究[D]. 济南: 山东轻工业学院, 2012: 25-61. [17] GUO J J, GAO Z P, LI G Y, et al. Antimicrobial and antibiofilm efficacy and mechanism of essential oil from Citrus Changshan - Huyou Y. B. Chang against Listeria monocytogenes[J]. Food Control, 2019, 105: 256-264. doi: 10.1016/j.foodcont.2019.06.014
[18] 杨天佑, 田静, 黄新, 等. TTC–脱氢酶测定法表征低温真空环境对细菌呼吸活性的影响[J]. 食品与机械, 2008, 24(6): 32-35. [19] 刘梦茵, 刘芳, 周涛, 等. 乌梅提取物对蜡状芽孢杆菌的抑菌机理研究[J]. 食品科学, 2012, 33(1): 103-105. [20] 李婷, 杨舒然, 陈敏, 等. 姜厚朴水提物对大肠杆菌和金黄色葡萄球菌的抑菌机理研究[J]. 现代食品科技, 2016, 32(2): 84-92. [21] 牛贞福, 国淑梅, 徐金强, 等. 大蒜提取液对平菇竞争性杂菌的抑制及对平菇生长的促进作用[J]. 江苏农业学报, 2016, 32(1): 73-77. doi: 10.3969/j.issn.1000-4440.2016.01.011 [22] 张宝善, 王君, 陈锦屏, 等. 大蒜汁对枯草芽孢杆菌抑制作用的研究[J]. 西北植物学报, 2009, 29(6): 1269-1275. doi: 10.3321/j.issn:1000-4025.2009.06.032 [23] 宋卫国. 大蒜提取液有效成分抑菌活性及其作用机理研究[D]. 泰安: 山东农业大学, 2004: 51-61. [24] 胡蕾. 蒜氨酸降解产物结构鉴定及抑菌机理研究[D]. 天津: 天津科技大学, 2017: 28-51. [25] FENG S L, EUCKER T P, HOLLY M K, et al. Investigating the responses of Cronobacter sakazakii to garlic-drived organosulfur compounds: A systematic study of pathogenic-bacterium injury by use of high-throughput whole-transcriptome sequencing and confocal micro-Raman spectroscopy[J]. Appl Environ Microbiol, 2014, 80(3): 959-971. doi: 10.1128/AEM.03460-13
-
期刊类型引用(2)
1. 高鹏华, 齐颖, 杨敏, 李丽芳, 黄飞燕, 刘佳妮, 赵建荣, 余磊. 感染白绢病对魔芋内源激素含量及相关基因表达的影响. 华北农学报. 2025(03)  百度学术
百度学术
2. 李雪峰,火来勇. 番茄抗病品种选育及其在农业生产中的应用效果. 河北农业. 2025(04): 89-90 . 百度学术
其他类型引用(0)