Fermentation optimization of Penicillium citrinum PA-33 strain by response surface method
-
摘要:目的
优化桔青霉Penicillium citrinum PA-33发酵培养基和发酵条件,以提高桔青霉PA-33的抗菌活性。
方法采用单因素试验确定桔青霉PA-33发酵所需最适基础培养基、碳氮源和无机盐,并利用响应面法设计确定最适发酵培养基配方;在发酵条件单因素试验基础上,采用三元二次通用旋转组合设计和频率分析法优化其最适发酵条件组合。
结果经优化后,最佳发酵培养基配方为马铃薯汁液219.91 g·L−1、甘露醇34.11 g·L−1、黄豆粉6.25 g·L−1;最适发酵条件为装液量50 mL、接种量3.5% (φ)、发酵温度28 ℃,摇床转速150 r·min−1、发酵时间12 d。优化后发酵液对大肠埃希菌Escherichia coli的抑菌圈直径为28.99 mm,较优化前抑菌圈直径 (18.73 mm) 增加了10.26 mm。
结论采用响应面法、三元二次通用旋转组合设计和频率分析法优化发酵工艺,显著提高了桔青霉PA-33发酵液的抗菌活性,为该菌株的抗菌活性物质的分离以及工业化生产提供依据。
Abstract:ObjectiveTo optimize the medium composition and fermentation conditions of Penicillium citrinum PA-33 to improve its antibacterial activity.
MethodSingle factor experiments were used to determine the optimum basic medium, carbon sources, nitrogen sources and inorganic salts. The optimum formulation of fermentation medium was determined by response surface methodology. On the basis of single factor tests of fermentation conditions, the optimal combination of fermentation conditions was optimized by ternary quadratic rotation unitized design and frequency analysis method.
ResultThe optimal medium composition were: Potato juice 219.91 g·L−1, mannitol 34.11 g·L−1 and soybean powder 6.25 g·L−1. The optimal fermentation conditions were: Liquid volume 50 mL, inoculation concentration 3.5% (φ), culture temperature 28 ℃, rotation speed 150 r·min−1 and fermentation for 12 d. The inhibition zone diameter of fermentation broth after optimization on Escherichia coli reached 28.99 mm, increased by 10.26 mm compared with 18.73 mm under the original conditions.
ConclusionResponse surface methodology, ternary quadratic rotation unitized design and frequency analysis method significantly enhance the antibacterial activity of fermentation broth of Penicillium citrinum PA-33, and this study provides a basis for isolation of antibacterial active substances and industrial production of this strain.
-
荞麦(别名:甜荞)Fagopyrum esculentum Moench是蓼科Polygonaceae荞麦属Fagopyrum作物[1]。研究表明,荞麦是一种重要的食药兼用杂粮作物,蛋白质和维生素B1、B2的含量较高[1]。荞麦干粉中含有约65 mg·kg-1的芦丁,芦丁能够影响胰岛细胞的功能,促进胰岛素的分泌,从而降低血糖;芦丁还可以调节血压,强化血管,对脑溢血、网膜出血等有一定的预防、治疗作用[2]。此外,荞麦籽粒中所含有的可溶性膳食纤维约占膳食纤维总量的20%~30%,可溶性膳食纤维具有降低血清总胆固醇及LDL胆固醇含量的作用[3]。甜荞是荞麦属植物主要的栽培品种之一,但产量较低,影响了农户对甜荞的种植热情,严重制约了甜荞产业的发展。
源库理论是改良作物株型,提高作物产量的重要理论基础。调控作物源库器官是研究作物源库关系以及产量形成的有效途径[4-6]。郝满等[7]对玉米进行源库调节的研究发现,源对玉米的生长发育、干物质积累、产量形成有重要影响。周龙祥等[8]研究发现,随着疏花数量增多,稻米籽粒中的直链淀粉含量相应增加。陈若礼等[9]在研究源与库对小麦产量高低关系时得出,源和库共同制约着小麦产量高低。目前在水稻、玉米和小麦等作物中有关源-库关系的研究报道较多,其调控机制也较为清楚。甜荞具有无限花序特性,源库关系较为复杂,相关研究鲜见报道,其调控机理也不清楚。探索甜荞的源库关系,阐明其源库调控机理,对于荞麦育种与高产栽培技术研究具有重要意义。本试验以高产甜荞品种丰甜1号为材料,在灌浆期进行减库(剪花)处理,研究减库处理对丰甜1号农艺性状以及产量的影响,并使用Richards方程模拟减库处理后籽粒灌浆特性,探索甜荞源库关系,为甜荞的高产栽培提供理论依据。
1. 材料与方法
1.1 材料与试验设计
甜荞品种为丰甜1号。2015年3月28日播种于贵州省贵阳市永乐乡柏杨村试验基地(海拔908 m)。随机区组设计,开花期选取长势一致、无病虫害的植株进行标记。于开花后7 d剪去未开放的花朵及花序,作为减库处理;以未进行减库处理的植株为对照。试验小区面积为10 m2(2 m×5 m),3次重复。行距为33 cm,种植密度为120万株·hm-2。每小区施肥量分别为尿素[w(N)46%]225 g、过磷酸钙[w(P2O5)14%]246.43 g、氯化钾[w(K2O) 60%]4.25 g,3种肥料混匀后作为底肥一次性施入。生育期间采用常规大田管理,无明显病虫害发生。
1.2 样品制备
减库后第7天至籽粒成熟,每7 d取样1次,每小区选取10~15株生长一致的植株进行取样。将植株第3至第4节节间叶片迅速剪下,置于液氮中冷冻30 s后,保存于-80 ℃冰箱,用于叶绿素、可溶性糖和可溶性蛋白的含量测定。将植株籽粒收集于纸袋中,置于105 ℃烘箱中杀青10 min后,于60 ℃烘干至恒质量,将籽粒脱壳、粉碎、过100目筛,用于籽粒淀粉含量测定。
1.3 指标测定
1.3.1 农艺性状
成熟期随机选取10株进行株高、主茎分枝数、主茎节数、单株粒数、单株粒质量测定;按小区进行收获,测定小区产量。
1.3.2 代谢物质含量的测定
叶绿素含量采用丙酮研磨提取法进行测定[10]:称取0.5 g样品,使用乙醇丙酮混合液提取,于暗处保存至材料变为白色,然后在645和663 nm下比色,计算叶绿素含量。可溶性糖含量采用蒽酮比色法进行测定[10]:称取0.2 g样品,使用φ为80%乙醇溶液在80 ℃进行提取,经活性炭脱色过滤,加入蒽酮试剂后在620 nm下比色,根据葡萄糖标准曲线计算样品可溶性糖含量。可溶性蛋白含量采用考马斯亮蓝法进行测定[10]:称取0.1 g样品,使用磷酸缓冲液进行提取,加入考马斯亮蓝溶液后在595 nm下比色,根据牛血清蛋白标准曲线计算样品可溶性蛋白含量。总淀粉含量采用蒽酮硫酸比色法进行测定[11]:称取40 mg样品,使用φ为80%乙醇溶液在80 ℃提取,除去可溶性糖,使用高氯酸进行酸化提取,加入蒽酮试剂后在620 nm下比色,根据葡萄糖标准曲线计算样品可溶性糖含量。籽粒直链淀粉含量采用碘蓝比色法进行测定[12]:称取100 mg样品,经φ为95%乙醇溶液和1 mol·L-1 NaOH溶液脱脂、煮沸、冷却后,加入碘-乙酸溶液后在720 nm下比色,根据马铃薯淀粉标准曲线计算样品直链淀粉含量。支链淀粉含量=总淀粉含量-直链淀粉含量。
1.4 减库后籽粒灌浆进程拟合
参考朱庆森等[13]和顾世梁等[14]的方法,使用Richards方程对丰甜1号籽粒进行灌浆生长拟合。以剪花后天数(t,剪花当日为0) 为自变量,以各时期荞麦籽粒百粒质量(m)为应变量,Richards方程拟合的公式为:
$$ m=A/{{\left( 1+B{{\rm{e}}^{-Kt}} \right)}^{1/N}} $$ 式中,A为生长终值量,B为初值参数,K为生长速率参数,N为形状参数。
以决定系数R2(m依t的回归平方和占总平方和的比率)表示配合度。Richards方程生长曲线是由N值大小决定的一簇曲线,当0<N<1时,灌浆速率达到最大的位置(即曲线拐点)在0.367~0.5A之间,灌浆速率曲线左偏;当N=1时,即为Logistic方程,曲线拐点在0.5A处;当N>1时,曲线拐点大于0.5A,并随N的增大而逐渐接近于A,灌浆速率曲线右偏[13-14]。
1.5 数据分析
采用Excel 2003、SPSS 17.0对数据进行相关统计分析。
2. 结果与分析
2.1 减库处理对丰甜1号叶片生理特性的影响
由表 1可知,丰甜1号叶片的叶绿素含量在减库后21 d前减库处理与对照差异不显著; 随着灌浆进程,减库后28 d叶绿素含量均很低, 但减库处理叶片叶绿素含量显著高于对照。减库后7和21 d,其叶片叶绿素a/b值均显著高于对照,其余时期差异不显著。减库处理下灌浆期叶片可溶性糖含量变化幅度较小,且减库处理与对照均无显著差异,说明减库处理对叶片可溶性糖的合成与积累影响较小。减库处理后14 d叶片可溶性蛋白含量与对照差异不显著,但在7、21和28 d均显著高于对照。
表 1 减库处理后丰甜1号叶片代谢物质含量变化1)Table 1. Changes of metabolite contents in leaves of Fengtian 1 after sink-limiting treatment处理 w(叶绿素)/ (mg·g-1) 叶绿素a/b w(可溶性糖)/% w(可溶性蛋白)/ (mg·g-1) 7 d 14 d 21 d 28 d 7 d 14 d 21 d 28 d 7 d 14 d 21 d 28 d 7 d 14 d 21 d 28 d 减库 1.77a 1.78a 0.65a 0.10a 4.19a 4.06a 3.53a 2.84a 5.65a 5.61a 6.16a 5.56a 4.35a 6.46a 6.58a 6.02a 对照 1.79a 1.84a 0.62a 0.05b 4.02b 4.00a 3.44b 2.89a 5.64a 5.76a 6.12a 5.60a 3.63b 6.54a 6.35b 5.81b 1) 同列数据后凡是有一个相同小写字母者,表示差异不显著(P>0.05,Duncan’s法)。 2.2 减库处理对丰甜1号籽粒灌浆过程的影响
由表 2可见,减库处理和对照曲线方程的决定系数(R2)分别为0.996和0.992,拟合度较好,说明Richards方程适用于甜荞灌浆进程的模拟。减库处理和对照的N均小于1,灌浆速率曲线左偏;减库处理和对照的A都比实际值大,且减库处理的A小于对照。减库处理灌浆起始势(R0)、最大灌浆速率(Gmax)以及灌浆速率最大时的生长量与籽粒生长终值量比值(I)均大于对照;减库处理最大灌浆速率的时间(Tmax)、达到最大灌浆速率时的生长量(mmax)和灌浆活跃生长期(D)均小于对照。
表 2 减库处理下丰甜1号籽粒灌浆Richards方程参数Table 2. Richards equation parameters of grain filling of Fengtian 1 after sink-limiting treatment处理 A B K N R2 R0 Tmax/d Gmax1)/(g·d-1) mmax/g I/% D/d 减库 3.67 2.283 0.116 0.340 0.996 1.012 16.37 0.135 1.551 42.28 40.21 对照 4.92 0.276 0.069 0.068 0.992 0.342 20.51 0.120 1.870 38.00 50.13 1) 按100粒计。 丰甜1号灌浆前期和后期的灌浆持续时间(t)减库处理大于对照,中期则小于对照;灌浆前期和后期的灌浆平均速率减库处理小于对照,中期则大于对照;灌浆期减库处理和对照对粒质量的贡献率均表现为中期>后期>前期(表 3)。
表 3 籽粒灌浆阶段的划分1)Table 3. The division of grain filling stages处理 前期 中期 后期 t/d 速率/(g·d-1) 贡献率/% t/d 速率/(g·d-1) 贡献率/% t/d 速率/(g·d-1) 贡献率/% 减库 6.91 0.067 12.59 18.92 0.117 60.38 30.06 0.032 26.02 对照 6.02 0.069 8.40 28.97 0.104 60.94 17.14 0.057 19.87 1)t:灌浆持续时间;速率:灌浆平均速率,按100粒计;贡献率:对粒质量的贡献率。 2.3 减库处理对丰甜1号籽粒淀粉积累的影响
由图 1可知,丰甜1号籽粒中直链淀粉、支链淀粉及总淀粉积累量都随着灌浆进程逐渐增多; 灌浆中前期籽粒淀粉含量以减库处理高于对照,并在减库后21 d达到显著差异; 灌浆后期,减库处理的籽粒淀粉含量低于对照。支链淀粉和总淀粉含量的变化趋势相似,在灌浆中前期,减库处理的支链淀粉和总淀粉含量均高于对照,在灌浆后期均低于对照,且在减库后28 d处理和对照差异显著。在灌浆中后期,减库处理的直链淀粉含量明显高于对照,并在减库后21和28 d均达到显著差异。
![图 1 减库处理下灌浆期丰甜1号淀粉的积累]() 图 1 减库处理下灌浆期丰甜1号淀粉的积累*表示相同时间处理与对照差异显著(P<0.05, t检验)。Figure 1. Changes of starch contents in Fengtian 1 during grain filling after sink-limiting treatment
图 1 减库处理下灌浆期丰甜1号淀粉的积累*表示相同时间处理与对照差异显著(P<0.05, t检验)。Figure 1. Changes of starch contents in Fengtian 1 during grain filling after sink-limiting treatment2.4 减库处理对丰甜1号主要农艺性状的影响
由表 4可知,减库处理丰甜1号植株株高、主茎分枝数、主茎节数与对照差异不显著,说明减库处理并不影响植株的正常生长。同样,减库处理下,丰甜1号的单株粒数及单株粒质量与对照差异不显著,但显著降低了丰甜1号的产量,较对照减产5.59%。
表 4 减库处理下丰甜1号的主要农艺性状1)Table 4. The major agronomic characters of Fengtian 1 after sink-limiting treatment处理 株高/cm 主茎分枝数 主茎节数 单株粒数 单株粒质量/g 产量/ (kg·hm-2) 减库 56.9a 3.3a 6.8a 49.3a 1.68a 1 192.4b 对照 57.6a 3.3a 7.0a 45.0a 1.75a 1 263.0a 1) 同列数据后凡是有一个相同小写字母者,表示差异不显著(P>0.05,Duncan’s法)。 3. 讨论与结论
王楠等[15]研究认为作物产量的形成与库源变化关系密切。灌浆物质的充足与否直接影响产量的高低[16]。前人研究表明,叶绿素含量的多少是光合产物合成能力强弱的间接表现[17-19],可溶性蛋白和可溶性糖是籽粒灌浆物质的直接氮源和碳源[20]。库容量直接决定作物的产量潜力。减库是研究作物源库关系的重要途径[21]。从本研究结果看出,减库处理下叶片叶绿素和可溶性糖含量与对照差异不显著,同时,减库处理下植株农艺性状与对照也无显著差异,说明减库处理并未对源器官造成明显影响。但是,减库后甜荞产量较对照显著降低,这与钏兴宽等[22]在水稻上和陈若礼等[9]在小麦上的研究结果一致。钏兴宽等[22]研究认为,两系杂交粳稻和常规粳稻在强源的同时应适当扩库,以提高产量水平,库容量下降不利于产量形成。甜荞具有无限花序特性,库容量较为充足,但是其产量水平一直难以得到突破性的提高。袁继超等[23]研究认为源不足会降低光合产物的合成与积累,进而影响产量。本研究发现,减库处理后甜荞的单株粒数并未出现下降,单株粒质量与对照也无显著差异,说明开花7 d后甜荞新生花朵的结实率极低,并不能形成饱满种子,这可能与甜荞营养生长期和生殖生长期重叠时间较长,光合产物不足以及光合产物分配不均有关。
灌浆期是作物产量形成的关键时期,Richards方程生长曲线是广泛应用于模拟作物灌浆进程的重要工具[24-25]。用Richards生长曲线对减库处理后丰甜1号灌浆过程进行拟合,拟合度(R2)为0.996,说明用Richards生长曲线能够拟合减库处理后丰甜1号籽粒灌浆的连续过程。从本研究结果可以看出,减库处理和对照籽粒灌浆的N均小于1,生长曲线左偏,说明其灌浆物质相对充分,表现为灌浆前期生长迅速,随后逐渐减弱。籽粒灌浆起始势(R0)反映了其子房的生长潜势,R0值大,则胚乳细胞分裂周期短,分裂快,籽粒灌浆启动早,说明开花后较短时间内达到最大灌浆速率。从产量高低来看,减库处理的产量较低,其R0值较大,在较短的时间内达到最大灌浆速率,但灌浆持续时间较短;对照的产量较高,其R0值较小,需要较长时间才达到最大灌浆速率,灌浆持续时间较长。减库后籽粒中总淀粉的积累量在灌浆中、前期高于对照,在21 d时达到显著差异,但在灌浆后期籽粒淀粉积累量低于对照。减库处理可以避免光合产物向新生花朵输入,有利于前期种子的灌浆,但随着灌浆进程,当种子达到最大灌浆速率时,生长量(mmax)变低和总灌浆天数(D)缩短导致总积累量降低,成熟期单株粒质量与对照无显著差异,但产量较对照显著下降。甜荞源不足或碳水化合物分配不均导致的种子空瘪率高是限制甜荞产量提高的重要因素。因此,调控光合产物合成及其有效分配是提高甜荞产量的有效途径。
-
![]()
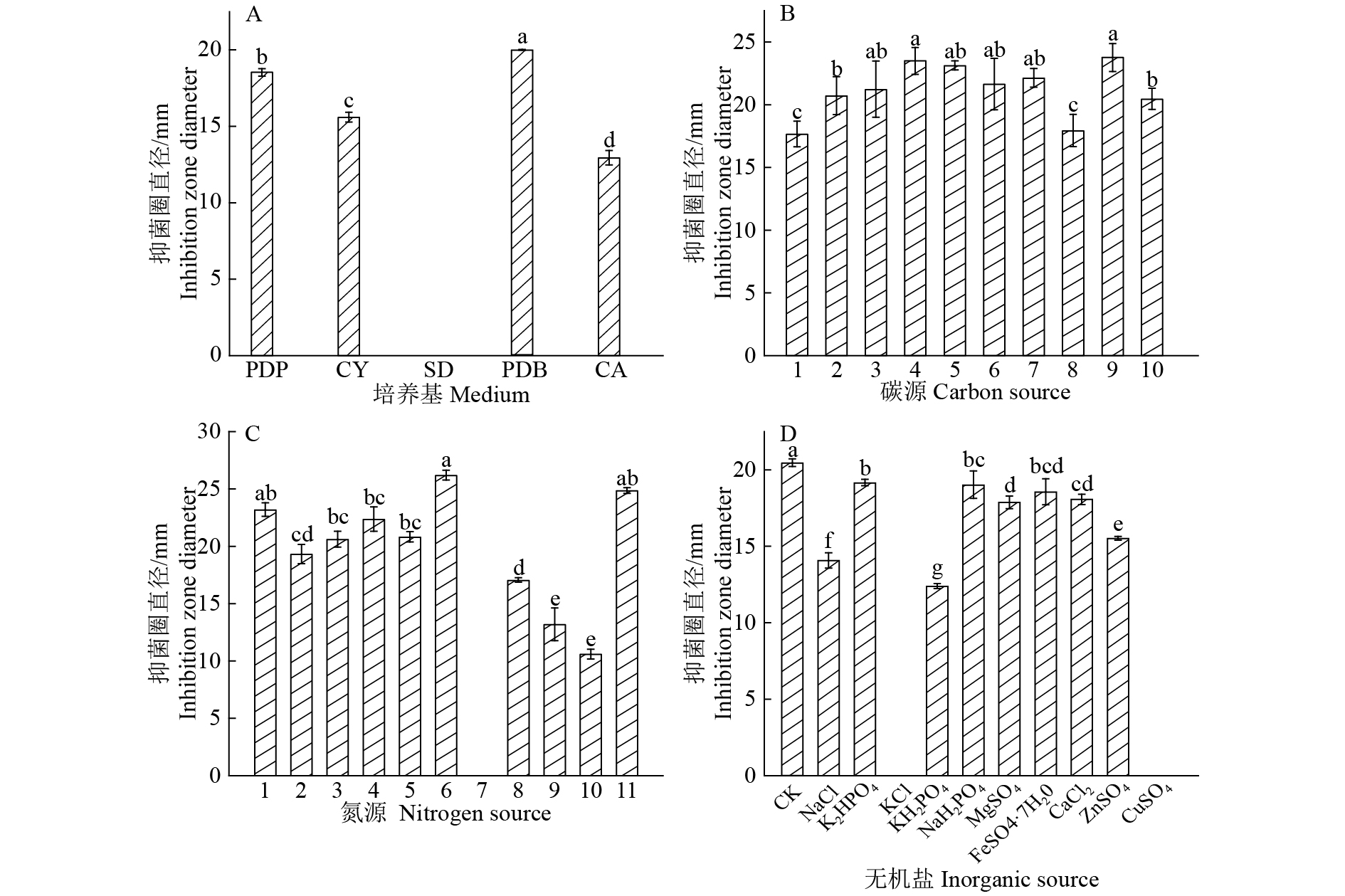
图 1 不同培养基成分对发酵液抗菌活性的影响
B图中横坐标的数字1、2、3、4、5、6、7、8、9和10分别代表葡萄糖、蔗糖、淀粉、玉米粉、乳糖、果糖、糊精、甘油、甘露醇和麦芽糖;C图中横坐标的数字1、2、3、4、5、6、7、8、9、10和11分别代表对照、蛋白胨、酵母浸粉、牛肉膏、胰蛋白胨、黄豆粉、尿素、KNO3、NH4NO3、NH4Cl和陈皮;各图中柱子上方的不同小写字母表示差异显著(P<0.05, Duncan’s法)
Figure 1. Effects of different medium ingredients on the antibacterial activities of fermentation broth
In figure B,number 1,2,3,4,5,6,7,8,9 and 10 represent glucose, sucrose, starch, corn flour, lactose, fructose, dextrin, glycerin, mannitol and maltose, respectively; In figure B,number 1,2,3,4,5,6,7,8,9, 10 and 11 represent control, peptone, yeast extract, beef extract, tryptone, soybean powder, urea, KNO3, NH4NO3, NH4Cl and bran, respectively; In each figure, different lowercase letters on the bars indicate significant difference (P<0.05, Duncan’s test)
![]()
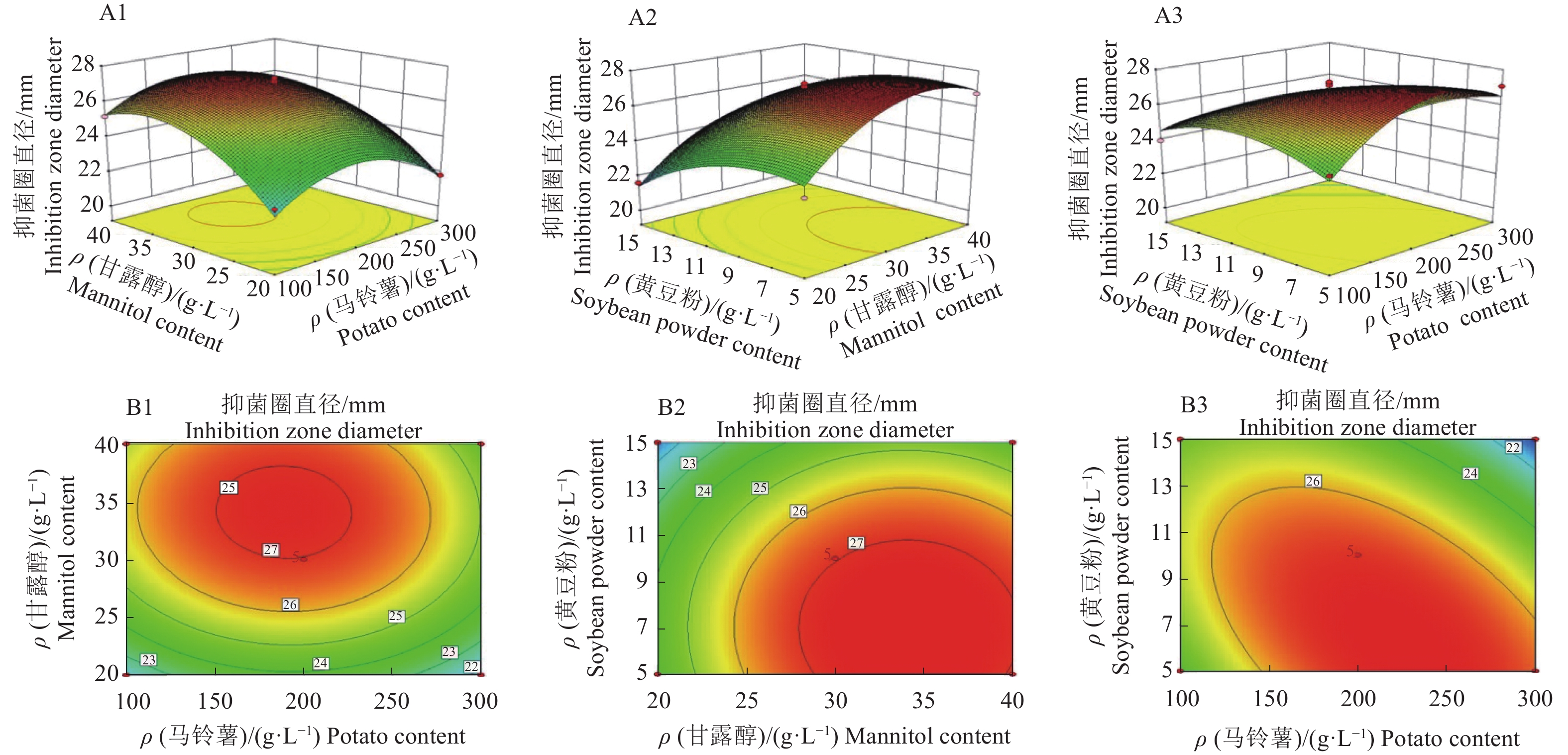
图 2 菌株PA-33抗菌活性的响应曲面图
A1、A2和A3为响应面3D图;B1、B2和B3为等高线图
Figure 2. Response surface plots of antibacterial activity of strain PA-33
A1, A2, and A3: 3D maps of response surfaces; B1, B2, and B3: Contour maps
![]()
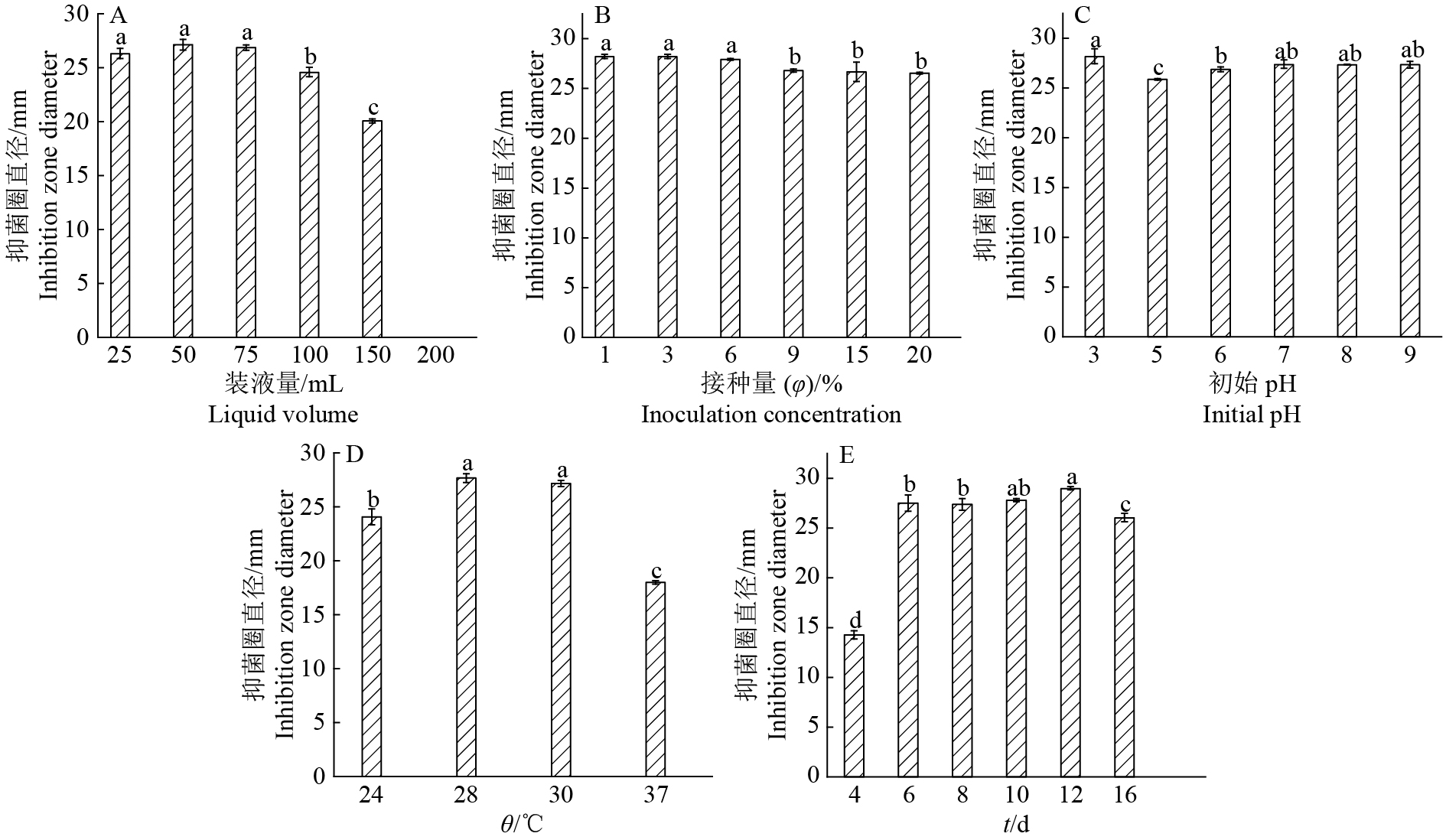
图 3 不同发酵条件对菌株PA-33抑菌活性的影响
各图中, 柱子上方的不同小写字母表示差异显著(P<0.05, Duncan’s法)
Figure 3. Effect of different dermentation condition on antimicrobial activity of strain PA-33
In each figure, different lowercase letters on the bars indicate significant difference (P<0.05, Duncan’s test)
表 1 不同质量浓度的培养基成分对菌株PA-33抗菌活性的影响1)
Table 1 Effects of different concentrations of medium components on antibacterial activities of fermentation broth
ρ(马铃薯)/(g·L−1) Patato content 抑菌圈直径/mm Inhibition zone diameter ρ(甘露醇)/(g·L−1) Mannitol content 抑菌圈直径/mm Inhibition zone diameter ρ(黄豆粉)/((g·L−1) Soybean powder content 抑菌圈直径/mm Inhibition zone diameter 100 22.93±0.52ab 20 22.03±0.12c 5.0 23.83±0.69c 150 23.13±0.25ab 25 23.33±0.09b 7.5 27.93±0.19a 200 23.47±0.25a 30 26.60±0.83a 10.0 25.77±0.31b 250 22.53±0.19b 35 25.50±0.08a 12.5 25.13±0.09b 300 21.80±0.16c 40 23.43±0.87b 15.0 22.43±0.17d 1) 同列数据后的不同小写字母表示差异显著 (P<0.05,Duncan’s 法 )
1) Different lowercase letters in the same column indicate significant difference (P<0.05,Duncan’s test) 下载: 导出CSV
下载: 导出CSV
表 2 Box-Behnken试验方差分析
Table 2 Analysis of variance for Box-Behnken experiment
来源1) Source SS DF MS F P2) 模型 Model 76.25 9 8.47 22.24 0.000 2** A 1.07 1 1.07 2.80 0.138 3 B 15.62 1 15.62 41.01 0.000 4** C 14.36 1 14.36 37.70 0.000 5** AB 0.07 1 0.07 0.17 0.691 9 AC 9.52 1 9.52 24.98 0.001 6** BC 0.01 1 0.01 0.02 0.894 4 A2 14.56 1 14.56 38.22 0.000 5** B2 12.31 1 12.31 32.30 0.000 7** C2 5.19 1 5.19 13.61 0.007 8** 残差
Residual2.67 7 0.38 失拟
Lack of fit1.96 3 0.65 3.71 0.118 9 误差
Error0.71 4 0.18 总变异
Total variation78.92 16 1) A、B 和 C 分别表示马铃薯、甘露醇和黄豆粉;2) “**” 表示模型具有极显著影响 (P<0.01)
1) A, B and C represent potato, mannitol and soybean powder, respectively; 2) “**” indicates the model is highly significant(P<0.01)
下载: 导出CSV
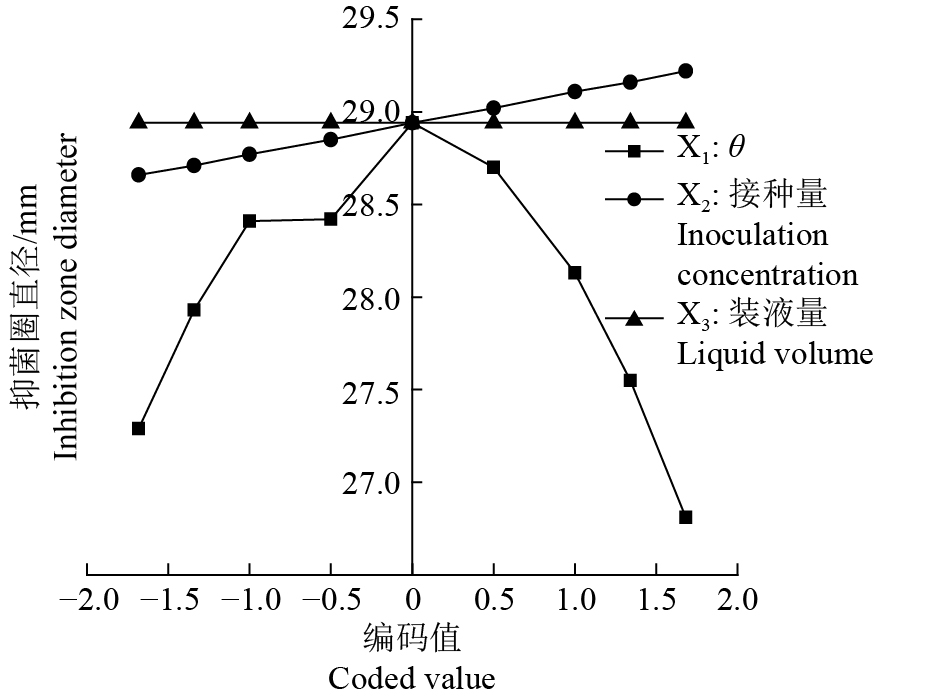
表 3 三元二次通用旋转组合试验设计及结果1)
Table 3 Design and results for quadratic general rotary composite
试验号
Test No.θ/℃ 接种量(φ)/ %
Inoculation concentration装液量/mL
Liquid volume抑菌圈直径/mm
Inhibition zone diameter1 30(1) 5(1) 60(1) 28.30 2 30(1) 5(1) 40(−1) 28.37 3 30(1) 2(−1) 60(1) 27.80 4 30(1) 2(−1) 40(−1) 27.93 5 26(−1) 5(1) 60(1) 28.73 6 26(−1) 5(1) 40(−1) 28.87 7 26(−1) 2(−1) 60(1) 28.40 8 26(−1) 2(−1) 40(−1) 28.57 9 24.6(−1.682) 3.5(0) 50(0) 26.88 10 31.4(1.682) 3.5(0) 50(0) 27.00 11 28(0) 0.977(−1.682) 50(0) 28.80 12 28(0) 6(1.682) 50(0) 29.23 13 28(0) 3.5(0) 33.2(−1.682) 28.53 14 28(0) 3.5(0) 66.8(1.682) 28.87 15 28(0) 3.5(0) 50(0) 28.80 16 28(0) 3.5(0) 50(0) 28.93 17 28(0) 3.5(0) 50(0) 29.20 18 28(0) 3.5(0) 50(0) 29.13 17 28(0) 3.5(0) 50(0) 29.00 18 28(0) 3.5(0) 50(0) 28.90 1) 括号中的数值为编码值
1) Numbers in the brackets are coded values
下载: 导出CSV
表 4 三元二次通用旋转组合试验结果方差分析
Table 4 Analysis of variance for results from quadratic general rotary united design
来源 Source SS DF MS PCC F P X1 0.283 6 1 0.283 6 −0.515 4 3.617 2 0.086 4 X2 0.385 1 1 0.385 1 0.573 9 4.910 4 0.051 0 X3 0.000 3 1 0.000 3 0.018 9 0.003 6 0.953 5 X12 6.406 3 1 6.406 3 −0.943 9 81.696 5 0.000 1 X22 0.064 5 1 0.064 5 0.275 6 0.822 2 0.385 9 X32 0.028 5 1 0.028 5 −0.187 3 0.363 6 0.559 9 X1X2 0.012 0 1 0.012 0 0.122 8 0.153 2 0.703 7 X1X3 0.001 5 1 0.001 5 0.043 9 0.019 3 0.892 3 X2X3 0.001 0 1 0.001 0 0.035 9 0.012 9 0.911 8 回归 Regression 7.349 1 9 0.816 6 10.413 2 0.000 8 残差 Residual 0.784 2 10 0.078 4 失拟 Lack of fit 0.541 6 5 0.108 3 2.233 2 0.130 9 误差 Error 0.242 5 5 0.048 5 总和 Total 8.133 2 19 1) X1、X2 和 X3 分别表示温度、接种量和装液量;PCC 为偏相关系数
1) X1,X2 andX3 represent temperature,inoculation concentration and liquid volume, respectively; PCC is patial correlation coefficient
下载: 导出CSV
表 5 各变量取值频率分布
Table 5 Frequency distribution of variable value
水平
Levelθ 接种量 Inoculation concentration 装液量 Liquid volume 次数 Times 频率 Frequency 次数 Times 频率 Frequency 次数 Times 频率 Frequency −1.682 0 0.000 5 0.143 7 0.200 −1.000 10 0.286 5 0.143 7 0.200 0.000 25 0.714 5 0.143 7 0.200 1.000 0 0.000 10 0.286 7 0.200 1.682 0 0.000 10 0.286 7 0.200
下载: 导出CSV
-
[1] 罗清, 彭程, 叶波平. 青霉属真菌研究新进展[J]. 药物生物技术, 2016, 5(23): 452-456. [2] 郑婕施, 江北, 张琪, 等. 青霉属真菌活性代谢产物研究进展[J]. 工业微生物, 2017(06): 50-56. doi: 10.3969/j.issn.1001-6678.2017.06.009 [3] LU Z Y, LIN Z J, WANG W L, et al. Citrinin dimers from the halotolerant fungus Penicillium citrinum B-57[J]. J Nat Prod, 2008, 71(4): 543-546. doi: 10.1021/np0704708
[4] DU L, LI D, ZHANG G, et al. Novel carbon-bridged citrinin dimers from a volcano ash-derived fungus Penicillium citrinum and their cytotoxic and cell cycle arrest activities[J]. Tetrahedron, 2010, 66(47): 9286-9290. doi: 10.1016/j.tet.2010.09.036
[5] CLARK B R, CAPON R J, LACEY E, et al. Citrinin revisited: From monomers to dimers and beyond[J]. Org. Biomol Chem, 2006, 4(8): 1520-1528. doi: 10.1039/b600960c
[6] LIU H C, DU L, ZHU T J, et al. Two new citrinin dimers from a volcano ash-derived fungus, Penicillium citrinum HGY1-5[J]. Helv Chim Acta, 2010, 93(11): 2224-2230. doi: 10.1002/hlca.v93.11
[7] NI M, LIN W L, YANG P, et al. A novel citrinin derivative from the marine-source fungus Penicillium citrinum[J]. Acta Pharm, 2015, 50(2): 203-206.
[8] LIU Y, LI X M, MENG L H, et al. Bisthiodiketopiperazines and acorane sesquiterpenes produced by the marine-derived fungus Penicillium adametzioides AS-53 on different culture media[J]. J Nat Prod, 2015, 78(6): 1294-1299. doi: 10.1021/acs.jnatprod.5b00102
[9] 申光辉, 郑丽君, 张志清, 等. 解淀粉芽胞杆菌PC2产抑菌物质培养基及发酵条件优化[J]. 微生物学通报, 2017, 44(6): 1358-1369. [10] 王昊鹏, 吴黎明, 赵柳微, 等. 烟曲霉素发酵培养基的优化研究[J]. 食品工业科技, 2017: 1-14. [11] RUQAYYAH T I D, JAMAL P, ALAM M Z, et al. Application of response surface methodology for protein enrichment of cassava peel as animal feed by the white-rot fungus Panus tigrinus M609RQY[J]. Food Hydrocolloid, 2014, 42: 298-303. doi: 10.1016/j.foodhyd.2014.04.027
[12] MUTHUKUMAR V, RAJESH N, VENKATASAMY R, et al. Mathematical modeling for radial overcut on electrical discharge machining of incoloy 800 by response surface methodology[J]. Procedia Materials Science, 2014, 6: 1674-1682. doi: 10.1016/j.mspro.2014.07.153
[13] YANG P, FANG M, LIU Y W. Optimization of a phase adjuster in a thermo-acoustic stirling engine using response surface methodology[J]. Energy Procedia, 2014, 61: 1772-1775. doi: 10.1016/j.egypro.2014.12.209
[14] 李莉, 张赛, 何强, 等. 响应面法在试验设计与优化中的应用[J]. 实验室研究与探索, 2015, 34(8): 41-45. doi: 10.3969/j.issn.1006-7167.2015.08.011 [15] 梁新乐, 黄莹莹, 张虹, 等. 响应面法优化桔青霉F-5-5核酸酶P1发酵培养基碳氮源[J]. 核农学报, 2011, 25(1): 57-61. [16] 谢祥聪, 刘琴英, 蒋冬花, 等. 淡色生赤壳菌(Bionectriao chroleuca)Bo-1菌株产生抗菌物质的发酵条件优化[J]. 生物技术通报, 2014(1): 166-170. [17] 郭雷, 朱文成, 刘玮炜, 等. 抗菌活性海洋真菌HN4-13的鉴定及其发酵优化[J]. 微生物学通报, 2013, 40(6): 951-958. [18] 胡一峰, 曹一岚, 黄俊龙. 美伐他汀发酵菌种筛选和发酵条件优化[J]. 浙江化工, 2012, 43(2): 4-6. doi: 10.3969/j.issn.1006-4184.2012.02.002 [19] 喻晨, 张亚雄, 赵劼, 等. 响应面法优化桔青霉产核酸酶P1培养基[J]. 食品科学, 2011, 32(17): 283-286. [20] MENG L H, LIU Y, LI X M, et al. Citrifelins A and B, citrinin adducts with a tetracyclic framework from cocultures of marine-derived isolates of Penicillium citrinum and Beauveria felina[J]. J Nat Prod, 2015, 78(9): 2301-2305. doi: 10.1021/acs.jnatprod.5b00450