SSR analysis on genetic diversity of peach cultivars and comparison of different genetic similarity coefficients
-
摘要:目的
利用NCBI数据库中桃的EST信息开发SSR引物, 分析成都平原主栽桃品种的遗传多样性,探讨各遗传相似系数在桃SSR分析中的适用性,以期为桃种质资源的研究提供参考。
方法利用来自NCBI-EST数据库的100对SSR引物对12个不同形态的桃品种进行PCR扩增,分析筛选20对多态性好、稳定性高且均匀分布于桃8个连锁群的引物对40个桃品种进行PCR扩增,并采用5个遗传相似系数分别对扩增结果进行分析。
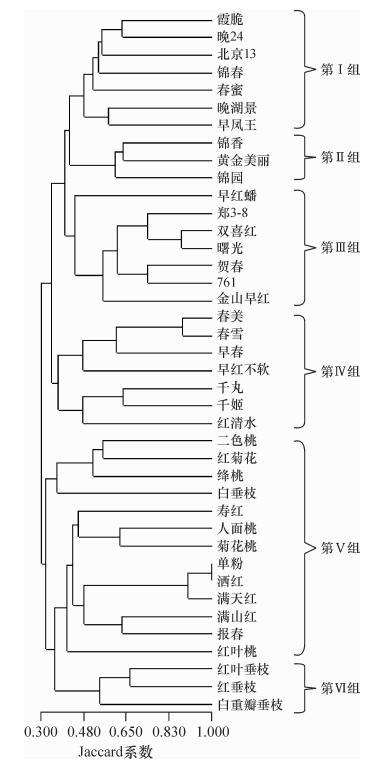
结果使用20对引物共检测出68个多态性等位基因位点,每对引物的等位基因数在2~5之间, 平均为3.4。多态信息含量在0.36~0.73间变化,平均为0.54。Jaccard相似系数为0.106~1.000; Jaccard系数的共表型相关系数rc最高,为0.772; Dice系数次之,为0.719; Jaccard和Dice系数的系统树一致性最高,CIc为1。Nei’s基因多样性指数在种级水平上和类群水平上分别为0.603和0.374,相应的Shannon’s信息指数分别为1.041和0.591。40个桃品种在聚类中首先分为观赏桃和食用桃2大类,再细分为各类群,聚类结果与传统系谱基本吻合。
结论基于NCBI数据库中桃的EST信息开发的SSR引物多态性较好,可用于桃的SSR分析。Jaccard和Dice系数适合桃的SSR分析。桃品种总体遗传多样性较丰富,但水蜜桃品种仍需加强种质创新。
Abstract:ObjectiveTo develop SSR primers based on peach ESTs in NCBI database, analyze genetic diversity of the major peach cultivars in Chengdu plain, investigate the applicability of different genetic similarity coefficients in SSR analysis of peach, and provide references for research on peach germplasm resources.
MethodIn total 100 pairs of SSR primers selected from NCBI-EST database were used in PCR amplification for 12 peach cultivars with various morphology. Twenty primers which had high polymorphism and stability, and evenly distributed in eight linkage groups were selected for PCR amplification of 40 peach cultivars. Five genetic similarity coefficients were used to analyze the amplification results.
ResultUsing 20 SSR primers, 68 polymorphic alleles were detected. The number of alleles per locus ranged from 2 to 5, with an average of 3.4. The polymorphism information content ranged from 0.36 to 0.73, and with an average of 0.54. The Jaccard genetic similarity coefficient ranged from 0.106 to 1.000. The Cophenetic correlation coefficient (rc) of Jaccard coefficient was the highest being 0.772, followed by Dice coefficient being 0.719. The consistency of consensus trees between Dice and Jaccard coefficients were the highest with CIc being 1.000. Nei's gene diversity index was 0.603 and 0.374 respectively at the species level and the level of different classified groups, the corresponding Shannon's information index was 1.041 and 0.591 respectively. The 40 peach cultivars were firstly divided into two major categories which were ornamental peach and edible peach, and then subdivided into different types of groups. The clustering result basically coincided with the traditional pedigree.
ConclusionThe SSR primers developed based on peach ESTs in NCBI database have high polymorphism and can be used for SSR analysis of peach. Jaccard and Dice coefficients are the most appropriate for SSR analysis of peach. In general, the genetic diversity of peach is abundant, but variation of the honey peach group is relatively low, and germplasm innovation of this group needs to be improved.
-
Keywords:
- peach /
- genetic diversity /
- genetic similarity coefficient /
- EST-SSR /
- primer screening
-
体内脂肪过度沉积对人体健康和动物胴体品质有不良影响[1],降低体内脂肪沉积可改善动物胴体品质[2-3],提高人体健康。钙作为机体必需元素,参与脂肪沉积调控[4-5],但其具体作用及调节机制尚不完全清楚。因此,研究钙对动物机体脂肪沉积的影响及机制,对于提高动物胴体品质及人体健康具有重要意义。
肠道菌群数量众多,作用广泛,对机体健康具有重要意义[6]。肠道菌群和肥胖的关系是现在研究的热点之一。有研究报道,肥胖与肠道细菌分布有关,厚壁菌门Firmicutes多于拟杆菌门Bacteroidetes导致肠道更有效吸收食物中的热量从而导致肥胖[7]。但目前的研究中,有关钙离子是否通过改变肠道菌群来调控肥胖的报道却较少。因此,研究钙离子对肠道菌群的影响有利于进一步深入了解钙离子调控脂肪沉积的机制。
本试验以C57BL小鼠为试验对象,研究饮水添加氯化钙对高脂日粮饲喂小鼠的脂肪沉积的影响,同时分析钙离子对肠道菌群的影响,以及肠道菌群与脂肪沉积可能的关联,旨在为提高动物胴体品质和人体健康提供理论依据。
1. 材料与方法
1.1 试验动物与试验设计
选用27只4周龄C57BL雄性小鼠(购于广东省医学实验动物中心),试验用的高脂日粮(High-fat diet, HFD)购于广东省医学实验动物中心,氯化钙购于Sigma公司。预饲小鼠正常日粮1周后,根据体质量随机分为2组:高脂组(HFD组),n=15,饲喂高脂日粮,脂肪提供60%的能量;高脂日粮+饮水添加氯化钙组(HFD+Ca2+组),n=12,饲喂高脂日粮,脂肪提供60%的能量,饮水中添加16.647 g/L的氯化钙。将小鼠分笼饲养,自由采食和饮水;每天光照与黑暗各12 h,环境温度控制在(25±1) ℃,湿度(60±5)%。饲养13周后,通过颈部脱臼方式处死小鼠,采集小鼠附睾脂肪、皮下脂肪组织并称质量。并在小鼠饲养至10~12周时采集小鼠的新鲜粪便样,于–80 ℃条件下进行保存。
1.2 测定指标及方法
饲养期间每周测定小鼠体质量和体内脂肪含量,体内脂肪含量通过核磁共振成像仪(上海纽迈科技有限公司,MesoQMR23-060H型)进行测定。样品采样后测定小鼠附睾脂肪、皮下脂肪质量。粪便样菌群送公司(上海微基生物科技有限公司)进行16S rRNA高通量测序,测定指标包括菌群丰度,菌群多样性。
1.3 统计分析
数据结果用平均值±标准误表示,统计分析采用SigmaPlot 12.5软件分析,采用t检验比较不同分组的统计差异。
2. 结果与分析
2.1 饮水添加氯化钙对高脂日粮饲喂小鼠体质量和脂肪沉积的影响
本研究中,与高脂组相比,饮水添加氯化钙使小鼠的体质量(图1A)、体脂含量(图1B)、皮下脂肪指数(图1C)和附睾脂肪指数(图1D)分别下降了12.85%、32.69%、26.65%和18.60%,差异均显著。
![图 1 饮水添加氯化钙对高脂日粮饲喂小鼠体质量(a)、体脂含量(b)、皮下脂肪指数(c)和附睾脂肪指数(d)的影响]() 图 1 饮水添加氯化钙对高脂日粮饲喂小鼠体质量(a)、体脂含量(b)、皮下脂肪指数(c)和附睾脂肪指数(d)的影响“*” 和 “**” 分别表示与HFD组差异达到0.05和0.01的显著水平(t检验)Figure 1. Effects of calcium chloride supplementation in drinking water on body weight (a), body fat content (b), subcutaneous fat index (c) and epididymal fat index (d) of mice fed high-fat diet“*” and “**” indicate the significant difference between HFD and HFD+Ca2+ at 0.05 and 0.01 levels, respectively (t test)
图 1 饮水添加氯化钙对高脂日粮饲喂小鼠体质量(a)、体脂含量(b)、皮下脂肪指数(c)和附睾脂肪指数(d)的影响“*” 和 “**” 分别表示与HFD组差异达到0.05和0.01的显著水平(t检验)Figure 1. Effects of calcium chloride supplementation in drinking water on body weight (a), body fat content (b), subcutaneous fat index (c) and epididymal fat index (d) of mice fed high-fat diet“*” and “**” indicate the significant difference between HFD and HFD+Ca2+ at 0.05 and 0.01 levels, respectively (t test)2.2 饮水添加氯化钙对高脂日粮饲喂小鼠粪便菌群的影响
2.2.1 小鼠粪便菌群的物种丰度及多样性
利用16S rRNA高通量测序方法,得到了粪便菌群的菌群多样性和菌群丰度。香农指数显示,饮水添加氯化钙可提高小鼠的粪便菌群群落多样性(图2A)。Chao指数显示,饮水添加氯化钙能提高小鼠的粪便菌群丰度(图2B)。
![图 2 饮水添加氯化钙对高脂日粮饲喂小鼠粪便菌群的香农指数(a)及Chao指数(b)的影响]() 图 2 饮水添加氯化钙对高脂日粮饲喂小鼠粪便菌群的香农指数(a)及Chao指数(b)的影响Figure 2. Effects of calcium chloride supplementation in drinking water on Shannon index (a) and Chao index (b) of mice fed high-fat diet
图 2 饮水添加氯化钙对高脂日粮饲喂小鼠粪便菌群的香农指数(a)及Chao指数(b)的影响Figure 2. Effects of calcium chloride supplementation in drinking water on Shannon index (a) and Chao index (b) of mice fed high-fat diet2.2.2 小鼠粪便菌群组成
PCA结果显示,高脂组的菌群聚集与饮水添加氯化钙组的菌群聚集有明显的分离情况(图3)。
![图 3 饮水添加氯化钙对高脂日粮饲喂小鼠粪便菌群的影响的主成分分析]() 图 3 饮水添加氯化钙对高脂日粮饲喂小鼠粪便菌群的影响的主成分分析Figure 3. Principal component analysis (PCA) of the effect of calcium chloride supplementation in drinking water on fecal flora of mice fed high-fat diet
图 3 饮水添加氯化钙对高脂日粮饲喂小鼠粪便菌群的影响的主成分分析Figure 3. Principal component analysis (PCA) of the effect of calcium chloride supplementation in drinking water on fecal flora of mice fed high-fat diet2.2.3 小鼠粪便菌群的物种相对丰度
16S rRNA测序结果表明,粪便菌群主要由6个细菌门构成:拟杆菌门Bacteroidetes、厚壁菌门Firmicutes、变形菌门Proteobacteria、脱铁杆菌门Deferribacteres、放线菌门Actinobacteria、软壁菌门Tenericutes,其中拟杆菌门和厚壁菌门为主要优势菌群。在门水平下,高脂饲喂小鼠粪便中的拟杆菌门、脱铁杆菌门、放线菌门的相对丰度较高,饮水添加氯化钙饲喂小鼠粪便中的厚壁菌门、变形菌门、软壁菌门的相对丰度较高。但这6种菌门的丰度在2个处理组的差异不显著。
在纲水平下,与高脂组相比,饮水添加氯化钙可显著降低丹毒丝菌纲Erysipelotrichia和放线菌纲Actinobacteria的相对丰度,并且显著提高与胆汁酸代谢相关的梭菌纲Clostridia的相对丰度,Negativicoccus只存在高脂组中。
在目水平下,与高脂组相比,饮水添加氯化钙可显著降低丹毒丝菌目Erysipelotrichia和放线菌目Coriobacteriaceae的相对丰度,并且显著提高梭菌目Clostridia的相对丰度。
在科水平下,与高脂组相比,饮水添加氯化钙可显著降低丹毒丝菌科Erysipelotrichia的相对丰度,并且显著提高瘤胃菌科Ruminococcaceae、优干菌科Eubacteriaceae、消化链球菌科Peptostreptococcaceae的相对丰度。动球菌科Planococcaceae、短杆菌科Brevibacteriaceae、肉杆菌科Carnobacteriaceae只存在于饮水添加氯化钙组。韦荣氏杆菌科Veillonellaceae只存在于高脂组。
在属水平下,与高脂组相比,饮水添加氯化钙能显著降低Allobaculum的相对丰度。与高脂组相比,饮水添加氯化钙能显著提高紫单胞菌属Parabacteroides、Anaerofustis的相对丰度。但土壤芽胞杆菌属Solibacillus、短杆菌属Brevibacterium、Turicibacter、Desemzia只存在于饮水添加氯化钙中。其中,微生物Turicibacter已被证明与抗炎表型相关[8]。
3. 讨论与结论
3.1 饮水添加氯化钙对高脂日粮饲喂小鼠脂肪沉积的影响
有文献报道,高钙源的摄入可以显著减少肥胖病人的腹部脂肪[9],降低肥胖小鼠体内皮下和附睾脂肪含量[10]。与以上报道相一致的是,本研究结果表明,饮水添加氯化钙显著减少高脂日粮饲喂小鼠体质量,减少体内皮下和附睾脂肪含量。但也有研究发现,高钙饮食并不能起到减轻肥胖的作用[11],这可能与添加钙的浓度、动物品种和动物个体差异的因素有关。因此,钙的摄入对动物和人的体质量及肥胖具有一定的调控作用。
本研究结果表明,虽然本试验钙的饮水添加方式与膳食钙的添加方式不同,但是也起到了降低肥胖的作用。机体的低钙水平会对正常的生理活动造成影响,而过高剂量的钙摄入对机体也会造成损伤,尤其是对心血管、肾和肝组织损伤明显。有研究报道,高钙饲料(钙质量分数为3%)的摄入未对大鼠肝肾脏造成损伤[12],而本研究添加的氯化钙质量浓度为16.647 g/L,因此,本研究添加的剂量应该对小鼠机体健康无明显影响。
3.2 饮水添加氯化钙对高脂日粮饲喂小鼠粪便菌群的影响
本试验中,香农指数和Chao指数显示,饮水添加氯化钙可提高高脂日粮饲喂小鼠粪便菌群群落多样性和菌群丰度。样品组成越相似,反映在PCA图中的距离越近。本试验中,通过PCA分析可知,饮水添加氯化钙已经影响高脂日粮饲喂小鼠粪便菌群样本的组成。Duncan等[13]研究表明使肥胖受试者体质量减轻的饮食可显著改变肠道微生物群的物种组成,但没有发现粪便细菌中拟杆菌和厚壁菌的比例在人类肥胖中起作用的证据。我们的研究结果发现饮水添加氯化钙显著降低体质量,其饲喂小鼠粪便中的厚壁菌门相对丰度增加,拟杆菌门相对丰度减少。但Ley等[14]发现肥胖人群粪便菌群里的厚壁菌门和拟杆菌门比值高于正常体质量人群,给予肥胖志愿者低脂饮食1年后,其体质量出现下降,而体内的厚壁菌门比例有了下降,拟杆菌门的比例则上升,这与本研究的结果不一致。但这一观点受到后续研究的挑战。最近的研究表明,肥胖与微生物多样性或丰度降低相关[15]。也有文献报道,随着身体质量指数(Body mass index, BMI)大小上升,肠道菌群多样性与丰富度呈下降趋势[16]。因此,饮水添加氯化钙可能是通过改变高脂日粮饲喂小鼠粪便菌群物种丰度与群落多样性来减少脂肪沉积。
肠道菌群虽然有很大的个体差异,但也有一定的共性[17]。有研究表明,粪便菌群和盲肠内容物的菌群变化趋势是相似的[18]。Nadeem等[18]报道,高钙饮食能提高盲肠内容物里的瘤胃菌科水平,降低盲肠内容物丹毒丝菌科水平。这与本试验的饮水添加氯化钙组能提高粪便菌群里的瘤胃菌科相对丰度和降低丹毒丝菌科相对丰度结果一致。
胆汁酸在脂质代谢中起重要的调节作用。胆汁酸不仅参与胆固醇的调节,而且在三酰甘油的代谢中也发挥着重要作用[19]。肠道微生物群通过促进初代胆汁酸的去共轭、脱氢和脱羟基作用对胆汁酸代谢具有深远的影响[20]。胆汁盐脱羧由胆汁盐水解酶催化。这种酶存在于各种肠细菌中,包括:梭菌属、拟杆菌属、乳杆菌属、双歧杆菌属和肠球菌属[21]。在饮水添加氯化钙的高脂日粮饲喂小鼠中梭菌属显著增加,说明饮水添加氯化钙可能影响小鼠粪便菌群中胆汁酸在脂质代谢方面的作用,从而显著减少高脂日粮饲喂小鼠的脂肪含量。
综上所述,饮水添加氯化钙能缓解高脂日粮诱导的肥胖,降低体质量和体内脂肪含量,而这可能与饮水添加氯化钙提高小鼠粪便菌群多样性和特定菌群丰度有关。研究结果为深入了解钙离子调节机体脂肪沉积机制以及提高动物胴体品质和人体健康具有重要意义。
-
表 1 试验种质和分类
Table 1 Experimental germplasm and classification
品种类群 种质 水蜜桃 霞脆 晚湖景 春蜜 早凤王 北京13 晚24 锦春 蟠桃 早红蟠 白桃 千姬 寿星桃 寿红 黄肉桃 锦香 锦园 黄金美丽 千丸 油桃 郑3-8 金山早红 贺春 761 双喜红 曙光 硬肉桃 春美 早春 春雪 早红不软 红清水 直枝桃 二色桃 红菊花 单粉 洒红 绛桃 满天红 人面桃 菊花桃 红叶桃 满山红 报春 垂枝桃 红叶垂枝 白重瓣垂枝 红垂枝 白垂枝  下载: 导出CSV
下载: 导出CSV
表 2 不同遗传相似系数的计算方法
Table 2 Calculation methods of five genetic similarity coefficients
相似系数 计算公式1) 区间 Simple Matching (SM) $(a + d)/(b + c)$ (0, 1) Jaccard (J) $a/\left\{ {\left[ {(a + d) + (b + c)} \right] - d} \right\}$ (0, 1) Russel and Rao (RR) $a/\left[ {(a + d) + (b + c)} \right]$ (0, 1) Dice $2a/(2a + b + c)$ (0, 1) Phi $(ad - bc)/\sqrt {(a + d)(c + d)(a + c)(b + d)} $ (-1, 1) 1)a表示品种i、j共有条带数;b表示品种i特有条带数;c表示品种j特有条带数;d表示品种i、j共有缺失条带数。
下载: 导出CSV
表 3 20对桃EST-SSR引物信息
Table 3 Information of 20 pairs of peach EST-SSR primers
引物 引物序列(5′→3′) 重复单位1) 产物/bp 总条带数量 多态性条带数量 多态信息含量 连锁群 DW344741 GAAGCACTGTGGGATTTGGT (TA)17 173 5 5 0.73 scaffold_1 AACGTTCATCCGTTGGACA DW346224 TTTCTCGGATTTGGAAAACG (GA)24 154 5 5 0.70 scaffold_1 TATCACGCGCCATAATTTGA DW345934 CAATCGTTCAAACACGCATC (TC)17 259 2 2 0.37 scaffold_1 AGGGCTGGATTTTGGTAGGT DW343823 ATTCCAGAGCACGCTTTCAC (TC)13 180 3 3 0.55 scaffold_2 TGCTTCCTGTTTGGCTCTTT DW344625 TTTCCTCGTGATATCTGCCC (AG)14 181 3 3 0.42 scaffold_2 TCCATTTTGAGGTGGTGGTT DW347146 AGTGAAGATGGTGGGAGTGG (AG)15 258 3 3 0.56 scaffold_3 TCTTTCGTCTCTTCCTCCCC AM289990 TCAGTAATTACCATTCAGAACT (TC)27 230 4 4 0.44 scaffold_3 CGCAGCCCAAATAAAATGAC DW347571 TCCTGCAGAGATTTTGGACA (AG)29 172 4 4 0.60 scaffold_3 TGCTTTCCGCAAGTTAAAGG DW346505 AGCGGATTTGTTTTGTTTGG (AT)12 209 5 5 0.68 scaffold_4 CAGGCTGTTACTGCCACAGA BU041775 CTCCTCACTCCTTCAACTCATT (CT)18 230 3 3 0.58 scaffold_4 CTTTAGCCCATGCAGAGAGG DT455234 CAACCCAGAAAAGCCAGAAC (CT)15 125 4 4 0.51 scaffold_4 TGCAGGGGAATTGATTTAGC DW342538 TTCCAACCAGTTCCTTCTGG (CT)24 233 3 3 0.59 scaffold_5 GGCTGTACTAGAAGCATTCCG DW341234 TCGTCAGATTCAGATGCAGG (AG)19 142 3 3 0.58 scaffold_5 AAAAGTTGCACCACCACCTC BU041198 TTGCCAGTTCATCATTGTTTG (TC)18 200 3 3 0.52 scaffold_6 ACCATTATGCCTTGGTCAGG AJ826975 TGTTGCTTTTTCAAGTTGCG (AG)17 168 2 2 0.36 scaffold_6 AAGGTTTACATCAGTGCCCG DW345117 CACCACCACTTTCACCTTCA (GA)29 273 3 3 0.52 scaffold_7 CAAATTTTGGGTTCCCCCTA BU048891 CTTTCTGTCGTTCCGCTACC (TC)8 147 4 4 0.68 scaffold_7 TCGCTTGAGACTGAATGACG BU040491 ACCGCAGCTGCTTAATCAAT (AT)9 166 3 3 0.51 scaffold_7 GGAACAAATTGCGAAAAGGA DW344783 TTTGGGCCAAGAAAATCAAG (GA)17 227 3 3 0.38 scaffold_8 CTTTCACTCTCGCCTCAACC BU044962 TCCCGATCTGAGACTAACCC (TC)19 195 3 3 0.59 scaffold_8 ATTCGCTCCAAAAACCAATG 1) 括号内的英文字符为简单重复序列,括号外的数字为该序列的重复次数。
下载: 导出CSV
表 4 基于不同相似系数的遗传相似性矩阵间的相关系数
Table 4 Correlation coefficients of genetic similarity matrix based on different similarity coefficients
相似系数 Phi SM Jaccard RR SM 0.996 Jaccard 0.976 0.959 RR 0.898 0.861 0.954 Dice 0.982 0.963 0.990 0.961
下载: 导出CSV
表 5 基于不同相似系数的UPGMA系统树的一致性指数
Table 5 The consistency indexes of UPGMA system trees based on different similarity coefficients
相似系数 Phi SM Jaccard RR SM 0.868 Jaccard 0.974 0.842 RR 0.500 0.474 0.500 Dice 0.974 0.842 1.000 0.500
下载: 导出CSV
表 6 桃品种遗传多样性水平
Table 6 Genetic diversity levels of peach cultivars
分类水平 类群 观测杂合度 期望杂合度 Nei’s基因多样性指数 Shannon’s信息指数 类群水平 水蜜桃 0.310 7 0.398 7 0.369 7 0.622 2 蟠桃 0.631 6 0.631 6 0.315 8 0.437 8 黄桃 0.337 5 0.446 4 0.390 6 0.623 6 油桃 0.461 4 0.420 6 0.384 9 0.614 8 硬肉桃 0.482 5 0.519 0 0.466 4 0.751 1 白桃 0.473 7 0.473 7 0.236 8 0.328 3 直枝桃 0.386 4 0.523 2 0.499 4 0.858 4 寿星桃 0.500 0 0.500 0 0.250 0 0.346 6 垂枝桃 0.404 2 0.518 8 0.452 4 0.735 0 平均 0.443 0 0.492 0 0.374 0 0.591 0 种级水平 0.405 0 0.611 0 0.603 0 1.041 0
下载: 导出CSV
-
[1] 李雄伟, 孟宪桥, 贾惠娟, 等.桃品种特异性荧光SSR分子标记数据库构建[J].果树学报, 2013, 30(6): 924-932. http://www.cnki.com.cn/Article/CJFDTOTAL-GSKK201306005.htm [2] 汪祖华, 庄恩及.中国果树志:桃卷[M].北京:中国林业出版社, 2001: 86-89. [3] 郭瑞, 李晓燕, 王力荣, 等.桃SRAP体系的优化及与SSR在桃品种鉴定上的比较[J].华北农学报, 2009, 24(4): 102-105. doi: 10.7668/hbnxb.2009.04.021 [4] 汪国平, 牛玉, 汪文毅, 等.番茄SSR标记在茄子及其他茄科作物上的通用性分析[J].华南农业大学学报, 2014, 35(4): 56-60. doi: 10.7671/j.issn.1001-411X.2014.04.011 [5] VERDE I, ABBOTT A G, SCALABRIN S, et al. The high-quality draft genome of peach (Prunus persica) identifies unique patterns of genetic diversity, domestication and genome evolution[J]. Nat Genet, 2013, 45(5): 487-494. doi: 10.1038/ng.2586
[6] BOUHADIDA M, MORENO M , GONZALO M J, et al. Genetic variability of introduced and local Spanish peach cultivars determined by SSR markers[J]. Tree Genet Genomes, 2011, 7(2): 257-270. doi: 10.1007/s11295-010-0329-3
[7] 张佳俊, 许淑芳, 张学英, 等.桃新品种'保佳红'亲缘关系的SSR鉴定[J].分子植物育种, 2014, 12(4): 712-719. http://www.cnki.com.cn/Article/CJFDTOTAL-FZZW201404014.htm [8] 廖安红, 杨鑫, 陈红.贵州桃种质资源ISSR标记体系建立及遗传多样性分析[J].分子植物育种, 2015, 13(12): 2773-2781. http://www.cnki.com.cn/Article/CJFDTOTAL-FZZW201512023.htm [9] 黄平, 崔娇鹏, 郑勇奇, 等.基于月季微卫星标记的7个遗传相似系数比较[J].林业科学, 2013, 49(1): 68-76. doi: 10.11707/j.1001-7488.20130111 [10] MANTEL N. The detection of disease clustering and a generalized regression approach[J]. Cancer Res, 1967, 27(2): 209-220. http://www.oalib.com/references/8626986
[11] SOKAL R R, ROHLF F J. Taxonomic congruence in the Leptopodomorpha re-examined[J]. Syst Zool, 1981, 30(3): 309-325. doi: 10.2307/2413252
[12] 史红丽, 韩明玉, 赵彩平.桃遗传多样性的SRAP和SSR标记分析[J].华北农学报, 2009, 24(6): 187-192. doi: 10.7668/hbnxb.2009.06.038 [13] 俞明亮, 马瑞娟, 沈志军, 等.应用SSR标记进行部分黄肉桃种质鉴定和亲缘关系分析[J].园艺学报, 2010, 37(12): 1909-1918. http://www.cnki.com.cn/Article/CJFDTOTAL-YYXB201012004.htm [14] MURGUA M, VILLASEOR J L. Estimating the effect of the similarity coefficient and the cluster algorithm on biogeographic classifications[J]. Ann Bot Fenn, 2003, 40(6): 415-421.
[15] 程中平, 黄宏文.桃不同类群的遗传多样性及其遗传结构的RAPD分析[J].武汉植物学研究, 2004, 22(1): 27-32. http://www.cnki.com.cn/Article/CJFDTOTAL-WZXY200401005.htm [16] 陈巍, 王力荣, 张绍铃, 等.利用SSR研究不同国家桃育成品种的遗传多样性[J], 果树学报, 2007, 24(5): 580-584. http://www.cnki.com.cn/Article/CJFDTOTAL-GSKK200705004.htm [17] 魏姗姗, 刘兴菊, 杨敏生, 等.基于成熟期的桃品种遗传多样性SSR分析[J], 北方园艺, 2014(12): 88-93. http://www.cnki.com.cn/Article/CJFDTOTAL-BFYY201412027.htm [18] 陈霁, 马瑞娟, 沈志军, 等.基于SSR标记的观赏桃亲缘关系分析[J].果树学报, 2011, 28(4): 580-585. http://www.cnki.com.cn/Article/CJFDTOTAL-GSKK201104008.htm [19] 葛志刚, 俞明亮, 马瑞娟, 等.蟠桃种质SSR标记的遗传多样性分析[J], 果树学报, 2009, 26(3): 300-305. http://www.cnki.com.cn/Article/CJFDTOTAL-GSKK200903010.htm [20] 陆苏瑀, 俞明亮, 马瑞娟, 等.硬肉桃品种群SSR标记的遗传多样性分析[J].植物遗传资源学报, 2010, 11(3): 374-379. http://www.cnki.com.cn/Article/CJFDTOTAL-ZWYC201003023.htm -
期刊类型引用(2)
1. 马春丽,赵林昀,扈瑞平,李丽,曹丽丽,红梅,包玉龙. 钙离子对高脂饮食小鼠肠道菌群的影响. 畜牧与饲料科学. 2025(01): 1-9 .  百度学术
百度学术
2. 李玉萍,田晶晶,张凯,夏耘,王广军,郁二蒙,李志斐,龚望宝,谢骏. 皇竹草对草鱼脂肪蓄积及肠道菌群组成的影响. 上海海洋大学学报. 2022(04): 915-928 . 百度学术
其他类型引用(1)