
Research progress of grain shape genetics in rice
-
摘要:
粒形不仅是影响水稻产量的要素,还与稻米的外观品质密切相关,是决定稻米品质的重要因素。随着现代遗传学和基因组学相关理论和技术的发展,人们对粒形的遗传研究日趋深入,目前已有数十个控制水稻粒形的基因被克隆。本文概述了水稻粒形遗传研究主要进展,同时也指出了水稻粒形遗传研究中存在的主要问题,分析了粒形遗传研究的前景。
Abstract:Grain shape controls rice yield. Meanwhile, grain shape is related to rice appearance quality, and an important element determing rice quality. With the developments of modern genetics and genomics related theories and technologies, the genetic researches on grain shape are gradually deepened. Now dozens of QTLs controlling grain shape are cloned. This review summarizes the main progresses of genetic researches on grain shape, points out the major problems and analyzes the prospects.
-
Keywords:
- rice /
- grain shape /
- genetic /
- functional gene /
- molecular breeding
-
种业作为农业的“芯片”,是国家安全尤其是粮食安全的战略性产业。我国一直高度重视种业发展,从赋予育种者权利、推进种业商业化,到推动种业做大做强等方面,做出了一系列改革与部署[1]。2021年农业农村部公布的《种业振兴行动方案》强调,“农业现代化,种子是基础,必须把民族种业搞上去,把种源安全提升到关系国家安全的战略高度,集中力量破难题、补短板、强优势、控风险,实现种业科技自立自强、种源自主可控”[2];2022年的“中央一号文件”《关于做好2022年全面推进乡村振兴重点工作的意见》提出“全面实施种业振兴行动方案”;2023年发布的《粮食安全保障法(草案)》更是为保障粮食有效供给,提高防范和抵御粮食安全风险能力,确保国家粮食安全与维护经济社会稳定提供了法律支撑。
经过多年努力,我国种业发展取得了良好成效。据统计,2021年我国种子市场总规模约1 214.1亿元,农作物良种覆盖率在96%以上,自主选育品种面积占比超过95%。其中,水稻、小麦两大口粮作物的品种实现了完全自给,良种对粮食增产的贡献率超过了45%。水稻与玉米的杂交种子在国际种子贸易中也有突出的表现[3],并形成了海南、甘肃、四川三大国家级种业基地,总体上实现了种子供应的自我保障。2022年,我国粮食总产量达到68 653万t,分别是美国、印度的1.40和1.97倍;人均粮食占有量为486 kg;连续多年超过世界平均水平[1]。但需要注意的是,在我国种业不断发展壮大的同时,种业自主创新能力与发达国家仍有差距,表现为种子技术水平、市场规模和集中化程度较低,多元化竞争、国际市场竞争力不足,外来潜在风险突出[4]。重视粮食安全问题,在当下复杂国际形势和气候变化条件下,尤显其全局性和关键性。作为时代之需,当务之急,种业振兴将深远地塑造中国在地缘政治的博弈能力与国际社会的大国地位。
已有研究从世界粮食安全的现实局势到国家安全的政策引领、从种业产业的宏观结构到微观主体的创新研发行为、从种业技术水平的科学前沿价值到技术应用的多元价值实现等不同的层面,深度关注了种业振兴与粮食安全的逻辑关联性。本文试图从粮食安全决定因素的优先序视角进行全局性和趋势性研判,识别种业安全在其中的关键性地位;梳理中国种业发展历程及取得的成效;基于国家粮食安全的大食物观,剖析种业振兴现状与困境;并对未来中国种业振兴的健康良序发展进行展望。以期为推进中国种业振兴、赋能粮食安全提供科学的对策建议。
1. 粮食安全的决定因素:一个优先序
中国是农业大国,更是人口大国,面对气候尤其是国际地缘政治环境的巨大不确定性,中国农业现代化的核心目标应该是要提升安全保障力、资源控制力和国际竞争力,以谋求国家安全的“风险最小化”与“安全最大化”[2]。其中,粮食安全是最为根本的战略底线。夯实我国粮食安全的根基,必须识别粮食安全决定因素的优先序(图1)。
尽管中国粮食综合生产能力稳步提升,但一直受到包括耕地数量减少、质量退化、水资源短缺在内的资源环境“硬约束”,粮食持续增产难度加大。《中国统计年鉴》的数据表明,1949—2021年,中国粮食年总产量从1.13亿t增长至6.83亿t,年均增长率约为7.9%,同期人口总量从5.42亿增长至14.14亿,年均增长率为12.1%,粮食年均增长率比同期人口的年均增长率低4.2个百分点[5]。国内粮食消费需求呈刚性增长态势,预计到2030年,粮食供需缺口将达到3 000万t左右,缺口为4.6%~5.4%[6]。
从立足国内安全保障的层面来看,实现粮食充裕供给的路径:一是扩大种植面积,二是提高农业生产率。但面临的现状却是,一方面,在耕地资源的刚性约束下,我国粮食种植面积不可能大幅度扩张。第3次全国国土调查数据显示,我国耕地面积为1.279亿hm2,比第2次调查减少0.075亿 hm2;在耕地减少的同时,人口规模却从2000年的12.67亿增至2021年的14.13亿。人均耕地面积分别由前2次国土调查的0.106和0.101 hm2,降至第3次调查的0.091 hm2,已不足世界平均水平的40%[2]。中国现有农户数为2.3亿,户均耕地面积仅为0.520 hm2,仅相当于美国户均规模的1/400、欧盟户均规模的1/40,也不足日本户均规模的1/4[5]。另一方面,先进科技和适用的装备技术在农业中的推广应用并不充分,制约了农业生产效率的提升。尤为突出的是,“大国小农”的分散化小规模经营格局严重削弱了农产品生产的潜在供给能力。第3次农业普查数据显示,我国小农户的数量占农业经营主体98%以上。经营土地规模在0.667 hm2以下的农户占家庭承包户总数的比例从1996年的76.0%增至2018年的85.2%,2021年依然维持在82.8%的水平[2]。土地的均田承包与农业的分户经营不仅造成户均耕地面积小,而且造成地块细碎分散,约束了农业生产效率的提高。与此同时,农村劳动力非农转移不仅诱发了农业优质人力资本的流失,也进一步加剧了农业的边缘化与弱质化。必须清醒地认识到,在未来相当长的时期,中国“大国小农”和“人多地少”的基本国情和农情很难发生根本性改变[7-9]。因此,对我国的粮食安全问题不能有任何懈怠。
保障粮食及重要农产品的供给安全需要做出2个方面的努力:第一,要树立大资源观与大国土观,从耕地资源延伸到整个国土资源,宜粮则粮、宜经则经、宜牧则牧、宜渔则渔、宜林则林,广泛开发食物资源,不断拓展粮食安全空间。第二,要着力提高单产水平,在落实和拓展“藏粮于地”战略的基础上,把“藏粮于技”摆在更为重要的战略高度。这2个方面的努力都依赖于种业创新能力的提升,既涉及与大资源观相匹配的种源创新多样化,也包括与高产优质相关联的良种繁育精准化。
种业振兴的关键是良种繁育,而良种繁育的根基是种质资源保护和有序利用。中国是世界上主要农作物起源的中心之一,种质资源的丰富性和独特性为现代种业发展提供了基础。实践表明,种质资源分别对粮食增产、畜牧业发展的贡献率超过45%和40%[10]。良种繁育的前提是技术进步及创新。周华强等[11]通过构建育种科技创新项目绩效评价模型的分析发现,育种科技创新项目支撑了现代农业发展与粮食安全。周鸣川等[12]研究表明,精准、定性和定向的育种技术提高了作物在面对恶劣环境条件(如干旱、高温、病虫害等)时的适应能力,促进了作物生长和产量提高。尹春凤等[13]基于我国2001—2020年31个省(区、市)数据的分析发现,种业创新与粮食安全的耦合协调度、复合系统协同度均呈现明显的强化趋势。
种业振兴依赖于良种繁育的精准指向性。一是粮食作物类的种业。联合国粮农组织的研究显示,世界粮食总产量的增长80%依赖于粮食作物类的良种繁育与推广[14]。科学试验也表明,良种在提高粮食作物单产水平中发挥着关键性作用,所有增产技术及高产指标的实现很大程度上依赖于良种的内在潜力。良种在发达国家粮食增产中的贡献率为50%~60%[15]。在我国,粮食单产水平从新中国成立时的1029.33 kg/hm2提升至2020年5734 kg/hm2的过程中,良种的贡献率高达到45%[16]。二是食物营养类的种业。粮食仅仅是食物的一部分。《中国统计年鉴》的数据显示,2013—2021年,我国居民人均粮食消费量下降了2.76%,人均肉类、禽类、水产品、蛋类消费量分别增加了28.52%、70.83%、36.54%、60.98%[17]。这一消费趋势的转型为中国粮食安全体系建设提出了更为丰富的内涵,即从“吃饱”(产量)向“吃好”(结构、营养与品质)转变,从而使得良种繁育从过去的“增产导向”进一步向“提质导向”的结构性转换。显然,要实现这一历史性转型依赖于更为丰富多样的食物营养类的种业创新发展。三是具有生态包容性的种业。长期以来,单极化地提升农业生产效率的努力已造成相对严重的农业面源污染,表现为水体富营养化、土壤重金属污染加剧以及温室气体排放增加。2019年我国农用化肥施用折纯量仍然高达5403.59万t、农药使用量达139.17万t、农用塑料薄膜使用量达240.77万t,远高于世界平均使用强度[2, 18]。种业创新并非独立事件,需要良种与良田、良机、良法、良制等多个维度的相互依存与合理匹配,具有生态包容性的种业创新应该有助于维护耕地资源的可持续性,既能够节约用地又能够促进土地的永续利用;有助于维护水资源的可持续性,既能够节水又能够促进水环境生态的改善;有助于维护农业的生物多样性,既能够保障粮食安全又能够提升农业的生态功能并凸显中国农耕文明所秉承的生态智慧及时代价值。
2. 种业的发展历程及面临的问题
种业振兴是涉及面十分广泛的议题。从实施内容看,种业振兴表达为种子研发与技术扩散,前者是通过种源保护及关键核心技术攻关,打通种业“最先一公里”;后者是通过技术应用及市场机制运行,打通种业“最后一公里”。从实施路径来看,《种业振兴行动方案》提出了“加强种质资源普查和保护”“推进种业创新攻关和实施实物育种重大项目”“扶持优势种业企业发展”“提升种业基地建设水平”“严厉打击套牌侵权等违法行为”等一系列的关键性措施与行动重点。回顾种业的发展历程,发现所面临的问题,有助于厘清我国种业振兴的方向性策略。
2.1 种业的发展历程
种质资源保护和种业技术创新具有消费上的非排他性和收益上的非独占性,即种业具有公共物品性质。因此,种业振兴不仅要提高产业主体的参与程度,也要发挥政府的主导性作用。新中国成立以来,在党和政府的领导下,我国种业经历了育种群众化、育种专业化、种业产业化、种业现代化和种业振兴5个阶段。
2.1.1 第1个阶段(1949—1977年):育种群众化
标志性事件为1950年原农业部颁布的第1份种业规划《五年良种普及计划草案》,并架构了“家家种田、户户留种”为特征的群众化育种体系。经过群众选育、引进吸收和科研培育,到1957年我国共选育新品种555个[19]。1958年全国种子会议进一步提出了“主要依靠农业社自繁、自选、自留、自用,辅之以必要调剂”的“四自一辅”工作方针,规定种子“集中管理、统一调种”[19]。这套规则延续了较长时期, 1972年在国务院批转的原农林部《关于当前种子工作的报告》中确立了主导地位,形成了县、公社、生产队3级良种推广体系。事实上,这一推广体系已经成为我国种业创新的重要制度遗产。
2.1.2 第2个阶段(1978—1994年):育种专业化
标志性事件为1978年原农林部在《关于加强种子工作的报告》中提出的“四化一供”工作方针,并成立了中国种子公司。种业从传统的农户留种繁育逐渐向专业化转变,种业方针也转向“生产专业化、加工机械化、质量标准化、品种布局区域化”[3]。1988年,中国种子公司正式加入国际种子贸易协会,并开展种子进出口工作。1989年出台的《中华人民共和国种子管理条例》对种质资源管理、选育与审定、生产经营、检验与检疫、贮存等作出了相关规定,初步形成了“国有农业科研院所负责良种繁育、国有种子公司安排生产,乡镇农技推广部门负责良种推广和销售”的种业分工体系与上、下游格局,但同时也割裂了良种育种与研发、生产加工与销售之间的直接联系。
2.1.3 第3个阶段(1995—2010年):种业产业化
标志性事件为1995年全国农业种子工作会议提出的“创建种子工程”,为面向市场的种业产业化体系建设提供了支持。期间,全国商品种子繁育生产能力由50亿kg增加到80亿kg,加工能力由30亿kg增加到50亿kg,良种覆盖率达到95%以上[20]。在种业经营主体方面,育、繁、推一体化企业主体不断增多,至2010年发展到近100家。其中,前50强种业企业经营额由30亿元增加到100多亿元,市场占有率由10%提高到30%以上[19];在种业基础设施方面,累计投资近63亿元,建成农作物种质资源库(圃、区) 39个、品种改良(分)中心96个、品种区试站135个、国家级原种场54个、种子种苗繁育基地304个、国家救灾备荒种子储备库34座,以及国家、省、市、县4级种子检测中心92个,引进购置种子加工成套设备815条[19, 21];在种业立法方面也迈出了重要步伐,1997年颁布的《植物新品种保护条例》承认了育种者的权利;2000年颁布的《中华人民共和国种子法》进一步突破了种子产业的计划经济体制及国有种子公司的垄断经营。1996—2000年,国家审定的主要农作物品种达1838个,平均每年通过审定的新品种数量约为131个,为1990—1995年期间的2.7倍。2010年的“中央一号文件”提出“切实把农业科技的重点放在良种培育上,加快农业生物育种创新和推广应用体系建设”,商品种子供应率由20世纪90年代中期的30%提高到2010年的60%,良种对粮食增产的贡献率从2003年的36%提高到2009年的40%[20-21]。
2.1.4 第4个阶段(2011—2020年):种业现代化
标志性事件为2011年国务院颁布的《关于加快推进现代农作物种业发展的意见》,明确了种业的战略地位,提出了建构“以产业为主导、企业为主体、基地为依托、产学研相结合、育繁推一体化”的现代农作物种业体系[19]。2014年以后,我国专利申请数量超过国外专利申请数总和,但品种同质化、模仿育种、套牌侵权等问题仍然突出。据统计,在农业农村部保藏中心保存的1800份水稻品种中,有50%的品种与其近似品种的遗传差异低于10%,农业专利以简单修饰或选择变异株品种为主[22]。为此,2015年修订的《中华人民共和国种子法》新增了“新品种保护”。截至2019年,品种权申请数量为7 032件,大田作物申请数量接近5 000件,占比为70.58%。1999—2019年期间,国内植物新品种权累计申请数量年均增幅达28.80%[23]。2020年颁布的《中华人民共和国民法典》进一步将“植物新品种权”列为知识产权。
2.1.5 第5个阶段(2021年至今):种业振兴
标志性事件为2021年中央全面深化改革委员会审议通过《种业振兴行动方案》、2021年中央经济工作会议和中央农村工作会议提出要“深入实施种业振兴行动”。得益于政策努力,国家种质库保存的种质份数达到441 388份,作物种质资源长期保存的份数达到517 299份,保存总量稳居世界第2 [24]。不过,由于我国种子贸易国际地位较低,知识产权保护力度较弱,导致国外授权量全球占比不足1%。为此,2021年修订《中华人民共和国种子法》将惩罚性赔偿上限由3倍提高至5倍,并建立了实质性派生品种制度。截至2021年底,假劣种子案件比新《种子法》实施前减少了48%、侵权案件减少了36%,2012—2021年种子出口批次增长了约141.6倍[25]。
2.2 种业发展面临的问题
2.2.1 种业发展的法律保护和国际规则主导权不足
一方面,立规立法、司法保护、行政执法、技术标准、社会共治是构建有序的种业市场的关键,是增强种业科技创新能力、确保粮食安全的有效措施。从这个角度看,种质资源保护体系应由国家牵线、省级部门落实管理。但存在众多短板,包括在立法层面,种业知识产权保护法规体系还在逐步完善;在司法层面,行政与司法保护衔接机制没有形成;在执法层面,品种标准样品管理、分子检测技术研发应用水平低,打假护权技术难以提供执法支持。此外,侵权者逃避侵权赔偿责任的方式也更为多样,除“顶包” “白皮袋种子”外,还通过虚假的资本转让或利用注册商标等抗辩不侵权。上述种种因素,削弱了维权者的积极性。在新品种保护方面,套牌假冒、“三无”种子等侵权行为的出现,以及普遍存在的机会主义行为,导致“劣币驱逐良币”,进而抑制了大企业新品种创新的积极性。据统计,2021年4月至2022年1月,全国查办种业违法案件达6 000多起[26]。
另一方面,发达国家主导国际农业贸易投资规则,使得我国种业的国际化发展受阻。长期以来,在世界经济治理体系中,由于GATT/WTO规则引领全球贸易,发达国家在其中扮演着重要角色。中国作为农业大国,更容易受到国际社会高度关注,遭遇国际规则的歧视性阻挠和西方国家掌握的舆论工具的欺骗性报道,往往处于不利地位。近年来中国农作物种子的贸易逆差越来越大。虽然在“一带一路”倡议的指引下,我国先后出台了一系列鼓励加强农业科技合作的政策,但对外合作平台和运行机制难以适应国际种业发展的竞争模式,与我国强大的种业科技实力不相匹配。种质资源出口管理、对外合作平台、对外合作协调机制、目标国政策协调等方面亟待完善,为我国种业科技对外合作的主体提供有效支撑。
2.2.2 种业的国际市场竞争力有限
随着全球化进程与种子市场开放,跨国种子企业不断涌入中国市场,挤压中国种子企业的生存空间。有资料表明,跨国种子企业已经在中国实施了“产−研−发”部署,对中国种质资源及种子安全形成了威胁。根据邵长勇等[27]调研,在中国登记注册的外商投资农作物种业企业有76家,其中,26家为独资公司、42家为合资公司、8家为中外合作经营公司。这些名义上只占合资公司49%股份的外商却掌握了技术和专利等核心资源,随着跨国种子企业的主导作用逐渐增强,种业振兴赋能粮食安全的难度加大。
有证据表明,跨国种子企业利用技术优势对东道国产生了明显的农业垄断势力,削弱了发展中国家的粮食自主权[28]。马述忠等[29]构建了一个以本土和外国(商)企业为主体的仿真系统,模拟外企对中国种业的影响,结果表明跨国种业集团凭借资本和技术优势,形成了寡头垄断或垄断市场,挤占中国种子企业的市场份额。当外资撤出,中国种子企业研发产出和销售利润在短期内将提升,增幅却持续下滑。靖飞等[30]研究表明,中国种业上市公司与跨国种子企业在经营规模、经营绩效等方面存在明显差距;在种子业务毛利润率方面,跨国种子企业远高于中国种业上市公司,其中,孟山都高达60%以上,中国种业上市公司除登海种业外,其他公司都低于40%。
2.2.3 种业创新研发能力不足
种业竞争的关键是种子产品,而种子产品依赖于企业的研发行为。表1从研发模式、育种技术、企业发展和种质资源4个方面对国内外种业发展情况进行了比较。我国农业科技投入只占农业GDP 的0.71%,远低于发达国家2%~3%的投入水平;我国农业科技投入的80%以上主要用于农业科技应用技术,育种研发投入严重不足[26]。2019年拜耳作物科学事业部年度研发投入超过23亿欧元,其中,种子与性状研发方面的投入超过12亿欧元;科迪华农业科技公司的研发投入为11.47亿美元[31]。强大的资本整合优势不断加剧了种业企业的寡头化与垄断化。相反,2019年中国规模种业企业(注册资本≥3 000万元)研发投入累计才36.05亿元,其中,10家A股上市公司研发支出仅为6.69亿元[31]。尽管我国形成了以科研院所、高校与企业的“政府+市场”模式,但由于种质资源和育种人才集中于科研机构和高校,使得种子企业的技术创新能力及人力资本支持能力严重不足。同时,科研院所和种业企业缺乏有效的联结机制,不仅造成育种和经营等不同环节的脱节,也降低了良种推广的效率,导致企业往往通过直接购买种子经营权以获得种子销售资质,进一步抑制了企业参与育种研发的积极性,使得企业难以成为育种研发的核心主体。
表 1 种业发展的国内外比较1)Table 1. Comparison of seed industry development at home and abroad项目
Item一般趋势
General trend发达国家
Developed country中国
China研发模式
R&D mode种业研发模式的演变过程主要体现在研发主体的改变 经历了以政府(科研院所)研发为主、以政企联合研发为主和以大型跨国企业研发为主3个阶段,当前已经由政府为主过渡到以企业为主 经历了“自己留种−国家调配−市场供种”的发展过程,与之对应的种业研发主体则由以科研院所为主向以企业为主过渡 育种技术
Breeding technology4个阶段:以农民经验留种的1.0,以统计和遗传性质试验育种的2.0,以分子标记和生物技术育种的3.0,以生物技术和大数据为主的智能育种4.0 已经进入育种4.0 阶段,2019年美国转基因作物播种面积为
7 150万hm2,占全球转基因作物播种面积的37.6%处于2.0 向3.0过渡期,2019 年中国转基因棉花和木瓜播种面积为320万hm2,占全球转基因播种面积的1.7% 企业发展
Enterprise development3次并购潮:种子与农药的纵向并购(1996—2000年),种业之间的横向并购(2004—2008年),农化巨头的资本整合(2015—2018年) 德国拜耳与美国科迪华成为目前全球种子企业的第1梯队 中国先正达属于第2梯队,中国种子企业竞争力指数排名位居世界第3 种质资源
Germplasm resource世界各国争夺的重要战略资源,国际种子联盟(ISF)统计,截至2020年,全球共有740万份种质资源、1750个基因库 美国是全球种质资源保存数量和种类最多的国家,其中,国外种质资源数量占比高达72% 中国保存的种质资源总量超过52 万份,位居世界第2,国外种质资源占比仅为24% 1)资料来源:根据文献[32]进行整理
1) Source of data : Organized according to literature [32]另外,发达国家及种子企业长期在全世界收集种质资源。据国际种子联盟(ISF)统计,截至2020年底,全球共有740万份种质资源、1 750个基因库。美国早在第一次世界大战时就在全球搜集不同生态条件下的种质资源,现在已成为全球种质资源保存数量和种类最多的国家,其中,国外种质资源数量占比高达72%[32]。仅仅是科迪华公司,就在世界126 个试验站之间交流和共享种质资源,拥有全球60% 以上极具遗传潜力的玉米种质资源[32]。相反,我国早期对种质资源保护意识薄弱、保护措施不力,导致部分种质资源流失海外。目前,我国持证种子经营企业8 000余家,70%为小企业[33]。小企业获取、整合、利用资源的能力较弱,应对技术创新风险的能力较差,难以保证创新资源得以合理使用。李万君等[34]基于210家玉米种子企业的调研数据显示,规模化与种子企业技术创新产出数量成正比,规模较小的种子企业不具有持续创新能力。此外,由于种子在田间生长,创新质量难于观察,且产出与气候等自然因素相关,创新绩效往往具有不可考核性,这就使得品种权的保护取证、侵权界定和处罚相对困难,始终面临着套牌剽窃等侵权行为的长期挑战。加之种业的知识产权保护规则不完善、保护标准及范围偏低,使得模仿创新成为常态,从而对原始创新形成挤出效应[31, 35]。种子企业(尤其是小企业)不愿意投入时间和精力开发原创性品种,也不愿意采纳数字化技术扩展农业领域的应用,进一步诱发了企业的机会主义行为,往往以低成本扩张的方式通过边际改良开发新品种以快速占领市场,导致创新动力不足。
3. 大食物观视角下的种业振兴
3.1 大食物观
中国的粮食安全正经历由数量安全向品质安全和结构安全的双重转变,居民的食物消费模式进一步转向多元化、营养化、健康化。在2019年10月14日国务院新闻办公室发布的《中国的粮食安全》白皮书中,正式采用了“‘确保谷物基本自给、口粮绝对安全’的新粮食安全观”这一提法,从而确立了“以我为主、立足国内、确保产能、适度进口、科技支撑”的国家粮食安全战略[36]。提升粮食生产能力、重视粮食品质、重视“食物”概念、扩大“粮食”口径范围、充分利用国际市场、保护生态多样性、参与全球粮食安全治理等理念,均已被视为广义粮食安全观的重要内容。随着居民生活水平的提高,有必要将植物性食物的粮食概念转换成包含植物性食物、动物性食物甚至微生物食物的新概念,并将“粮食安全观”扩展为“食物安全观”的呼吁也愈发强烈[7, 37]。
习近平总书记指出,“要树立大食物观,从更好满足人民美好生活需要出发,掌握人民群众食物结构变化趋势,在确保粮食供给的同时,保障肉类、蔬菜、水果、水产品等各类食物有效供给,缺了哪样也不行”[38]。2023年的“中央一号文件”提出,“树立大食物观,加快构建粮经饲统筹、农林牧渔结合、植物动物微生物并举的多元化食物供给体系。”因此,端好14亿多中国人民的饭碗,不能仅在有限的耕地资源上不断索取。与传统粮食安全观相比,大食物观更具有引领性,通过全方位开发耕地、森林、海洋资源,拓宽动物、植物、微生物的食物来源,促进各类食物与营养的供求平衡,更好地满足人民群众日益多元化与高端化的消费需求[18];大食物观更具有全局性,通过拓宽粮食安全边界,既保障主粮生产对粮食安全的根基作用,又注重食物与营养的结构性拓展,有助于构建更为完整的国家食物安全体系。
3.2 种业转型升级
践行“大食物观”,全方位多途径开发食物资源必须在种子资源保护、种业科技创新、新品种培育、知识产权保护等方面做好文章,实现种业科技自立自强、种源自主可控,用中国种子保障中国粮食安全。大食物观势必倒逼种业的转型与提质升级,以下2个方面的转型升级尤为重要。
3.2.1 种业发展的品种均衡问题
目前,我国稻米和小麦等口粮可以实现自给,玉米从“供求宽裕”转向“产不足需”,而大豆供求缺口以及多种农产品供需失衡局面则更为突出。预计到2035年,我国许多食物需求量将高于国内生产量,产需缺口进一步拉大,如玉米自给率将下降到82%,牛羊肉和奶制品的自给率将降低为70% ~ 80%,食糖自给率将缩至40%,大豆供需缺口也将进一步扩大[39]。因此,种业振兴不仅要保证现有口粮的供给,更要重视玉米和大豆等粮食品种的研发,并提升牛羊等畜产品种源研发水平与品种效益。总体来说,促进种业振兴的品种均衡,必须打破“重粮轻经” “重平原轻山区” “重陆域轻海域” “重动植物轻微生物” “重数量轻营养” “重温饱轻健康”的偏差格局。
3.2.2 种业振兴的多目标兼容问题
种业振兴既要保障农产品供给的数量和质量,也要有助于支持食物生产的低碳化、绿色化与可持续发展。我国地理气候类型复杂多样,种业的发展应该支持农业结构与区域布局的调整优化,并促进从耕地资源向整个国土资源的拓展,向森林要食物,向江河湖海要食物,充分发挥不同地区自然资源的比较优势[18]。
传统食物的供给结构关注于2个食品供给生产部门:一是基于植物驯化发展起来的种植业,主要是提供粮食和果蔬等;二是基于动物驯化发展起来的养殖业,主要是提供肉、蛋、奶等。在此过程中,微生物仅发挥了支撑种植业和畜牧业发展的工具性作用[18]。与之相对应,传统的种业也主要关注于作物与动物的种质资源开发与技术创新。随着现代科技的不断进步与食物营养概念的不断拓展,微生物在作为种植业和养殖业工具性手段的同时,越发成为重要的食物来源或食物营养的替代品。中国已基本具备将微生物升级为现代食物供给部门的现实条件,因此,必须高度重视微生物领域的种业创新。此外,鉴于中国的粮食压力主要来源于膳食结构升级引发的“人畜争粮”,通过微生物饲料替代传统饲料,将显著增强中国粮食安全的兜底韧性与回旋空间。
4. 种业振兴的关键领域及展望
4.1 动态发展格局下解决种业的“卡脖子”问题
深化种业创新体系的体制机制改革,建立适应现代生物育种技术发展的种业科技创新体系,并重视作物的遗传特性、生理机制和适应性等基础研究,是推动种业振兴的根本举措。必须从以下2个方面解决“卡脖子”问题。
一是技术方面的问题。通过传统育种方法和现代生物技术相结合的策略,运用杂交与基因编辑等现代生物技术手段,加快品种改良和培育过程[40]。
二是体制机制方面的问题。重点在于:1)建立多方协同体系以及常态化政策扶持机制,以提高产业主体繁育良种的主动性[41];2)妥善保存种质遗传资源是关键技术攻关的基础,这需要建立种质资源的共享平台,加强稀有种质资源的收集与保存,重视种质资源的评价与鉴定,为培育适应性强的新品种提供遗传基础[42];3)建构“政府+企业+农户”模式,通过政府提供要素激励,企业将现代要素融入生产领域,农户(尤其是新型农业经营主体)负责留种和保护种质,实现“利益共享、风险共担”的种业发展机制;4)种业高质量发展在于改革以科研机构为主导的作物育种研发机制,这不仅需要进一步调整科研院所的研究定位,更需要规范种子研究课题的申报与考核,推动科研院所与其所办种子企业的“事企脱钩”;5)加强基地建设,建成一批现代化种养业良种生产基地,形成保(种质资源保护)、育(育种创新)、测(品种测试)、繁(良种繁育)分工合作和紧密衔接的现代种业发展格局;6)完善以企业为主体的商业化的育种研发推广体系,加大对“育−繁−推”一体化的种子企业研发资金扶持力度,实现优势资源重组[43]。其中,推广“公司+农户”基础上的种子产业的生产托管与服务外包等社会化组织方式,是将小农户融入现代种业发展体系的重要途径。
4.2 协同发展思路下重视种业振兴的匹配条件
种业振兴的实现在突破种业技术瓶颈的同时,还有赖于土地要素匹配、农资投入品绿色化、农业全产业链的数字化转型等方面的协同发展。首先,推进种业振兴不仅要保障耕地的数量,更要提升耕地的质量。农产品产量高低和品质好坏与良种以及耕地地力、土壤健康、灌溉条件、装备设施乃至于气候条件等多个因素紧密关联,农产品的持续产出及质量安全是一系列因素的函数。因此,种业振兴不仅依赖于技术创新,而且依赖于农地制度、耕地保护、公共政策等体制机制的创新。其次,种业振兴的增效提质有赖于生产的中间过程的绿色化转型。农业生产过程中的水、肥、药等中间投入品均会与种子共同发挥作用,影响农产品的生产过程以及最终产品的品质。值得注意的是,不同类型农户投入品的减量具有差异性,并与其要素禀赋特性和要素匹配度紧密关联,生存型农户偏好以家庭生态圈内部要素替代外部要素来弱化毒性成分以保护土壤;生活型农户偏好以节省劳动力的施肥措施改进来降低肥料损耗;生产型农户偏好以改善化肥养分元素配比来增加作物吸收率;功能型农户偏好通过危害度更低的外部要素替代来弱化毒性成分。而且新型经营主体和服务供应商偏好以施用工具的改进来降低化肥损耗[44]。因此,种业振兴不仅是技术要素的匹配问题,更要考虑农业经营主体尤其是技术采纳主体的适配性问题。最后,种业创新的赋能增效有赖于农业全产业链的数字化转型。农业产业的数字化有利于数据共享流动、优化生产决策效能;互联网信息技术数字红利的正向作用有助于缩小群体间和地区间的数字鸿沟、发挥小农户比较优势并调动其积极性,促进包容性的现代农业发展[45-46]。
4.3 国际视野下拓展种业振兴的发展空间
必须考虑到中国种业企业竞争力和影响力的短板问题,并强化种业知识产权保护机制,积极与国际接轨[47]。在研发阶段,应持续扩大植物新品种权保护范围和保护环节,建立实质性派生品种制度,激励育种原始创新,着力降低种业研发及知识产权获取的交易成本,如缩短品种权的审批时间,提高品种创新积极性,尽快制定适应国际知识产权保护的政策体系;在生产与经营阶段,推进第三方机构对植物品种权的价值评估,健全种子企业并购重组定价标准和交易程序,提高种子企业产权转让效率和成功率;完善侵权赔偿制度,加大惩罚性赔偿数额,进一步打击知识产权侵权行为。构建种业领域的信用机制,建立完善的种子生产与质量监管制度及准入、退出机制。在供给环节,加强对种子企业的品牌营销,包括种子质量、良种示范营销、深化市场细分营销;在需求环节,组建高效技术服务队伍、搭建技术服务培训推广网络。支持中国种子企业通过共建试验站共享种质资源,加强非中国原生农作物种质资源的引进,积极开展种业知识产权的国际布局,联合开展全球化育种与种子生产技术研发,通过种业企业的跨国合并重组等方式提高国际影响力;制定匹配的保险、财政政策,支持种业龙头企业做大做强,深度融入全球农业产业链、价值链和供应链,不断提升种业的生态保障能力、资源保障能力、技术保障能力、市场保障能力、制度保障能力和风险保障能力[18] 。
-
[1] ZUO J, LI J. Molecular dissection of complex agronomic traits of rice: A team effort by Chinese scientists in recent years[J]. Natl Sci Rev, 2014, 1(2): 253-276. doi: 10.1093/nsr/nwt004
[2] 袁隆平. 种业竞争时代的科技创新:超级杂交水稻育种研究新进展[J]. 中国农村科技, 2010(2): 22-25. doi: 10.3969/j.issn.1005-9768.2010.02.006 [3] FITZGERALD M A, MCCOUCH S R, HALL R D. Not just a grain of rice: The quest for quality[J]. Trends Plant Sci, 2009, 14(3): 133-139. doi: 10.1016/j.tplants.2008.12.004
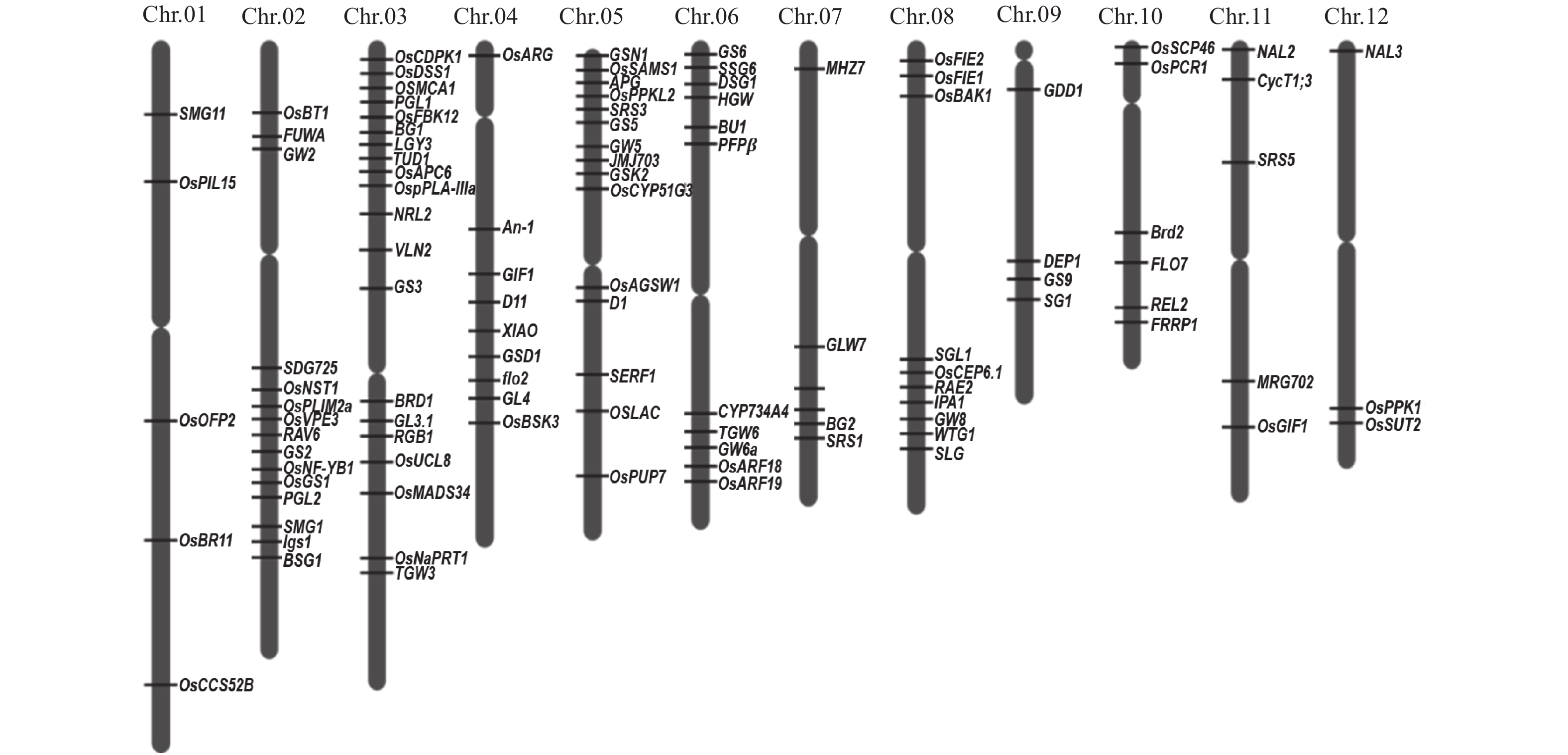
[4] 周开达, 李宏伟. 优质是杂交稻发展的必由之路[J]. 杂交水稻, 1994(z1): 42-45. [5] 肖国樱, 陈芬, 孟秋成, 等. 论湖南水稻育种的主攻方向和技术策略[J]. 杂交水稻, 2015, 30(4): 1-5. [6] HRBERD N P. Shaping taste: The molecular discovery of rice genes improving grain size, shape and quality[J]. J Genet Genomics, 2015, 42(11): 597-599. doi: 10.1016/j.jgg.2015.09.008
[7] XING Y, ZHANG Q. Genetic and molecular bases of rice yield[J]. Annu Rev Plant Biol, 2010, 61(1): 421-442. doi: 10.1146/annurev-arplant-042809-112209
[8] SONG X J, ASHIKARI M. Toward an optimum return from crop plants[J]. Rice, 2008, 1(2): 135-143. doi: 10.1007/s12284-008-9018-3
[9] 杨联松, 白一松, 张培江, 等. 谷粒形状与稻米品质相关性研究[J]. 杂交水稻, 2001(4): 51-53. doi: 10.3969/j.issn.1005-3956.2001.04.024 [10] HUANG R, JIANG L, ZHENG J, et al. Genetic bases of rice grain shape: So many genes, so little known[J]. Trends Plant Sci, 2013, 18(4): 218-226. doi: 10.1016/j.tplants.2012.11.001
[11] 黎毛毛, 徐磊, 刘昌文, 等. 水稻粒形遗传及QTLs定位研究进展[J]. 中国农业科技导报, 2008, 10(1): 34-42. doi: 10.3969/j.issn.1008-0864.2008.01.007 [12] CALINGACION M, LABORTE A, NELSON A, et al. Diversity of global rice markets and the science required for consumer-targeted rice breeding[J]. PLoS One, 2014, 9(1): e85106. doi: 10.1371/journal.pone.0085106.
[13] ZHAO D S, LI Q F, ZHANG C Q, et al. GS9 acts as a transcriptional activator to regulate rice grain shape and appearance quality[J]. Nat Commun, 2018, 9(1). doi: 10.1038/s41467-018-03616-y.
[14] 莫惠栋. 种子性状及其遗传效应的鉴别[J]. 扬州大学学报(农业与生命科学版), 1990, 11(2): 11-15. [15] 林鸿宣, 闵绍楷, 熊振民, 等. 应用RFLP图谱定位分析籼稻粒形数量性状基因座位[J]. 中国农业科学, 1995, 28(4): 1-7. doi: 10.3321/j.issn:0578-1752.1995.04.008 [16] 周清元, 安华, 张毅, 等. 水稻子粒形态性状遗传研究[J]. 西南农业大学学报, 2000(2): 102-104. doi: 10.3969/j.issn.1673-9868.2000.02.002 [17] 谭友斌. 利用高世代回交群体分析水稻粒型QTLs[D]. 武汉: 华中农业大学, 2006. [18] TAN Y F, LI J Y S, XU C G, et al. Genetic bases of appearance quality of rice grains in Shanyou 63, an elite rice hybrid[J]. Theor Appl Genet, 2000, 101(6): 823-829.
[19] 苪重庆, 赵安常. 籼稻粒重及粒形性状F1遗传特性的双列分析[J]. 中国农业科学, 1983, 16(5): 14-20. [20] 石春海, 申宗坦. 籼稻粒形及产量性状的加性相关和显性相关分析[J]. 作物学报, 1996, 22(1): 36-42. doi: 10.3321/j.issn:0496-3490.1996.01.006 [21] 姚国新, 卢磊. 水稻粒重基因定位克隆研究[J]. 安徽农业科学, 2007, 35(27): 8468. doi: 10.3969/j.issn.0517-6611.2007.27.023 [22] 莫惠栋. 我国稻米品质的改良[J]. 扬州大学学报(农业与生命科学版), 1994, 15(1): 8-14. [23] 李欣, 莫惠栋, 王安民, 等. 粳型杂种稻米品质性状的遗传表达[J]. 中国水稻科学, 1999, 13(4): 197-204. [24] 万向元, 陈亮明, 王海莲, 等. 水稻品种胚乳淀粉RVA谱的稳定性分析[J]. 作物学报, 2004(12): 1185-1191. doi: 10.3321/j.issn:0496-3490.2004.12.002 [25] 熊振民, 孔繁林. 大粒型水稻品种的遗传动态及其选育[J]. 浙江农业科学, 1976(2): 26-29. [26] MCKENZIE K S, RUTGER J N. Genetic analysis of amylose content, alkali spreading score, and grain dimensions in rice[J]. Crop Sci, 1983, 23(2): 306-313. doi: 10.2135/cropsci1983.0011183X002300020031x
[27] JUN C L, NUTIK S L. Surgical approaches to intraventricular meningiomas of the trigone[J]. Neurosurgery, 1985, 16(3): 416-420. doi: 10.1227/00006123-198503000-00025
[28] 林荔辉, 吴为人. 水稻粒型和粒重的QTL定位分析[J]. 分子植物育种, 2003(3): 337-342. doi: 10.3969/j.issn.1672-416X.2003.03.007 [29] 石春海, 申宗坦. 早籼粒形的遗传和改良[J]. 中国水稻科学, 1995(1): 27-32. doi: 10.3321/j.issn:1001-7216.1995.01.006 [30] 杨联松, 白一松, 陈多璞, 等. 粳稻粒形遗传初步研究[J]. 杂交水稻, 2002, 17(6): 46-48. doi: 10.3969/j.issn.1005-3956.2002.06.021 [31] 邱先进, 袁志华, 何文静, 等. 水稻垩白性状遗传育种研究进展[J]. 植物遗传资源学报, 2014(5): 992-998. [32] 彭波, 孙艳芳, 李琪瑞, 等. 水稻垩白性状的遗传研究进展[J]. 信阳师范学院学报(自然科学版), 2016, 29(2): 304-312. doi: 10.3969/j.issn.1003-0972.2016.02.035 [33] 国家市场监督管理总局中国国家标准化管理委员会. 大米: GB/T—2018[S]. 北京: 中国标准出版社, 2018:1-10. [34] 徐正进, 陈温福, 马殿荣, 等. 稻谷粒形与稻米主要品质性状的关系[J]. 作物学报, 2004(9): 894-900. doi: 10.3321/j.issn:0496-3490.2004.09.009 [35] 李泽福, 夏加发, 刘礼明, 等. 籼粳交重组自交系群体主要稻米品质性状分析[J]. 安徽农业科学, 2004, 32(6): 1112-1115. doi: 10.3969/j.issn.0517-6611.2004.06.003 [36] 石春海, 朱军. 水稻植株农艺性状与稻米碾磨品质的遗传相关性分析[J]. 浙江农业大学学报, 1997(3): 105-111. [37] 杨联松, 白一松, 许传万, 等. 水稻粒形与稻米品质间相关性研究进展[J]. 安徽农业科学, 2001(3): 312-316. doi: 10.3969/j.issn.0517-6611.2001.03.018 [38] LI Y, FAN C, XING Y, et al. Chalk5 encodes a vacuolar H+-translocating pyrophosphatase influencing grain chalkiness in rice[J]. Nat Genet, 2014, 46(4): 398-404. doi: 10.1038/ng.2923
[39] 李仕贵, 黎汉云, 周开达, 等. 杂交水稻稻米外观品质性状的遗传相关分析[J]. 西南农业大学学报, 1995, 17(3): 197-201. [40] 石春海. 水稻粒形与优质米育种[J]. 中国农学通报, 1994(1): 41-45. [41] ZUO J, LI J. Molecular genetic dissection of quantitative trait loci regulating rice grain size[J]. Annu Rev Genet, 2014, 48: 99-118. doi: 10.1146/annurev-genet-120213-092138
[42] ZHAO H, SUN L, XIONG T, et al. Genetic characterization of the chromosome single-segment substitution lines of O. glumaepatula and O. barthii and identification of QTLs for yield-related traits[J]. Mol Breeding, 2019, 39(4). doi: 10.1007/s11032-019-0960-0.
[43] BHATIA D, WING R A, YU Y, et al. Genotyping by sequencing of rice interspecific backcross inbred lines identifies QTLs for grain weight and grain length[J]. Euphytica, 2018, 214(2). doi: 10.1007/s10681-018-2119-1.
[44] HE N, WU R, PAN X, et al. Development and trait evaluation of chromosome single-segment substitution lines of O. meridionalis in the background of O. sativa[J]. Euphytica, 2017, 213(12). doi: 10.1007/s10681-017-2072-4.
[45] QI L, SUN Y, LI J, et al. Identify QTLs for grain size and weight in common wild rice using chromosome segment substitution lines across six environments[J]. Breeding Sci, 2017, 67(5): 472-482. doi: 10.1270/jsbbs.16082
[46] GAO F, ZENG L, QIU L, et al. QTL mapping of grain appearance quality traits and grain weight using a recombinant inbred population in rice (Oryza sativa L.)[J]. J Integr Agr, 2016, 15(8): 1693-1702. doi: 10.1016/S2095-3119(15)61259-X
[47] LIU D, KANG M, WANG F, et al. Mapping of the genetic determinant for grain size in rice using a recombinant inbred line (RIL) population generated from two elite indica parents[J]. Euphytica, 2015, 206(1): 159-173. doi: 10.1007/s10681-015-1493-1
[48] 刘丹, 王嘉宇, 柴永山, 等. 利用籼粳交RILs群体的水稻粒形QTLs定位[J]. 沈阳农业大学学报, 2015, 46(4): 385-390. doi: 10.3969/j.issn.1008-9713.2015.04.001 [49] 孙滨, 占小登, 林泽川, 等. 水稻粒形和粒重性状的相关性分析及QTL定位[J]. 分子植物育种, 2015, 13(12): 2663-2672. [50] YAN B, LIU R, LI Y, et al. QTL analysis on rice grain appearance quality, as exemplifying the typical events of transgenic or backcrossing breeding[J]. Breeding Sci, 2014, 64(3): 231-239. doi: 10.1270/jsbbs.64.231
[51] ZHANG H, WANG H, QIAN Y, et al. Simultaneous improvement and genetic dissection of grain yield and its related traits in a backbone parent of hybrid rice (Oryza sativa L.) using selective introgression[J]. Mol Breeding, 2013, 31(1): 181-194. doi: 10.1007/s11032-012-9782-z
[52] SINGH R, SINGH A K, SHARMA T R, et al. Fine mapping of grain length QTLs on chromosomes 1 and 7 in Basmati rice (Oryza sativa L.)[J]. J Plant Biochem Biot, 2012, 21(2): 157-166. doi: 10.1007/s13562-011-0080-3
[53] 梅德勇, 朱玉君, 樊叶杨. 籼稻稻米碾磨与外观品质性状的QTL定位[J]. 遗传, 2012(12): 1591-1598. [54] LIANG Y, ZHAN X, GAO Z, et al. Mapping of QTLs associated with important agronomic traits using three populations derived from a super hybrid rice Xieyou9308[J]. Euphytica, 2012, 184(1): 1-13. doi: 10.1007/s10681-011-0456-4
[55] MARATHI B. QTL analysis of novel genomic regions associated with yield and yield related traits in new plant type based recombinant inbred lines of rice (Oryza sativa L.)[J]. BMC Plant Biol, 2012, 12(1): 137. doi: 10.1186/1471-2229-12-137
[56] 张强, 姚国新, 胡广隆, 等. 利用极端材料定位水稻粒形性状数量基因位点[J]. 作物学报, 2011, 37(5): 784-792. [57] 姚国新, 李金杰, 张强, 等. 利用4个姊妹近等基因系群体定位水稻粒重和粒形QTL[J]. 作物学报, 2010, 36(8): 1310-1317. [58] AMARAWATHI Y, SINGH R, SINGH A K, et al. Mapping of quantitative trait loci for basmati quality traits in rice (Oryza sativa L.)[J]. Mol Breeding, 2008, 21(1): 49-65.
[59] 陈冰嬬, 石英尧, 崔金腾, 等. 利用BC2F2高代回交群体定位水稻籽粒大小和形状QTL[J]. 作物学报, 2008(8): 1299-1307. [60] 赵明富, 黄招德, 吴春珠, 等. 水稻谷粒粒长显性主效QTL的遗传分析与定位[J]. 分子植物育种, 2008(6): 1057-1060. doi: 10.3969/j.issn.1672-416X.2008.06.005 [61] YOON D B, KANG K H, KIM H J, et al. Mapping quantitative trait loci for yield components and morphological traits in an advanced backcross population between Oryza grandiglumis and the O. sativa japonica cultivar Hwaseongbyeo[J]. Theor Appl Genet, 2006, 112(6): 1052-1062. doi: 10.1007/s00122-006-0207-4
[62] ZHOU L Q, WANG Y P, LI S G. Genetic analysis and physical mapping of Lk-4(t), a major gene controlling grain length in rice, with a BC2F2 population[J]. Acta Genetica Sinica, 2006, 33(1): 72-79. doi: 10.1016/S0379-4172(06)60011-5
[63] WAN X Y, WAN J M, WENG J F, et al. Stability of QTLs for rice grain dimension and endosperm chalkiness characteristics across eight environments[J]. Theor Appl Genet, 2005, 110(7): 1334-1346. doi: 10.1007/s00122-005-1976-x
[64] MARRI P R, SARLA N, REDDY L V, et al. Identification and mapping of yield and yield related QTLs from an Indian accession of Oryza rufipogon[J]. BMC Genet, 2005, 6. doi: 10.1186/1471-2156-6-33.
[65] ALUKO G, MARTINEZ C, TOHME J, et al. QTL mapping of grain quality traits from the interspecific cross Oryza sativa × O. glaberrima[J]. Theor Appl Genet, 2004, 109(3): 630-639. doi: 10.1007/s00122-004-1668-y
[66] RABIEI B, VALIZADEH M, GHAREYAZIE B, et al. Identification of QTLs for rice grain size and shape of Iranian cultivars using SSR markers[J]. Euphytica, 2004, 137(3): 325-332. doi: 10.1023/B:EUPH.0000040452.76276.76
[67] LI Z F, WAN J M, XIA J F, et al. Mapping quantitative trait loci underlying appearance quality of rice grains (Oryza sativa L.)[J]. Acta Genetica Sinica, 2003, 30(3): 251-259.
[68] YAN C J, LIANG G H, FENG C, et al. Mapping quantitative trait loci associated with rice grain shape based on an indica/japonica backcross population[J]. Acta Genetica Sinica, 2003, 30(8): 711-716.
[69] 汪斌, 兰涛, 吴为人. 应用微卫星图谱定位稻米性状的QTL[J]. 福建农业学报, 2003(1): 11-15. doi: 10.3969/j.issn.1008-0384.2003.01.003 [70] YOSHIDA S, IKEGAMI M, KUZE J, et al. QTL analysis for plant and grain characters of sake-brewing rice using a doubled haploid population[J]. Breeding Sci, 2002, 52(4): 309-317. doi: 10.1270/jsbbs.52.309
[71] 吴长明, 孙传清, 陈亮, 等. 应用RFLP图谱定位分析稻米粒形的QTL[J]. 吉林农业科学, 2002, 27(5): 3-7. doi: 10.3969/j.issn.1003-8701.2002.05.001 [72] 徐建龙, 薛庆中, 罗利军, 等. 水稻粒重及其相关性状的遗传解析[J]. 中国水稻科学, 2002(1): 7-11. [73] BRONDANI C, RANGEL P, BRONDANI R, et al. QTL mapping and introgression of yield-related traits from Oryza glumaepatula to cultivated rice (Oryza sativa L.) using microsatellite markers[J]. Theor Appl Genet, 2002, 104(6): 1192-1203. doi: 10.1007/s00122-002-0869-5
[74] 邢永忠, 谈移芳, 徐才国, 等. 利用水稻重组自交系群体定位谷粒外观性状的数量性状基因[J]. 植物学报, 2001, 43(8): 840-845. doi: 10.3321/j.issn:1672-9072.2001.08.012 [75] MONCADA P, MARTÍNEZ C P, BORRERO J, et al. Quantitative trait loci for yield and yield components in an Oryza sativa×Oryza rufipogon BC2F2 population evaluated in an upland environment[J]. Theor Appl Genet, 2001, 102(1): 41-52. doi: 10.1007/s001220051616
[76] LI J X, YU S B, XU C G, et al. Analyzing quantitative trait loci for yield using a vegetatively replicated F2 population from a cross between the parents of an elite rice hybrid[J]. Theor Appl Genet, 2000, 101(1/2): 248-254.
[77] REDOÑA E D, MACKILL D J. Quantitative trait locus analysis for rice panicle and grain characteristics[J]. Theor Appl Genet, 1998, 96(6/7): 957-963.
[78] HUANG N, PARCO A, MEW T, et al. RFLP mapping of isozymes, RAPD and QTLs for grain shape, brown planthopper resistance in a doubled haploid rice population[J]. Mol Breeding, 1997, 3(2): 105-113. doi: 10.1023/A:1009683603862
[79] LI Z K, PINSON S R M, PARK W D, et al. Epistasis for three grain yield components in rice (Oryza Sativa L.)[J]. Genetics, 1997, 145(2): 453-465.
[80] 尉鑫, 曾智锋, 杨维丰, 等. 水稻粒形遗传调控研究进展[J]. 安徽农业科学, 2019, 47(5): 21-28. doi: 10.3969/j.issn.0517-6611.2019.05.006 [81] SONG X, HUANG W, SHI M, et al. A QTL for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase[J]. Nat Genet, 2007, 39(5): 623-630. doi: 10.1038/ng2014
[82] CHE R, TONG H, SHI B, et al. Control of grain size and rice yield by GL2-mediated brassinosteroid responses[J]. Nat Plants, 2015, 2(1). doi: 10.1038/NPLANTS.2015.195.
[83] DUAN P, NI S, WANG J, et al. Regulation of OsGRF4 by OsmiR396 controls grain size and yield in rice[J]. Nat Plants, 2016, 2(1). doi: 10.1038/NPLANTS.2015.203.
[84] FAN C, XING Y, MAO H, et al. GS3, a major QTL for grain length and weight and minor QTL for grain width and thickness in rice, encodes a putative transmembrane protein[J]. Theor Appl Genet, 2006, 112(6): 1164-1171. doi: 10.1007/s00122-006-0218-1
[85] QI P, LIN Y S, SONG X J, et al. The novel quantitative trait locus GL3.1 controls rice grain size and yield by regulating Cyclin-T1;3[J]. Cell Res, 2012, 22(12): 1666-1680. doi: 10.1038/cr.2012.151
[86] ZHANG X, WANG J, HUANG J, et al. Rare allele of OsPPKL1 associated with grain length causes extra-large grain and a significant yield increase in rice[J]. Proc Natl Acad Sci USA, 2012, 109(52): 21534-21539. doi: 10.1073/pnas.1219776110
[87] LIU Q, HAN R, WU K, et al. G-protein βγ subunits determine grain size through interaction with MADS-domain transcription factors in rice[J]. Nat Commun, 2018, 9(1). doi: 10.1038/s41467-018-03047-9.
[88] SHOMURA A, IZAWA T K, EBITANI T, et al. Deletion in a gene associated with grain size increased yields during rice domestication[J]. Nat Genet, 2008, 40(8): 1023-1028. doi: 10.1038/ng.169
[89] LIU J, CHEN J, ZHENG X, et al. GW5 acts in the brassinosteroid signalling pathway to regulate grain width and weight in rice[J]. Nat Plants, 2017, 3(5). doi: 10.1038/nplants.2017.43.
[90] LI Y, FAN C, XING Y, et al. Natural variation in GS5 plays an important role in regulating grain size and yield in rice[J]. Nat Genet, 2011, 43(12): 1266-1269. doi: 10.1038/ng.977
[91] SUN L, LI X, FU Y, et al. GS6, a member of the GRAS gene family, negatively regulates grain size in rice[J]. J Integr Plant Biol, 2013, 55(10): 938-949.
[92] ISHIMARU K, HIROTSU N, MADOKA Y, et al. Loss of function of the IAA-glucose hydrolase gene TGW6 enhances rice grain weight and increases yield[J]. Nat Genet, 2013, 45(6): 707-713. doi: 10.1038/ng.2612
[93] SONG X J, KUROHA T, AYANO M, et al. Rare allele of a previously unidentified histone H4 acetyltransferase enhances grain weight, yield, and plant biomass in rice[J]. Proc Natl Acad Sci USA, 2015, 112(1): 76-81. doi: 10.1073/pnas.1421127112
[94] WANG Y, XIONG G, HU J, et al. Copy number variation at the GL7 locus contributes to grain size diversity in rice[J]. Nat Genet, 2015, 47(8): 944-948. doi: 10.1038/ng.3346
[95] WANG S, LI S, LIU Q, et al. The OsSPL16-GW7 regulatory module determines grain shape and simultaneously improves rice yield and grain quality[J]. Nat Genet, 2015, 47(8): 949-954. doi: 10.1038/ng.3352
[96] SI L, CHEN J, HUANG X, et al. OsSPL13 controls grain size in cultivated rice[J]. Nat Genet, 2016, 48(4): 447-456. doi: 10.1038/ng.3518
[97] WANG S, WU K, YUAM Q, et al. Control of grain size, shape and quality by OsSPL16 in rice[J]. Nat Genet, 2012, 44(8): 950-954. doi: 10.1038/ng.2327
[98] LI N, XU R, LI Y. Molecular networks of seed size control in plants[J]. Annu Rev Plant Biol, 2019, 70: 435-463. doi: 10.1146/annurev-arplant-050718-095851
[99] JIAO Y, WANG Y D, WANG J, et al. Regulation of OsSPL14 by OsmiR156 defines ideal plant architecture in rice[J]. Nat Genet, 2010, 42(6): 541-544. doi: 10.1038/ng.591
[100] MATSUOKA M, ASHIKARI M. A quantitative trait locus regulating rice grain width[J]. Nat Genet, 2007, 39(5): 583-584. doi: 10.1038/ng0507-583
[101] SHI C, REN Y, LIU L, et al. Ubiquitin specific protease 15 has an important role in regulating grain width and size in rice[J]. Plant Physiol, 2019, 180(1): 381-391. doi: 10.1104/pp.19.00065
[102] WENG J, GU S, WAN X, et al. Isolation and initial characterization of GW5, a major QTL associated with rice grain width and weight[J]. Cell Res, 2008, 18(12): 1199-1209. doi: 10.1038/cr.2008.307
[103] SUN S, WANG L, MAO H, et al. A G-protein pathway determines grain size in rice[J]. Nat Commun, 2018, 9. doi: 10.1038/s41467-018-03141-y.
[104] ASHIKARI M, SAKAKIBARA H, Lin S Y, et al. Cytokinin oxidase regulates rice grain production[J]. Science, 2005, 309(5735): 741-745. doi: 10.1126/science.1113373
[105] XU C, LIU Y, LI Y, et al. Differential expression of GS5 regulates grain size in rice[J]. J Exp Bot, 2015, 66(9): 2611-2623. doi: 10.1093/jxb/erv058
[106] SAKAMOTO T, MATSUOKA M. Identifying and exploiting grain yield genes in rice[J]. Curr Opin Plant Biol, 2008, 11(2): 209-214. doi: 10.1016/j.pbi.2008.01.009
[107] MIURA K, MATSUOKA M. Rice genetics: Control of grain length and quality[J]. Nat Plants, 2015, 1(8). doi: 10.1038/NPLANTS.2015.112.
[108] 万建民. 作物分子设计育种[J]. 作物学报, 2006, 32(3): 455-462. doi: 10.3321/j.issn:0496-3490.2006.03.023 [109] QIAN Q, GUO L, SMITH S M, et al. Breeding high-yield superior quality hybrid super rice by rational design[J]. Natl Sci Rev, 2016, 3(3): 283-294. doi: 10.1093/nsr/nww006
[110] HU J, WANG Y, FANG Y, et al. A rare allele of GS2 enhances grain size and grain yield in rice[J]. Mol Plant, 2015, 8(10): 1455-1465. doi: 10.1016/j.molp.2015.07.002
[111] GONG J, MIAO J, ZHAO Y, et al. Dissecting the genetic basis of grain shape and chalkiness traits in hybrid rice using multiple collaborative populations[J]. Mol Plant, 2017, 10(10): 1353-1356. doi: 10.1016/j.molp.2017.07.014
[112] GAO X, ZhANG X, LAN H, et al. The additive effects of GS3 and qGL3 on rice grain length regulation revealed by genetic and transcriptome comparisons[J]. BMC Plant Biol, 2015, 15. doi: 10.1186/s12870-015-0515-4.
[113] ZENG D, TIAN Z, RAO Y, et al. Rational design of high-yield and superior-quality rice[J]. Nat Plants, 2017, 3(4). doi: 10.1038/nplants.2017.31.
[114] 黄海祥, 钱前. 水稻粒形遗传与长粒型优质粳稻育种进展[J]. 中国水稻科学, 2017, 31(6): 665-672. [115] 杨梯丰, 曾瑞珍, 朱海涛, 等. 水稻粒长基因GS3在聚合育种中的效应[J]. 分子植物育种, 2010, 8(1): 59-66. [116] DAI Z, LU Q, LUAN X, et al. Development of a platform for breeding by design of CMS lines based on an SSSL library in rice (Oryza sativa L.)[J]. Euphytica, 2015, 205(1): 63-72. doi: 10.1007/s10681-015-1384-5
-
期刊类型引用(0)
其他类型引用(2)

 下载:
下载:
计量
- 文章访问数: 2182
- HTML全文浏览量: 9
- PDF下载量: 2371
- 被引次数: 2
