
Regulation mechanism of Mycobacterium bovis PtpA protein on NF-κB signaling pathway
-
摘要:目的
研究牛分枝杆菌Mycobacterium bovis PtpA蛋白对免疫应答相关的NF-κB信号通路的影响,以揭示PtpA蛋白在机体免疫应答中的作用。
方法构建PtpA基因真核表达载体FLAG-PtpA,并将其转染到HEK293T中,进行SDS-PAGE分析及Western blot检测。激活NF-κB信号通路后,通过双荧光素酶试验和qPCR方法探究PtpA蛋白对NF-κB信号通路的影响。
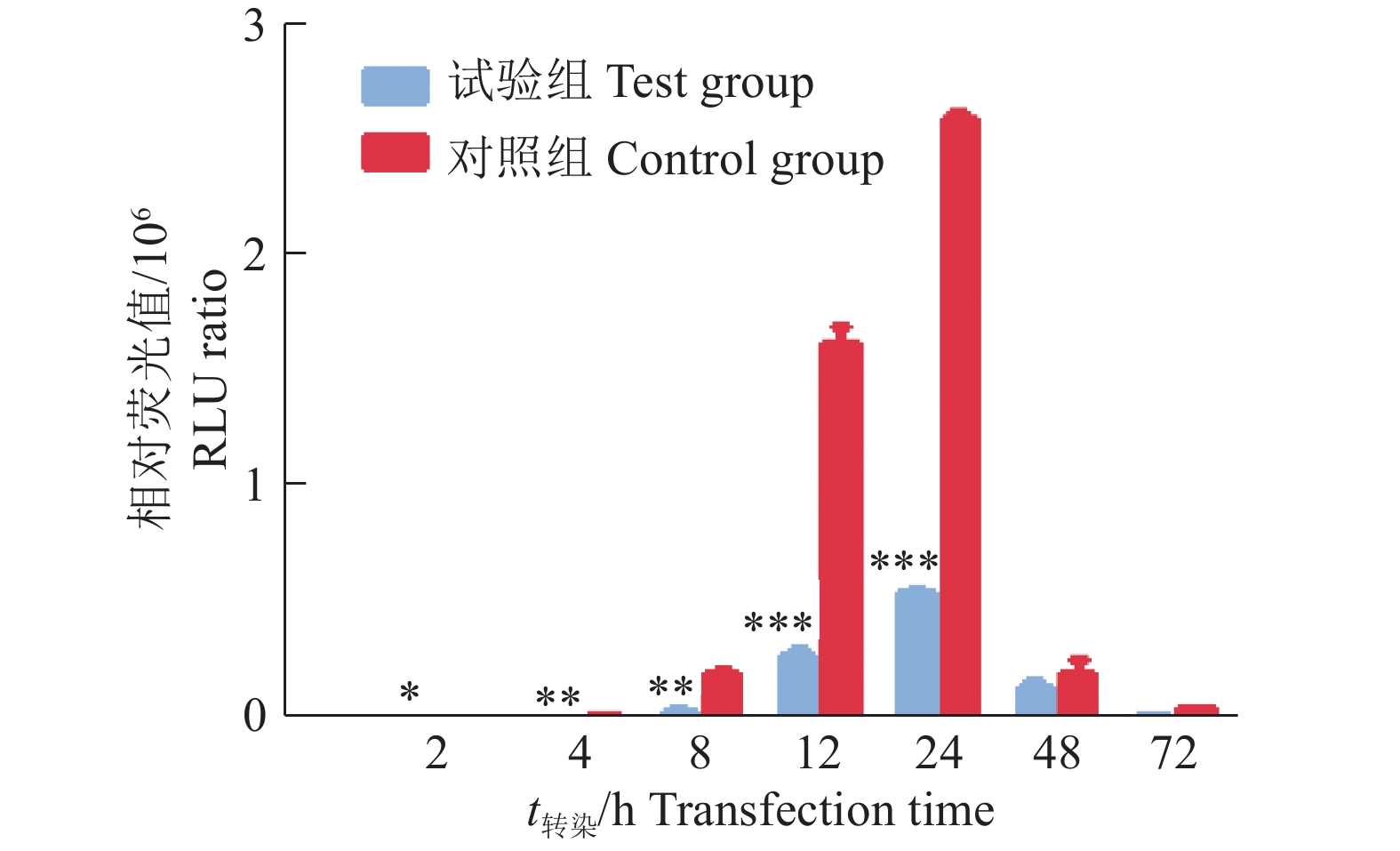
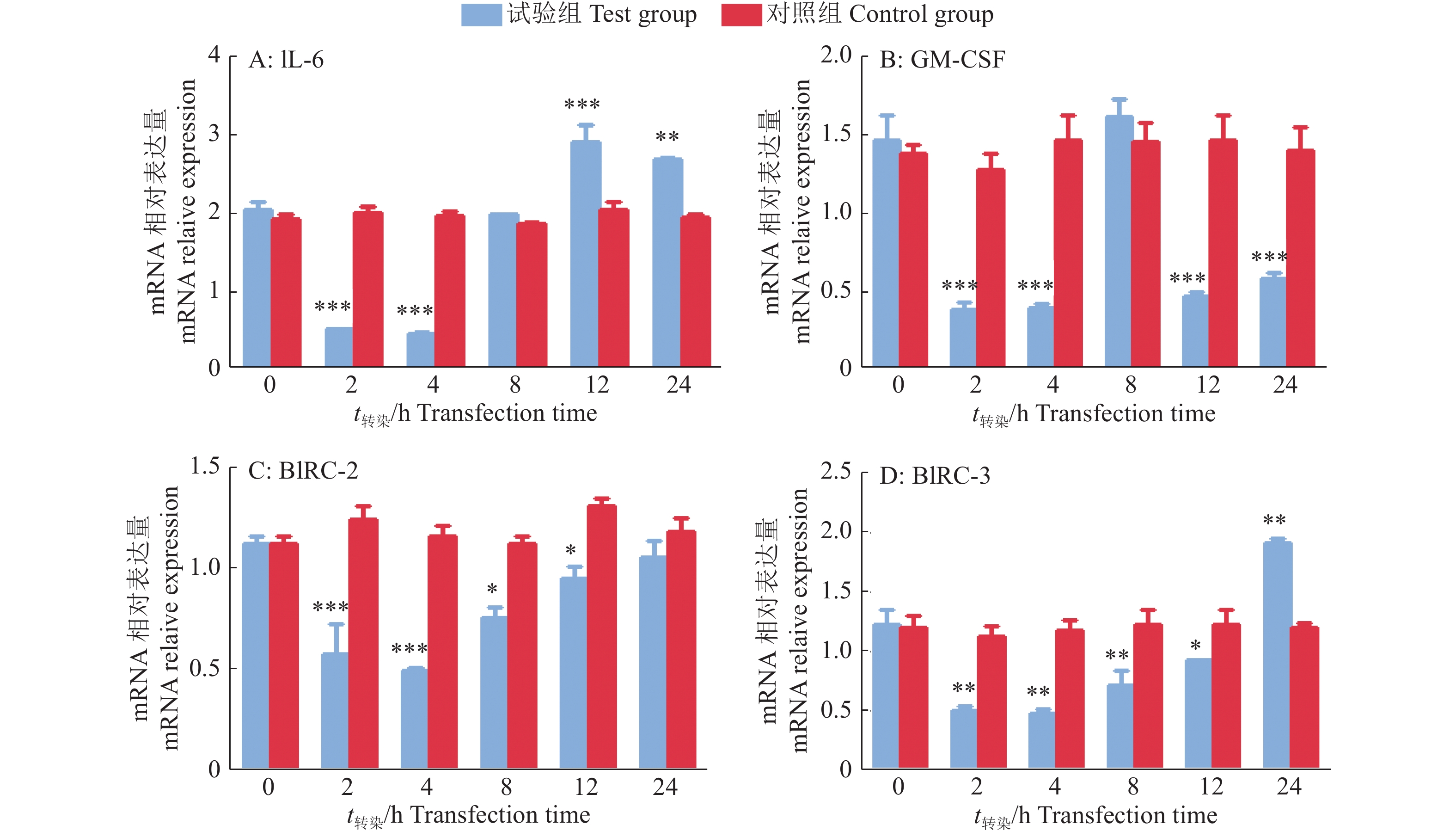
结果成功构建PtpA基因真核表达载体FLAG-PtpA,转染HEK293T后经SDS-PAGE分析,相对分子质量约为22 000处可见特异性蛋白条带。Western blot结果显示,表达产物可与一抗特异性结合,证明该蛋白是PtpA蛋白。双荧光素酶试验中,转染2~24 h,试验组与对照组萤火虫荧光素酶和海肾荧光素酶的相对荧光强度的比值差异显著(P<0.05);转染2 h后,对照组萤火虫荧光素酶和海肾荧光素酶的相对荧光值是试验组的2.93倍,说明PtpA蛋白对NF-κB信号通路激活早期具有显著的抑制作用。qPCR结果显示,转染2 h后,对照组的IL-6、GM-CSF、BIRC-2和BIRC-3的表达量分别是试验组的3.93、3.42、2.17和2.30倍(P<0.01);转染4 h后,对照组的IL-6、GM-CSF、BIRC-2和BIRC-3的表达量分别是试验组的4.26、3.93、2.36和2.50倍(P<0.01),说明PtpA蛋白对NF-κB信号通路相关的细胞因子(IL-6、GM-CSF、BIRC-2、BIRC-3)在免疫早期具有显著抑制作用。
结论qPCR结果与双荧光素酶试验结果一致,表明牛分枝杆菌PtpA蛋白对NF-κB信号通路的影响主要发生在早期。本研究为后续研究有效的结核病防控药物提供了理论基础。
Abstract:ObjectiveTo investigate the effect of Mycobacterium bovis PtpA protein on NF-κB signaling pathway which is related to immune response, and reveal the role of PtpA protein in the body's response to immunity.
MethodThe PtpA gene eukaryotic expression vector FLAG-PtpA was transfected into HEK293T for SDS-PAGE analysis and Western blot detection. After activation of NF-κB signaling pathway, the effects of PtpA protein on NF-κB signaling pathway were investigated by dual luciferase assay and qPCR.
ResultThe eukaryotic expression vector FLAG-PtpA of PtpA gene was successfully constructed, transfected into HEK293T and analyzed by SDS-PAGE. The specific molecular band was visible with relative molecular mass of 22 000. Western blot results showed that the expression product specifically bound to the primary antibody, demonstrating that the protein was a PtpA protein. In the double luciferase assay, the ratios of the relative fluorescence intensities of firefly luciferase and Renilla luciferase in the test group and the control group were significantly different from 2 to 24 h after transfection (P<0.05). The relative fluorescence values of firefly luciferase and Renilla luciferase in the control group were 2.93 times higher than those in the experimental group 2 h after transfection, indicating that PtpA protein had a significant inhibitory effect on the early activation of NF-κB signaling pathway. qPCR results showed that the expression levels of IL-6, GM-CSF, BIRC-2 and BIRC-3 in the control group were 3.93, 3.42, 2.17 and 2.30 times respectively of those in the test group 2 h after transfection(P<0.01), and were 4.26, 3.93, 2.36 and 2.50 times respectively of those in the test group 4 h after transfection(P<0.01). These results indicated that PtpA protein had a significant inhibitory effect on NF-κB signaling pathway-associated cytokines (IL-6, GM-CSF, BIRC-2 and BIRC-3) in the early stage of immunization.
ConclusionqPCR results are consistent with the results of dual luciferase assay, indicating that the effect of M. bovis PtpA on NF-κB signaling pathway mainly occurs in the early stage. This study provides a theoretical basis for the follow-up study of effective tuberculosis prevention and control drugs.
-
Keywords:
- bovine tuberculosis /
- Mycobacterium bovis /
- PtpA protein /
- NF-κB signaling pathway /
- cytokine /
- immune response
-
水稻Oryza sativa L.是极为重要的粮食作物,世界上超过50%的人口以大米为主食[1]。水稻半矮化育种和籼型杂交水稻的应用带来了水稻产量的2次巨大飞跃,为保障我国的粮食安全作出极大贡献[2-3];但长期高度重视产量的育种也导致了大部分高产品种品质相对较差。随着社会经济的发展,人们对稻米,特别是优质稻米的需求量日益增多,高产优质成为当下水稻育种的重要目标之一[4]。
稻米品质主要包括碾磨品质、外观品质、营养品质、蒸煮食味品质4个方面[5];这4个品质性状直接决定稻米的商品价值及群众的消费行为,其中稻米在蒸煮后带给消费者的感官体验更是决定消费者购买的关键因素[6-7]。稻米蒸煮食味品质是精米在蒸煮与食用过程中所呈现的各种理化性质及感官特征,主要包括米饭的外观、气味、口感、硬度、黏性、冷饭质地等[8]。在稻米蒸煮过程中,米粒经吸水蒸煮后发生纵向延伸和横向膨胀[9-10],这种米粒蒸煮时的延伸特性称为饭粒延伸性(Cooked rice elongation,CRE),如图1所示,以蒸煮后饭粒长度增加值与蒸煮前米粒长度的比值来衡量。优质稻米往往在蒸煮时纵向延伸明显,而横向伸长较少。因此,饭粒延伸性被认为是优质稻米,特别是优质籼米的重要评价指标之一[11-12]。
长期以来,对稻米蒸煮食味品质的研究重点都集中于直链淀粉含量、胶稠度和糊化温度这3个指标[13]。尽管饭粒延伸性是评价稻米蒸煮食味品质的重要指标之一,但受限于该性状遗传基础复杂,且易受环境和加工条件影响,鉴定方法复杂,目前对饭粒延伸性这一性状的遗传机理及调控网络所知甚少。本文就饭粒延伸性的研究方法和研究进展进行综述,以期为挖掘优异饭粒延伸性基因位点、培育优质水稻新品种提供理论依据。
1. 饭粒延伸性相关评价指标
用于评价米饭延伸特性的相关指标较多,主要包括精米长、精米宽、米粒长宽比、饭粒长、饭粒宽、饭粒长宽比、饭粒延伸性、饭粒伸长率、米粒延伸指数、米粒膨胀率[9, 12-47]等。衡量米粒蒸煮时延伸特性主要有饭粒延伸性、米粒延伸指数和饭粒伸长率3个指标。
饭粒延伸性=(饭粒长−米粒长)/米粒长[16, 18-20, 22, 24, 26, 30, 33, 36-37, 41, 43-45, 47-48],
米粒延伸指数=饭粒长宽比/米粒长宽比[9, 14, 27-28, 32, 36],
饭粒伸长率=饭粒长/米粒长[17, 20-21, 25, 27-29, 31-32, 34-35, 38-39, 46, 49]。
2. 饭粒延伸性测定方法
准确测量和评价饭粒延伸性受到研究人员的高度重视,多种米饭样品制备方法被用于试验探究。在早期,通常用游标卡尺测定蒸煮前后的米粒长度与饭粒长度,然后根据测量值进行计算。常见的方法是将米粒在室温条件下浸泡30 min,然后直接将浸泡过的米粒转移到沸水浴中煮制10 min,冷却后立即测量米饭长度;与其他米饭制备方法相比,浸泡30 min可有效缩短米饭样品制备时间,更为便捷[14]。在具体测定中,通常任取10粒整精米,用游标卡尺测量米粒长度后,将10粒米装入相同型号的试管中再利用沸腾的水浴锅煮成饭粒,取出饭粒在滤纸上静置3 h以上,测定长度;饭粒延伸性依据公式“(饭粒长-米粒长)/米粒长”计算,重复测定3次取平均值。
多粒稻米同时在同一器皿中蒸煮会导致一定程度上的黏连,而且蒸煮所用的开放性器具也难以保证饭粒加工环境的一致性,在客观上影响了饭粒延伸度测定的准确度和可重复性。Okpala等[37, 41, 44]和Potcho等[45]改进了制作饭粒的方法,利用PCR仪实现单粒制作饭粒。具体为一次测量20粒精米,将每粒精米转移到预先加有150 μL蒸馏水的PCR板孔上,浸泡30 min,将其放入PCR热循环器中,设定程序为98 ℃ 20 min、20 ℃ 5 min,然后将煮熟的米粒从PCR板中取出,放在滤纸上,室温干燥5 min后测量,重复测定3次。
游标卡尺测量需要大量的人工操作,不仅耗时费力,而且不同批次材料、不同人员在操作规范上难以完全统一,使得饭粒延伸性的测量效率低、误差大、重复性差。随着数字图像技术的发展,图片分析软件被用于测量谷物的长度和宽度,这显著提高了测量效率,减少了试验操作间的主观误差[50-51]。Amarawathi等[25]使用CCD相机首先校准游标卡尺和放大照片进行读数,再使用软件Image Pro Plus software version 4.1自动估计10种谷物烹饪前后的长度。Jinorose等[52]将米粒表观特征的图像分析与米粒的物理化学和蒸煮特性相关联,监测稻米加工和蒸煮过程,利用蒸煮前后米粒图像参数来评价与饭粒延伸性相关的各项指标。有研究使用扫描仪(Microtek ScanMaker系列)进行蒸煮前后所有长度的测量,经软件分析得到结果,消除了手动测量和视觉观察的主观误差[36-37, 41, 44-45],这也成为近年来测量饭粒延伸性的主要方法。
3. 稻米品质性状与饭粒延伸性的关系
3.1 淀粉对饭粒延伸性的影响
淀粉是水稻胚乳的主要成分,占80%左右,主要由直链淀粉和支链淀粉组成。直链淀粉是由α−1,4糖苷键连接的葡萄糖组成的线性分子,通常用直链淀粉含量、凝胶稠度和糊化温度3个理化特性来衡量,是影响精米加工质量和蒸煮的重要指标[53-54]。支链淀粉是通过α−1,4糖苷键连接成主链,加上由β−1,6糖苷键连接的支链共同构成分枝的多聚体[54]。直连淀粉与支链淀粉共同组成淀粉的精细结构,与稻米的理化性质和品质密切相关[55-56]。
优质稻米在蒸煮时一般纵向伸长较多,而横向伸长较少[12]。淀粉精细结构中的淀粉粒排列取向和紧密程度会影响米粒吸水膨胀过程,使之产生纵向特定方向的伸展,从而影响籽粒的伸长率[19, 39]。已知高含量直链淀粉可通过与脂质形成复合体来抑制颗粒膨胀和直链淀粉浸出,增加水解酶的颗粒阻力,从而抑制烹饪过程中的膨胀现象[57-59]。在高水分含量蒸煮过程中,直链淀粉含量较高的稻米的延伸率高于直链淀粉含量较低的稻米的延伸率[53]。支链淀粉的结构对熟饭的适口性有重要影响[40],支链淀粉的长支链在淀粉粒外层形成梳状结构和b型结晶度,这种梳状结构可以抑制沸腾过程中饭粒的膨胀和糊化,从而影响饭粒延伸率[60-61]。
3.2 蛋白质含量对饭粒延伸性的影响
蛋白质是稻米胚乳的第二大组分,占8%左右,胚乳中的蛋白主要是谷蛋白和醇溶蛋白,蛋白质含量是影响稻米食用品质的最重要因素之一,低蛋白质含量稻米往往具有更好的食味品质[54, 62]。蛋白质以蛋白体形式填充在淀粉粒之间,会影响米饭的黏性和硬度,是影响淀粉吸水糊化的主要因子之一[63-64]。一般认为稻米的蛋白质含量与食味品质呈负相关[65],蛋白质可能通过影响淀粉的糊化特性来影响稻米的蒸煮品质[66],高蛋白质含量会抑制淀粉的糊化及籽粒的吸水率,从而影响稻米的蒸煮特性[67-69]。蛋白质对淀粉颗粒具有保护作用,稻米外层的蛋白质作为淀粉晶体结构的网络骨架,会阻隔水分的迁移并抑制淀粉的糊化和膨胀[70-72]。稻米蛋白质还可与淀粉相互作用,形成网状结构,增强淀粉的溶胀性,从而改变蒸煮过程中米粒的膨胀,进而影响米饭的延伸率[73]。
3.3 外观品质对饭粒延伸性的影响
稻米外观品质包括粒宽、粒长、垩白度等评价指标。外观品质的大部分性状与米饭延伸性状存在显著相关[43],米饭延伸率与精米长呈极显著负相关,与饭粒长宽比、精米宽、饭粒长、饭粒宽、米饭膨胀率、饭粒质量及米饭吸水性这些性状呈极显著正相关,部分QTLs同时控制米饭延伸率和其他米粒品质性状[9, 18, 20, 22, 24-26, 29-32, 36, 43, 46-49]。张光恒等[18]定位到控制米饭延伸率的基因座位同时控制饭粒长,说明饭粒长与米粒延伸性可能密切相关。何予卿等[9]检测到一个同时决定米粒长宽比、米饭长宽比和米饭延伸指数的QTL,表明这几个性状存在一定程度的相关性。研究发现,蒸煮过程中影响籽粒长、宽伸长的主效QTL与籽粒本身的长度和宽度无关[29],这表明,饭粒延伸性并非由米粒宽度和长度直接决定,而是受到其他独立的遗传因素影响。
水稻籽粒垩白是指胚乳中不透明的部分,通常由不规则的淀粉颗粒或异常的蛋白质体组成,垩白对稻米碾磨、蒸煮、食用和籽粒外观都有不利影响[74-76]。垩白通过影响籽粒的质构特性及加工过程从而影响饭粒延伸性。高垩白米粒与低垩白米粒的微观结构明显不同,高垩白米粒的胚乳中充满了排列疏松的球形淀粉颗粒,具有较大的间隙,低垩白米粒由密集且堆积规则的多面体淀粉颗粒组成[74, 76]。低垩白籽粒和高垩白籽粒质构特性的差异可能是直链淀粉含量、长/短链支链比和粒状结构的差异所致。直链淀粉含量较低的高垩白籽粒可能具有较少的无定形区域和较多的结晶区域,从而增加糊化温度和吸热焓;高垩白籽粒稻米质构特性改变,降低了稻米的蒸煮质量,导致蒸煮时间较短,米饭质地较硬、黏性较小,且饭粒延伸率比低垩白米粒的小,可能是淀粉含量和颗粒结构的差异导致了较低的膨胀度[77-79]。垩白米粒加工过程容易发生断裂,影响整精米率,从而影响饭粒的完整性。
3.4 碾磨品质对饭粒延伸性的影响
碾磨品质是决定水稻品种市场价值和生产效益的重要指标之一。人们普遍认为碾米度是决定碾米品质的关键因素,碾米度也称为加工精度,指稻米籽粒表面除去糠皮的程度,它影响稻米的感官、蒸煮和质构品质[80-82]。
碾米度通过淀粉结构影响米饭的质构,从而影响饭粒延伸性。随着碾米度的增加,与米饭食味品质相关的直链淀粉含量、胶稠度和糊化温度大多显著或极显著提高。碾磨度通过影响淀粉结构和其他非淀粉多糖的含量来极大地影响稻米的质地特征。随着碾磨度增加,最佳蒸煮时间减少,最佳蒸煮时间显著影响饭粒的长度、膨胀率和黏聚性;高碾磨度可以提高稻米的吸水性、溶胀性,从而提高米饭感官品质。为稻米品种选择合适的碾米度,能够最大限度地减少加工损失,提高稻米的蒸煮和食用品质[83-85]。
4. 影响饭粒延伸性的主要因素
4.1 环境因素对饭粒延伸性的影响
除了受淀粉特性、蛋白质含量、外观品质和碾磨品质等内在理化性质影响,饭粒延伸性与水稻栽培环境和米饭制备过程等环境因素也息息相关。
稻米中氨基酸、蛋白质的分布和支链淀粉中的支链长度在很大程度上受氮肥影响[68-69, 86]。氮肥可以通过改变米粒中蛋白质和淀粉的结构,最终影响稻米的蒸煮品质[68]。不同氮源对水稻的饭粒伸长性的影响可能有所不同。有研究表明,与采用碳酸氢铵(NH4HCO3)、尿素(H2NCONH2)和硝酸钠(NaNO3)作为氮源相比,施加硝酸钾(KNO3)水稻的饭粒延伸率与其差异极显著[45]。这也为通过改良栽培条件提高稻米食味品质提供了一定的理论依据。
灌浆期的环境温度和加工储存条件对饭粒延伸率也存在一定的影响。外界温度等气候条件可能通过影响淀粉粒的结构、代谢产物中环己醇的含量,进而影响饭粒延伸性和蒸煮特性[14, 40];但不同品种对温度的反应不一样[44],这也表明饭粒延伸性还受遗传因素影响。在稻米储存过程中,可能发生支链淀粉水解,这将导致淀粉颗粒在糊化过程中的溶胀度变高;随着精米储藏时间延长,饭粒延伸性随之增加[82]。人工老化处理(90 ℃处理3 h)亦能增加饭粒延伸性,提高稻米蒸煮品质[46]。
饭粒延伸受蒸煮过程中糊化温度的影响[87]。在相似的直链淀粉含量条件下,不同糊化温度材料的饭粒延伸特性不同[29]。糊化峰值温度、糊化峰值时间与饭粒延伸性呈极显著正相关[26]。蒸煮加工条件也是影响饭粒延伸性的因素之一。米饭样品制备过程中,蒸煮会导致稻米中淀粉、蛋白质等组分发生物质迁移和结构变化,最终影响米饭的外观、质地等食味品质特征[14, 88-89]。蒸煮行为与籽粒的溶胀特性有关,溶胀又受到糊化特性影响[53]。淀粉糊化温度还是决定稻米沸水蒸煮速度的主要因素,沸水蒸煮速度也会影响饭粒延伸率的测定[14]。在蒸煮过程中,蒸煮米水比和吸水率对稻米蒸煮影响极大,低直链淀粉含量品种蒸煮过程中需水量较少,根据不同稻米材料控制加水量能够提高米饭外观品质和食味品质[69, 88]。蒸煮前的浸泡也影响饭粒延伸性。米粒吸水发生膨胀,以浸泡膨胀率来衡量,较高的浸泡温度可以在一定时间内增加精米含水量,但可能会降低浸泡膨胀率[72]。饭粒延伸率与吸水率和膨胀率呈极显著正相关[47]。但是,饭粒延伸率不等同于浸泡膨胀率,这是2个具有密切关系的不同性状[30, 36-37],尽管有研究在同一个区间同时检测到控制米粒浸泡膨胀率和饭粒延伸率的QTL[30]。
4.2 遗传因素对饭粒延伸性的贡献
饭粒延伸性用饭粒延伸长度与米粒长的比值表示;米粒延伸指数用米粒长宽比与饭粒长宽比的比值表示,这一性状在一定程度上综合反映了饭粒形状和米粒形状之间的关系[9, 14]。饭粒延伸性相关的性状均属于多基因控制的数量遗传性状,加性效应和显性效应对饭粒延伸性状的遗传起主要作用[16, 36, 43]。饭粒延伸性受到基因型与环境互作效应的控制,狭义遗传率偏低,对其进行早代选择效果不佳,通常需要通过反复选择来改良这一性状[9, 15-16, 18-19, 22, 28, 30, 46, 90]。深入研究控制饭粒延伸性的遗传位点,开发高效分子标记,将有利于提高早期世代的选择效率,加速优质稻米新品种的选育。
前人的研究认为,控制米粒延伸性的等位基因多来自籼稻,在粳稻上也存在正效等位基因,说明控制饭粒延伸性的QTL客观存在,并且在水稻籼粳分化过程中未被丢失[9, 15-16, 18-19, 21, 25, 27, 49]。利用籼稻品种‘Basmati’‘Neda’‘窄叶青8’‘密阳46’‘珍汕97’‘Pusa112’‘台中本地1号’‘IR641’‘93-11’[16-18, 20-21, 24-26, 29-30, 32-34, 36-39, 45-46, 49]、粳稻品种‘沈农265’‘丽江新团黑谷’‘Asominori’‘春江06’‘越光’[27-28, 30, 47]和O. nivara IRGC[31]等材料定位了大量与饭粒延伸性相关的QTLs。其中,‘Basmati’为国际著名优质稻,蒸煮过程中具有极高的线性籽粒伸长和最低的横向膨胀,饭粒长度高达22 mm,饭粒延伸率高达2.5,是研究饭粒延伸性遗传基础和改良饭粒延伸性极佳的材料资源[38-39]。
评价水稻饭粒延伸性主要有饭粒延伸性、米粒延伸指数、饭粒伸长率3个指标。围绕这3个主要指标,已有70多个相关QTLs被定位,包括38个饭粒延伸性QTLs[18-20, 26, 29-30, 33, 43, 47]、10个米粒延伸指数QTLs[9, 19, 28, 32, 36, 43]和28个饭粒伸长率QTLs[17, 20, 25, 27-30, 32, 34-35, 39, 49]。
目前,被鉴定的饭粒延伸性相关QTLs在水稻的12条染色体上均有分布。以在第6号染色体上定位到的QTL数量最多,超过10个;多数染色体上有2~3个相关QTLs被定位,第5号染色体上被定位的QTL最少,仅有1个(图2)。控制水稻直链淀粉含量的主效基因Wx和影响糊化温度的主效基因ALK均位于第6号染色体短臂末端。水稻饭粒延伸性与直链淀粉含量、糊化温度等相关性状关系密切,被定位于第6号染色体短臂末端的多个饭粒延伸性相关QTLs所在区间与Wx和ALK临近或重合[9, 18-19, 26, 29-30, 32-33, 36, 39, 43, 91]。在已定位饭粒延伸性相关QTL区间内,还覆盖了与支链淀粉生物合成相关的SSIIa、SBEIIa和SBEIII基因,这表明饭粒延伸性与支链淀粉密切相关[39]。此外,尽管采用不同的衡量指标,有的区间反复被不同的学者鉴定到携带饭粒延伸性相关QTL,例如,第2号和第4号染色体的多个QTLs存在重叠定位区间[18-20, 26, 28, 32-33, 35, 39],这暗示相关区间可能存在影响饭粒延伸性的主效位点。
![]() 图 2 部分已鉴定的水稻饭粒延伸性QTLsChr.:染色体;黑色、红色和绿色条块分别代表饭粒延伸性、饭粒伸长率和米粒延伸指数的QTL,条块位置代表QTL的估计位置;蓝色为已克隆淀粉相关基因的物理位置Figure 2. Partial identified QTLs for cooked rice elongationChr.: Chromosome; The bars in black, red, and green represent QTLs for cooked rice elongation, cooked rice elongation rate, and rice grain elongation index, respectively, the location of the bar represents the estimated location of the QTL; Blue is the physical location of the cloned starch-related genes
图 2 部分已鉴定的水稻饭粒延伸性QTLsChr.:染色体;黑色、红色和绿色条块分别代表饭粒延伸性、饭粒伸长率和米粒延伸指数的QTL,条块位置代表QTL的估计位置;蓝色为已克隆淀粉相关基因的物理位置Figure 2. Partial identified QTLs for cooked rice elongationChr.: Chromosome; The bars in black, red, and green represent QTLs for cooked rice elongation, cooked rice elongation rate, and rice grain elongation index, respectively, the location of the bar represents the estimated location of the QTL; Blue is the physical location of the cloned starch-related genes5. 饭粒延伸性的设计育种
广东省植物分子育种重点实验室张桂权教授团队(本团队)创建了以‘华粳籼74’为单一受体亲本的水稻染色体单片段代换系(Single segment substitution line,SSSL)文库,该文库现有2 360份材料,覆盖水稻染色体近30倍,所有SSSL遗传背景与受体亲本近乎一致[92]。SSSL是基因定位、克隆,基因互作效应分析乃至设计育种的极佳材料。本团队已利用该材料定位并克隆了一系列控制水稻重要农艺性状的复杂QTL位点,包括粒形、垩白、柱头外露率、抽穗期等[93]。SSSL的运用将有助于解析水稻饭粒延伸性相关遗传位点,本团队已利用该材料文库连续多年多点定位到11个控制饭粒延伸率的QTLs,其加性效应值为0.57~10.44,这为相关基因的克隆奠定了坚实的基础。通过常规杂交和分子标记辅助选择,利用鉴定到的携带饭粒延伸性QTL的SSSL发展多片段聚合系材料,获得了携带不同QTL数目的聚合系。多片段聚合系的饭粒延伸性均显著优于受体亲本‘华粳籼74’,并且随着QTL聚合数目的增加,聚合系的饭粒延伸率总体呈现平稳上升趋势(图3)。这为深入解析饭粒延伸性的遗传基础,并将优异基因资源运用于优质稻米的设计改良提供了理论依据和种质资源。
6. 问题及展望
饭粒延伸性是衡量稻米蒸煮食味品质的重要指标。数十年来,人们对水稻饭粒延伸性及其影响因素做了大量的探讨,并逐步改进该性状的测定方法,取得了一定的进展。进入21世纪以来,随着水稻功能基因组研究的快速发展,越来越多的水稻重要农艺性状基因被挖掘,并被用于水稻品种设计改良;但受限于性状的复杂性,目前对水稻饭粒延伸性的遗传研究还相当有限,这严重限制了基于饭粒延伸率提高的水稻品质改良。
饭粒延伸性是受多基因控制,且受环境因素影响极大的数量性状遗传位点。在前期的研究中,不同研究者通过发展重组自交系(Recombinant inbred line,RIL)[9, 19-20, 25-26, 28, 36]、F2[16, 21, 34, 39]、F3[49]、F6[49]、F2:3[27]、双单倍体(Doubled haploid,DH)[18, 29-30]、染色体片段代替系(Chromosome segment substitution line,CSSL)[28, 33, 47]、BC2F3[31]、BC3F1[17]、BC3F2[36]和回交自交系(Backcross inbred line,BIL)[32]等遗传群体定位与饭粒延伸性相关的QTL,也有利用3K RGP(3 000 rice genomes project)等自然材料进行全基因组关联分析[35, 43]。所用的材料遗传背景复杂,差异较大,相关QTL定位区间较大,甚至难以确保QTL定位的可靠性,这在客观上造成了鲜见控制饭粒延伸性的功能基因被报道。
克隆水稻饭粒延伸性基因并且对其功能进行解析、阐明水稻饭粒延伸性的遗传调控网络、探讨水稻饭粒延伸性与其他稻米品种性状的遗传互作和协同改良将成为未来水稻饭粒延伸性相关研究的主要方向与目标。
-
![]()
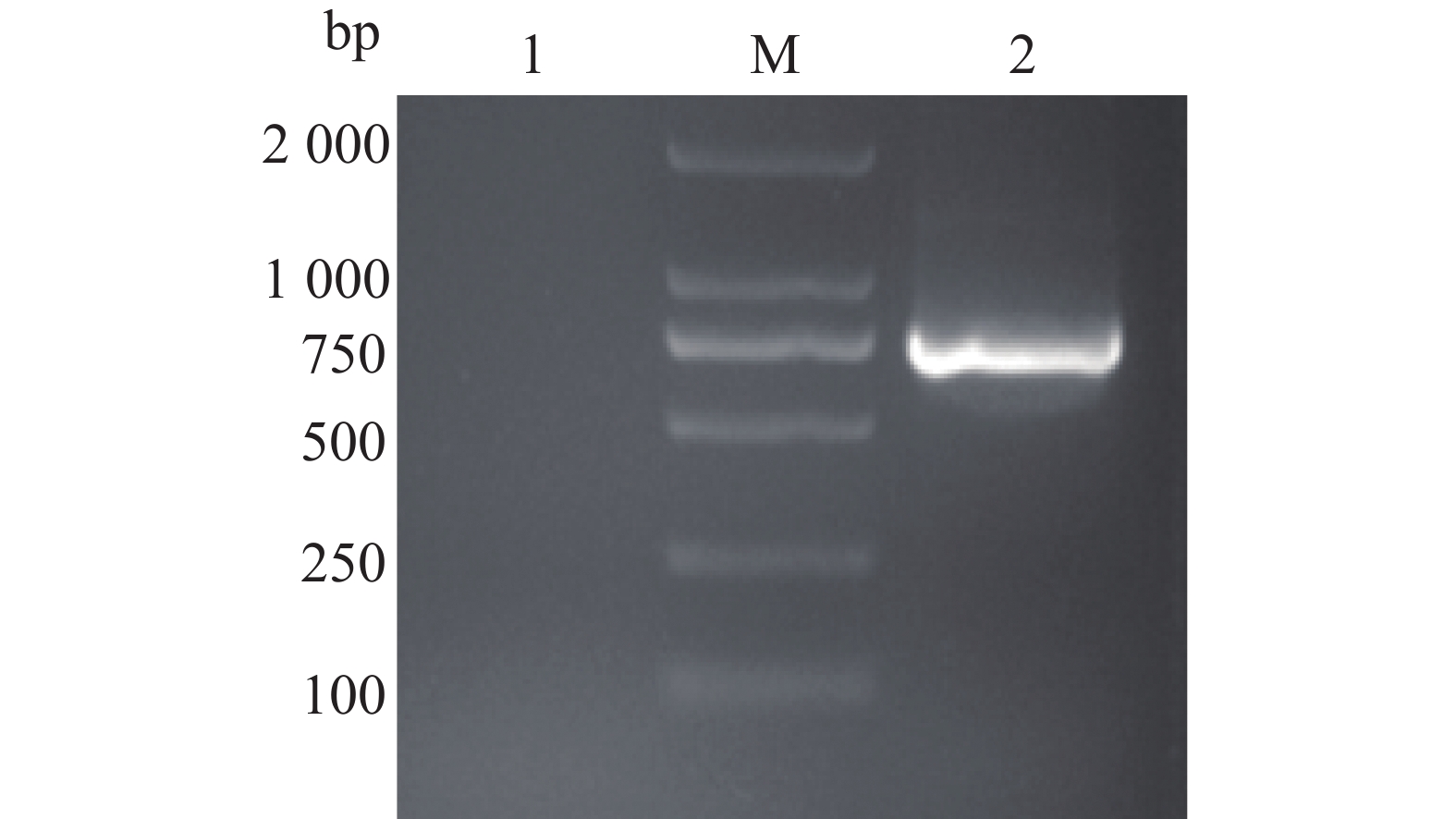
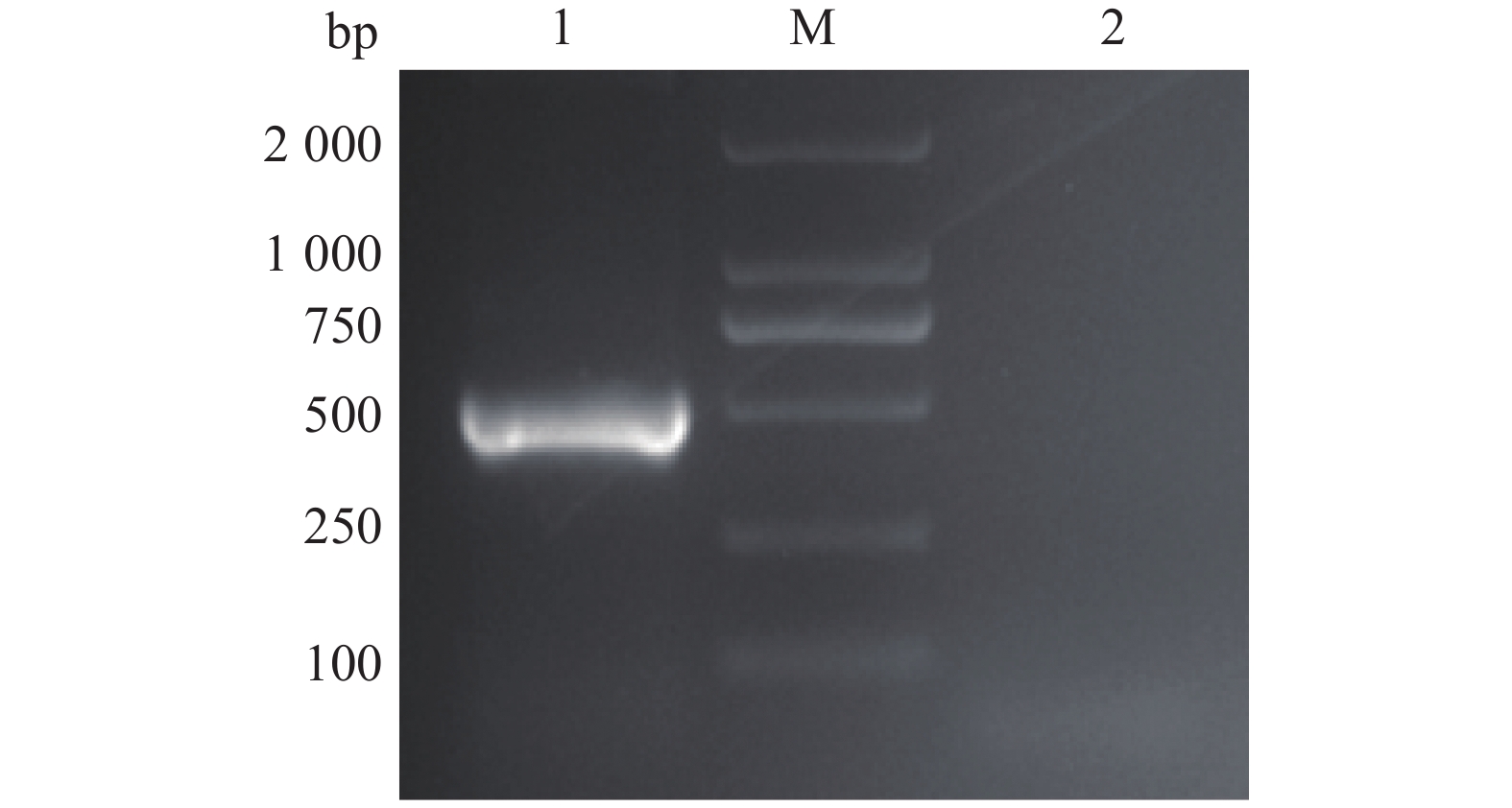
图 1 PtpA基因的PCR扩增
1:PtpA基因的PCR扩增产物;M:DL2000 DNA marker;2:阴性对照
Figure 1. PCR amplification of the PtpA gene
1: PCR product of PtpA gene; M: DL2000 DNA marker;2: Negative control
![]()
图 2 FLAG-PtpA重组质粒中PtpA基因的PCR扩增
1:阴性对照;M:DL2000 DNA marker;2:PtpA基因的PCR扩增产物
Figure 2. PCR amplification of PtpA gene in FLAG-PtpA recombinant plasmid
1: Negative control; M: DL2000 DNA marker; 2: PCR product of PtpA gene
![]()
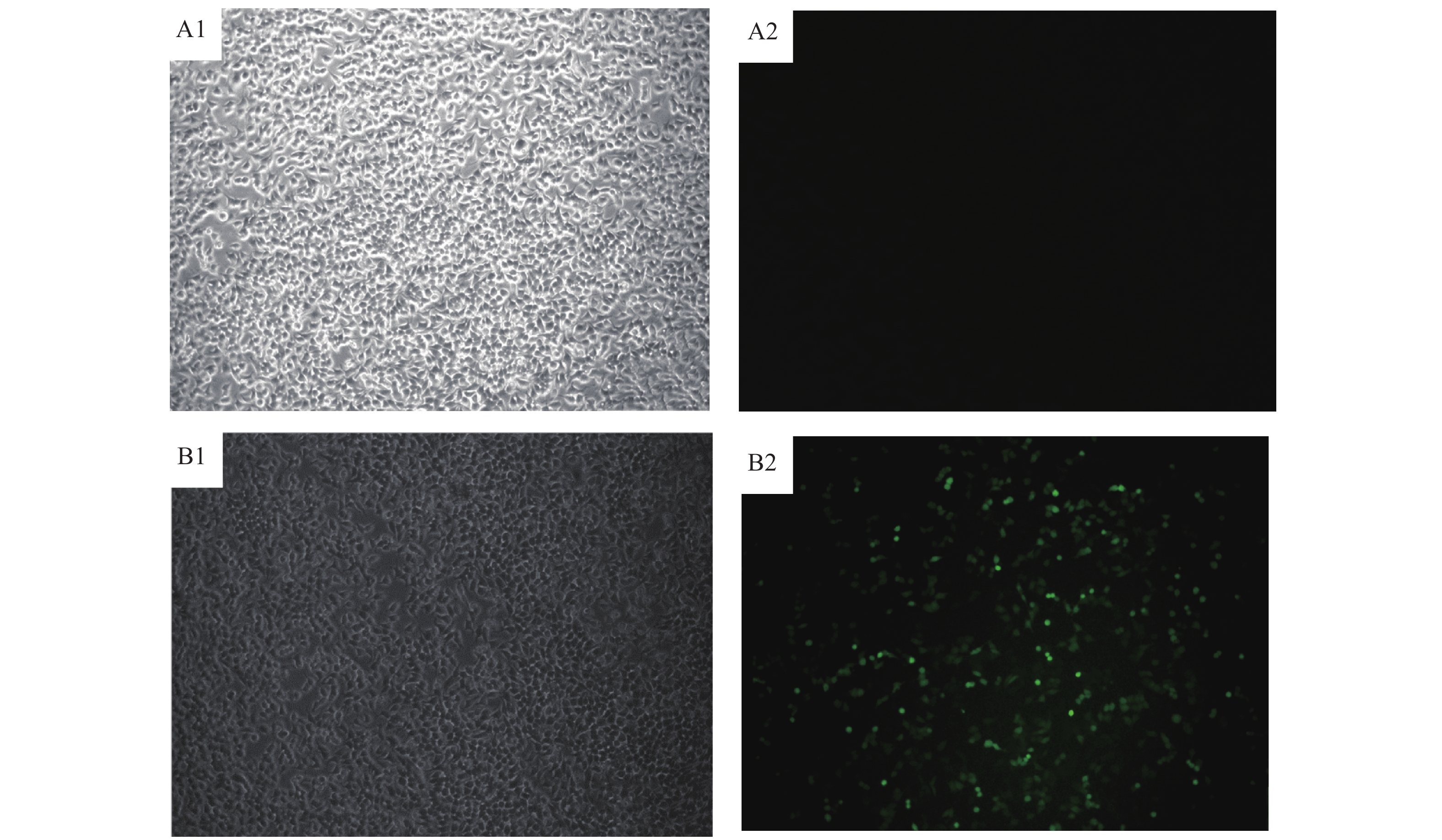
图 3 真核表达载体转染293T细胞
A1:293T未转染质粒(白镜);A2:293T未转染质粒(荧光);B1:293T转染FLAG-PtpA载体质粒+pEGFP-N1质粒(白镜);B2:293T转染FLAG-PtpA载体质粒+pEGFP-N1质粒(荧光)
Figure 3. Eukaryotic expression vector transfected 293T cells
A1: 293T untransfected plasmid (white mirror); A2: 293T untransfected plasmid (fluorescent); B1: 293T transfected FLAG-PtpA plasmid and pEGFP-N1 plasmid (white mirror); B2: 293T transfected FLAG-PtpA plasmid and pEGFP-N1 plasmid (fluorescence)
![]()
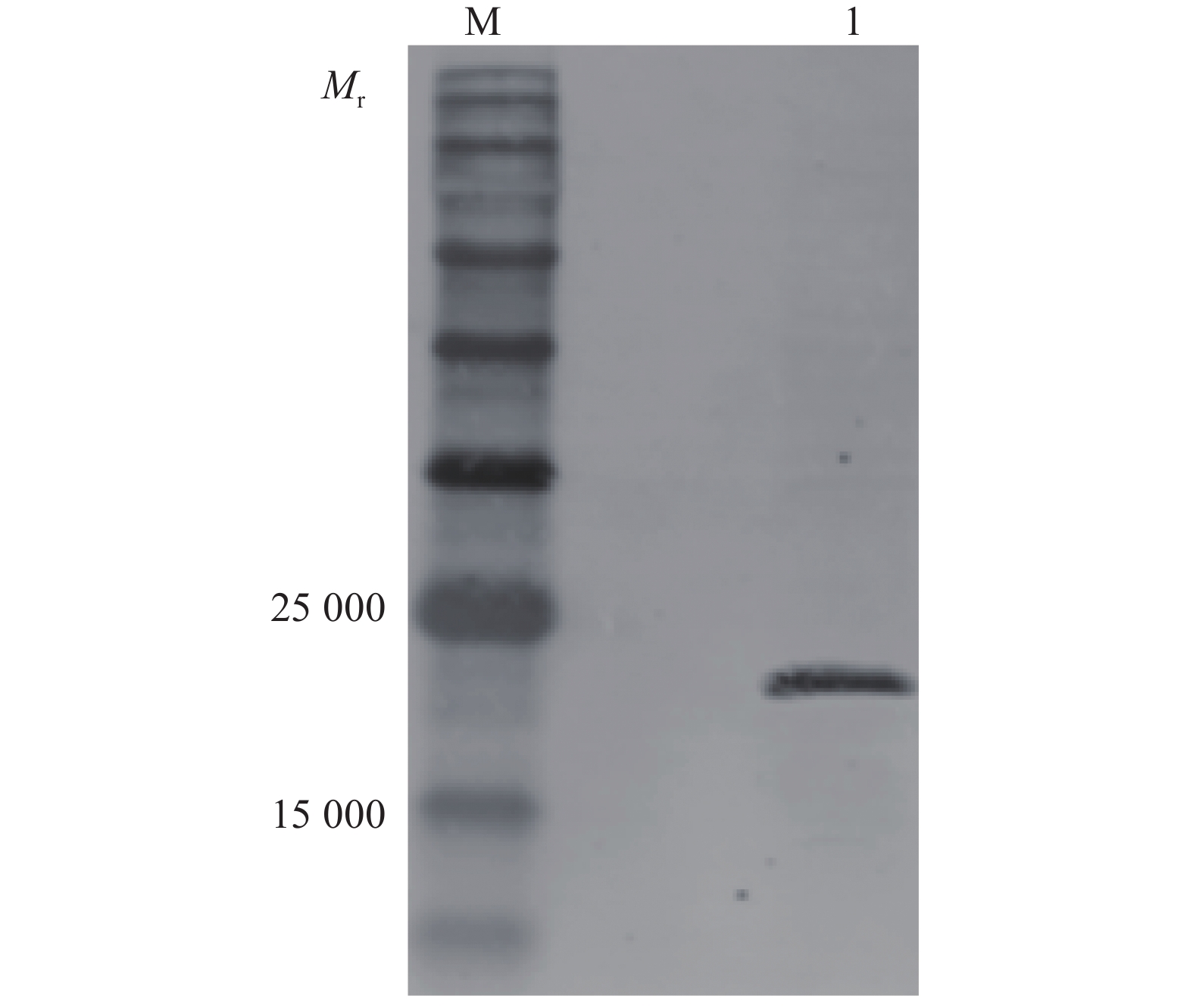
图 4 HEK293T细胞中FLAG-PtpA表达产物鉴定
M:低相对分子质量蛋白质标准;1:空白对照;2:HEK293T细胞中转染p3×FLAG-CMV-10质粒;3:HEK293T细胞中转染FLAG-PtpA质粒
Figure 4. Identification of FLAG-PtpA expression product in HEK293T cells
M: Low molecular weight protein standard; 1: Blank control; 2: p3×FLAG-CMV-10 plasmid transfected into HEK293T cells; 3: FLAG-PtpA plasmid transfected into HEK293T cells
![]()
图 5 HEK293T表达FLAG-PtpA蛋白特异性分析
M:低相对分子质量蛋白质标准;1:HEK293T表达的FLAG-PtpA蛋白
Figure 5. HEK293T expression FLAG-PtpA protein specificity analysis
M: Low molecular weight protein standard; 1: FLAG-PtpA protein expressed by HEK293T
![]()
图 6 PtpA蛋白对NF-κB信号通路激活的影响
“*”、“**”和“***”分别表示差异达到0.05、0.01和0.001的显著水平(t检验)
Figure 6. Effect of PtpA protein on activation of NF-κB signaling pathway
“*”, “**” and “***” indicate the difference reaches 0.05, 0.01 and 0.001 significance levels respectively (t test)
![]()
图 7 细胞因子的mRNA表达量
“*”、“**”和“***”分别表示差异达到0.05、0.01和 0.001的显著水平(t检验)
Figure 7. mRNA expression of cell factors
“*”,“**” and “***” indicate the difference reaches 0.05,0.01 and 0.001 significance levels respectively (t test)
表 1 引物名称及序列
Table 1 Primer name and sequence
引物名称
Primer name引物序列(5′→3′)1)
Primer sequence产物大小/bp
Product sizeFLAG-PtpA-F AAGGAAAAAA GCGGCCGCGGTGTCTGATCCGCTGCACG 492 FLAG-PtpA-R CTAG TCTAGATCAACTCGGTCCGTT IL-6-F GGTGTTGCCTGCTGCCTTCC 100 IL-6-R GTTCTGAAGAGGTGAGTGGCTGTC GM-CSF-F TCCTGAACCTGAGTAGAGACACTGC 100 GM-CSF-R CAGGTCGGCTCCTGGAGGTC BIRC-2-F AGACACATGCAGCTCGAATGAGAAC 100 BIRC-2-R AACACCTCAAGCCACCATCACAAC BIRC-3-F CTGTGATGGTGGACTCAGGTGTTG 100 BIRC-3-R TGGCTTGAACTTGACGGATGAACTC 1) 有下划线的序列为Not I、Xba I酶切位点
1) Sequences with underlines are Not I and Xba I restriction sites 下载: 导出CSV
下载: 导出CSV
-
[1] SMITH N H, GORDON S V, DE LA RUA-DOMENECH R, et al. Ecotypes of the Mycobacterium tuberculosis complex[J]. J Theor Biol, 2006, 239(2): 220-225. doi: 10.1016/j.jtbi.2005.08.036
[2] DELAHAY R J, DE LEEUW A N, BARLOW A M, et al. The status of Mycobacterium bovisinfection in UK wild mammals: A review[J]. Vet J, 2002, 164(2): 90-105. doi: 10.1053/tvjl.2001.0667
[3] MACIEL A, LOIKO M R, BUENO T S, et al. Tuberculosis in Southern Brazilian wild boars (Sus scrofa): First epidemiological findings[J]. Transbound Emerg Dis, 2018, 65(2): 518-526. doi: 10.1111/tbed.2018.65.issue-2
[4] PHILLIPS C J C, FOSTER C R W, MORRIS P A, et al. The transmission of Mycobacterium bovis infection to cattle[J]. Res Vet Sci, 2003, 74(2): 1-15.
[5] PUCKEN V B, KNUBBEN-SCHWEIZER G, DOPFER D, et al. Evaluating diagnostic tests for bovine tuberculosis in the southern part of Germany: A latent class analysis[J]. PLoS One, 2017, 12(6): e0179847. doi: 10.1371/journal.pone.0179847
[6] DE KANTOR I N and RITACCO V. An update on bovine tuberculosis programmes in Latin American and Caribbean countries[J]. Vet Microbiol, 2006, 112(2/3/4): 111-118. doi: 10.1016/j.vetmic.2005.11.033
[7] STONE M J, BROWN T J and DROBNIEWSKI F A. Human Mycobacterium bovis infections in London and Southeast England[J]. J Clin Microbiol, 2012, 50(1): 164-165. doi: 10.1128/JCM.05692-11
[8] COWLEY S C, BABAKAIFF R and AV-GAY Y. Expression and localization of the Mycobacterium tuberculosis protein tyrosine phosphatase PtpA[J]. Res Microbiol, 2002, 153(4): 233-241. doi: 10.1016/S0923-2508(02)01309-8
[9] CHAO Y, XING Y, CHEN Y, et al. Structure and mechanism of the phosphotyrosyl phosphatase activator[J]. Mol Cell, 2006, 23(4): 535-546. doi: 10.1016/j.molcel.2006.07.027
[10] WANG J, LI B X, GE P P, et al. Mycobacterium tuberculosis suppresses innate immunity by coopting the host ubiquitin system[J]. Nat Immunol, 2015, 16(3): 237-245. doi: 10.1038/ni.3096
[11] PFAFFI M W. A new mathematical model for relative quantification in real-time RT-PCR[J]. Nucleic Acids Res, 2001, 29(9): e45. doi: 10.1093/nar/29.9.e45
[12] TONKS N K. Protein tyrosine phosphatases: From genes, to function, to disease[J]. Nat Rev Mol Cell Biol, 2006, 11(7): 833-846.
[13] FU Y and GALAN J E. The Salmonella typhimurium tyrosine phosphatase SptP is translocated into host cells and disrupts the actin cytoskeleton[J]. Mol Microbiol, 1998, 27(2): 359-368. doi: 10.1046/j.1365-2958.1998.00684.x
[14] GOEBEL-GOODY S M, WILSON-WALLIS E D, ROYSTON S, et al. Genetic manipulation of STEP reverses behavioral abnormalities in a fragile X syndrome mouse model[J]. Genes Brain Behav, 2012, 11(5): 586-600. doi: 10.1111/j.1601-183X.2012.00781.x
[15] SHI L, POTTS M and KENNELLY P J. The serine, threonine, and/or tyrosine-specific protein kinases and protein phosphatases of prokaryotic organisms: A family portrait[J]. FEMS Microbiol Rev, 1998, 22(4): 229-253. doi: 10.1111/j.1574-6976.1998.tb00369.x
[16] YONGFANG Z, KURUP P, JIAN X, et al. Genetic reduction of striatal-enriched tyrosine phosphatase (STEP) reverses cognitive and cellular deficits in an Alzheimer's disease mouse model[J]. Proc Natl Acad Sci USA, 2010, 107(44): 19014-19019. doi: 10.1073/pnas.1013543107
[17] WONG D, BACH H, SUN J, et al. Mycobacterium tuberculosis protein tyrosine phosphatase (PtpA) excludes host vacuolar-H+-ATPase to inhibit phagosome acidification[J]. Proc Natl Acad Sci USA, 2011, 108(48): 19371-19376. doi: 10.1073/pnas.1109201108
[18] WANG J, GE P, QIANG L, et al. The mycobacterial phosphatase PtpA regulates the expression of host genes and promotes cell proliferation[J]. Nat Commun, 2017, 8(1): 244. doi: 10.1038/s41467-017-00279-z
[19] 孟露萍, 史梦婷, 包海洋, 等. 结核分枝杆菌Rv2626c蛋白对RAW264.7细胞凋亡的影响[J]. 中国畜牧兽医, 2016, 43(4): 892-898. [20] VOLKMAN H E, POZOS T C, ZHENG J, et al. Tuberculous granuloma induction via interaction of a bacterial secreted protein with host epithelium[J]. Science, 2010, 327(5964): 466-469. doi: 10.1126/science.1179663
-
期刊类型引用(0)
其他类型引用(4)



计量
- 文章访问数: 1393
- HTML全文浏览量: 11
- PDF下载量: 1365
- 被引次数: 4
