Influence of substituent groups on the antibacterial effect of pleuromutilin derivatives with thiadiazole moiety against Mycoplasma gallisepticum
-
摘要:目的
为研发抗鸡毒支原体感染的药物提供试验依据。
方法合成18种C14支链含噻二唑片段的截短侧耳素衍生物,采用高分辨质谱进行结构确证,通过体外药敏试验研究所合成苯环取代基衍生物对鸡毒支原体抑菌活性的影响。
结果化合物12、15和17对鸡毒支原体S6的最小抑菌质量浓度(Minimum inhibitory concentration,MIC)均为0.125 0 μg·mL-1,化合物18和对照泰妙菌素对鸡毒支原体S6的MIC值最高,均为0.015 6 μg·mL-1,化合物18氨基被苯甲酰基化后所得化合物17的抗菌活性减弱。
结论化合物18可作为候选化合物进行深入研究,以开发新的抗支原体感染药物。
Abstract:ObjectiveTo provide experimental data for the research and development of new antibacterials against Mycoplasma gallisepticum.
MethodEighteen analogues of pleuromutilin possessing thiadiazole moieties at the C14 chains were synthesized. High resolution mass spectrometry was used to verify the structures of the synthesized compounds. Influence of substituted benzene on the antibacterial effects of these pleuromutilin derivatives against M. gallisepticum were investigated by in vitro drug sensitivity tests.
ResultThe obtained MIC values revealed that analogues 12, 15 and 17 exhibited potential antibacterial activities against M. gallisepticum S6 strain with MIC of 0.125 0 μg·mL-1. Analogue 18 and Tiamulin (control) showed the most potent antibacterial activities against S6 strain in vitro with MIC of 0.015 6 μg·mL-1. Analogue 17, which was the product from benzoylation of the amino group of analogue 18, had lower antibacterial activity compared with analogue 18.
ConclusionAnalogue 18 can be used as a candidate compound for further development of antibacterials against M. gallisepticum.
-
草地贪夜蛾Spodoptera frugiperda是一种原产于美洲的热带和亚热带地区的杂食性害虫,严重威胁农业生产。其寄主范围广,能够为害玉米、水稻、棉花、大豆等80多种作物[1]。不同的草地贪夜蛾群体通常对特定作物具有取食趋性[2]。田间草地贪夜蛾主要分为玉米型(Corn strain)和水稻型(Rice strain)2个亚群[3]。玉米型主要取食玉米、高粱、棉花,而水稻型的取食性相对较为广谱,主要取食水稻、苜蓿、牧草、小米等作物[4]。除了取食偏好外,2个品系的草地贪夜蛾在生理特征、交配行为和性外激素的组成方面有很大的不同。但是,由于2种寄主型形态学和行为方面的相似性,在田间难以被区分开[5]。因此,对2种寄主型的草地贪夜蛾开展分子鉴定,将有助于了解该入侵物种的遗传结构,掌握害虫的入侵和扩散规律以及制定有效的防控策略[6-7]。
草地贪夜蛾亚群的分类主要根据田间采集的幼虫寄主范围,并结合分子标记的分化进行界定。基于线粒体细胞色素C氧化酶亚基I基因(COI)的分子标记不仅能够区分玉米型和水稻型2个亚群的,而且具备鉴别其亚型的能力,是目前最常用的有效分子条形码[7-10]。另外,有研究表明,根据Z染色体上编码磷酸丙糖异构酶基因(Tpi)开发的分子标记,能够更好地鉴定到草地贪夜蛾雌性“水稻型”和雄性“玉米型”的杂交型[7, 11]。2种标记有机结合能够提高鉴定的准确率[12-13]。
2018年底草地贪夜蛾通过中缅边境进入我国境内,目前已在云南、湖南、广西、广东等地发现其对玉米的为害[14]。广东省是水稻主要种植区,严密监测草地贪夜蛾是否存在为害水稻的风险显得格外重要[15]。另外,广东省为热带和亚热带季风气候区,草地贪夜蛾可在该省越冬,是可阻止其北迁的桥头堡[16]。本研究通过基于COI和Tpi 2个基因片段的分子标记,比较它们对入侵广州的草地贪夜蛾的鉴别能力,为监测虫源扩散规律以及预测预警虫情提供参考。
1. 材料与方法
1.1 供试昆虫采集
供试草地贪夜蛾于2019年5月12日在广州花都花山镇儒林村玉米地采集。寄主为喇叭口期的春玉米,所有样本均为4~6龄幼虫,6个样本分别标记为1#、2#、3#、4#、5#和6#。单头活体幼虫采集后放入含有新鲜玉米叶片的50 mL离心管,带回实验室进行形态学鉴定后,用液氮速冻储存于−80 ℃用于DNA提取。
1.2 DNA提取和扩增
采用DNA/RNA共提取试剂盒(TIANGEN,中国)分别提取单头幼虫DNA和RNA。虫体放入含有1.5 mL DNA抽提液的2 mL离心管内,加入钢珠,放入样品研磨机(Bullet Blender®,美国)处理后,按照试剂盒说明书操作。DNA使用NanoDrop分光光度计(2000,赛默飞世尔)测定质量浓度,稀释到10 ng/μL后,用作PCR扩增模板。
用于PCR扩增COI和Tpi部分基因片段的引物如表1所示,选用的PCR引物在COI基因的相对位置如图1所示。采用PrimeSTAR® Max DNA Polymerase PCR(TaKaRa,日本)进行PCR扩增,反应体系包括PrimeSTAR® Max DNA Polymerase PCR(2×)10 μL,上游引物和下游引物各0.4 μL,模板1 μL,加ddH2O至20 μL;PCR反应为:98 ℃预变性2 min;98 ℃变性10 s,60 ℃退火15 s,72 ℃延伸30 s,35个循环;72 ℃延伸5 min。
表 1 扩增COI和Tpi基因片段所用引物Table 1. Primers used for amplifying COI and Tpi gene fragments![图 1 用于单倍型分析的COI基因3 个片段]() 图 1 用于单倍型分析的COI基因3 个片段Figure 1. Three fragments of COI gene used for haplotype analysis
图 1 用于单倍型分析的COI基因3 个片段Figure 1. Three fragments of COI gene used for haplotype analysis使用15 g/L的琼脂糖凝胶电泳检测扩增片段特异性后,PCR产物由广州天一辉远基因科技有限公司进行切胶测序。对于测序结果为双峰或者低质量的样品,则将其PCR产物克隆到pEASY®-Blunt Cloning Vector后,每个样品各挑取10个克隆进行测序。
1.3 序列分析
利用Lastergene软件包对正反向测序序列进行人工拼接、峰型校正和序列比对。从GenBank数据库下载已经发布的草地贪夜蛾COI和Tpi基因序列。针对已报道的基于COI和Tpi基因序列的“水稻型”和“玉米型”单倍型特点,分别进行差异位点分析,确定本研究中草地贪夜蛾的具体亚型。
2. 结果与分析
2.1 COI标记鉴定草地贪夜蛾单倍型
利用标记COI-A,COI-B和COI-C对所采集的6个样品进行了寄主型分析。结果(图2)显示,有5个样品扩增出了预期大小的单一特异性条带,将其PCR产物用于直接测序。
![图 2 6头草地贪夜蛾COI-A、COI-B和COI-C标记的PCR扩增]() 图 2 6头草地贪夜蛾COI-A、COI-B和COI-C标记的PCR扩增Figure 2. PCR amplification of COI-A, COI-B and COI-C markers from six Spodoptera frugiperda
图 2 6头草地贪夜蛾COI-A、COI-B和COI-C标记的PCR扩增Figure 2. PCR amplification of COI-A, COI-B and COI-C markers from six Spodoptera frugiperda对COI-A的序列分析表明,来自4个样品(1#、2#、4#、6#)的序列与已报道的草地贪夜蛾水稻型(HM136593)仅存在一处碱基差异,与玉米型(HM136586)存在8处单碱基差异,说明根据COI-A标记鉴定的寄主型与实际样品来源寄主型并不一致(图3);对样品3#的COI-A产物测序分析发现其为双峰杂合,克隆重新测序后,分别获得了样品3#-1和3#-2两种单倍型。其中,3#-1与上述4个样品检测到的单倍型完全一致,3#-2则与Mythimna separate(KX863042) 对应的片段序列上存在1个碱基的差异,说明3#样品可能是草地贪夜蛾和M. separate的杂合(图3)。
![图 3 基于COI-A标记的单倍型序列比对]() 图 3 基于COI-A标记的单倍型序列比对HD majority:花都检测到的主要单倍体型;3#-1, 3#-2:样品3#的2种克隆类型Figure 3. Haplotype sequence slignments based on COI-A markersHD majority: Major haplotype in Huadu; 3#-1, 3#-2: Two types of clones from sample 3#
图 3 基于COI-A标记的单倍型序列比对HD majority:花都检测到的主要单倍体型;3#-1, 3#-2:样品3#的2种克隆类型Figure 3. Haplotype sequence slignments based on COI-A markersHD majority: Major haplotype in Huadu; 3#-1, 3#-2: Two types of clones from sample 3#进一步用COI-B和COI-C标记对样品进行寄主型鉴定,结果表明,除了样品5#无法扩增出条带之外,其余的5个样品均扩增出单一的预期条带。PCR产物测序结果显示所测样品序列完全一致且为水稻型(图2)。
将COI-A、COI-B和COI-C等3个标记的结果相比较,发现COI-A能够区分出草地贪夜蛾和M. separate的2种单倍型,而COI-B和COI-C仅能区分出草地贪夜蛾1种单倍型,说明虽然基于COI基因片段序列鉴定到的草地贪夜蛾寄主型与实际采集寄主种类不相符,但是,COI-A标记具有较好的物种鉴别能力,能够区分出杂合类型。
2.2 TPi标记鉴定草地贪夜蛾单倍型
基于Tpi基因片段对所采集的6个样品进行了寄主型分析。除了样品5#无法扩增出条带之外,其余的5个样品均扩增出了预期大小的单一特异性条带(图4)。
![图 4 6头草地贪夜蛾Tpi基因片段的PCR扩增]() 图 4 6头草地贪夜蛾Tpi基因片段的PCR扩增Figure 4. PCR amplification of Tpi gene fragment from six Spodoptera frugiperda
图 4 6头草地贪夜蛾Tpi基因片段的PCR扩增Figure 4. PCR amplification of Tpi gene fragment from six Spodoptera frugiperda对PCR产物进行双向测序并分析其序列差异,结果显示,所检测的序列与已报道玉米型一致,说明基于Tpi基因片段的鉴定结果与样品的寄主来源是一致的。样品2#和样品3#,在第126和127碱基处出现GA/AT的杂合双峰(图5),说明入侵广东的草地贪夜蛾种类并不单一。
![图 5 基于Tpi基因标记的单倍型序列比对]() 图 5 基于Tpi基因标记的单倍型序列比对红色标记为花都样品2#和3#出现杂合(RW)的位置Figure 5. Haplotype sequence alignments based on Tpi gene markerRed line indicates the heterozygous base pairs (RW) in sample 2# and 3# from Huadu
图 5 基于Tpi基因标记的单倍型序列比对红色标记为花都样品2#和3#出现杂合(RW)的位置Figure 5. Haplotype sequence alignments based on Tpi gene markerRed line indicates the heterozygous base pairs (RW) in sample 2# and 3# from Huadu3. 结论与讨论
本研究采集了入侵广州市的草地贪夜蛾样本,利用基于COI和Tpi基因片段的分子标记进行了寄主型的鉴定。基于COI-A、COI-B和COI-C标记的鉴定结果均显示入侵广州的草地贪夜蛾寄主单倍型为水稻型,与实际采集寄主不相符,可能原因是该标记不适合于检测入侵广州的草地贪夜蛾群体。但基于COI-A的标记能够区分草地贪夜蛾与其他近源种的基因型,为研究入侵草地贪夜蛾的田间行为和扩散提供丰富的信息。而COI-B和COI-C只鉴定到1种单倍型。COI-A和COI-B标记事实上扩增的是COI基因的同一片段,差别在于COI-A标记扩增的片段较小,且使用的是简并引物。因此,COI-A标记具有比COI-B标记和COI-C标记更强的鉴别能力。
Tpi基因标记的鉴定结果显示入侵广州的草地贪夜蛾寄主型为玉米型,与实际采集的寄主一致。基于Tpi基因标记,入侵广州的草地贪夜蛾群体被检测到具有2种单倍亚型,在该片段第126和第127位碱基具有RW杂合。先前报道的入侵云南的群体只有一种单倍型[12],说明草地贪夜蛾可能在扩散的过程中出现遗传杂交现象。
草地贪夜蛾是联合国粮农组织全球预警的跨国界迁飞性重大害虫。目前,入侵我国的草地贪夜蛾主要为害玉米,未见在水稻上定殖为害。华南地区是我国水稻主产区,应慎重评估草地贪夜蛾为害水稻的风险。对于“玉米型”和“水稻型”的区分,仅依靠个别基因的片段进行评估是不全面的[6]。基于微卫星标记的分子指纹分析可以较全面地了解草地贪夜蛾的遗传结构动态变化规律[17]。获得与寄主取食相关基因的信息进而开发基因功能性分子标记,是精准鉴定草地贪夜蛾寄主单倍型的前提[18]。
-
![]()
图 1 截短侧耳素衍生物的合成路线
试剂及反应条件:(ⅰ)对甲苯磺酰氯,吡啶,0 ℃,3 h,产率56.3%;(ⅱ) 2-氨基-5-巯基噻二唑,NaOH,0 ℃,2 h ; (ⅲ)乙酸乙酯,三乙胺,50~70 ℃,48 h。
Figure 1. The synthetic route of pleuromutilin derivatives
表 1 18种衍生物精确质量数和18种衍生物及泰妙菌素对S6支原体MIC结果
Table 1 The MIC against Mycoplasma gallisepticum S6 strain and HR-MS of 18 pleuromutilin derivatives and Tiamulin
化合物 R1) HR-MS([M+H]+) MIC/(μg·mL-1) 1 
616.233 1 0.250 0 2 
616.233 0 0.500 0 3 
616.231 0 0.250 0 4 
632.203 5 0.250 0 5 
632.202 9 0.500 0 6 
632.204 4 0.250 0 7 
643.227 2 0.500 0 8 
643.227 6 0.500 0 9 
643.228 4 0.500 0 10 
666.230 1 0.500 0 11 
666.229 9 0.500 0 12 
612.258 2 0.125 0 13 
612.258 4 0.250 0 14 
612.258 2 0.500 0 15 
628.253 6 0.125 0 16 
628.253 9 0.250 0 17 
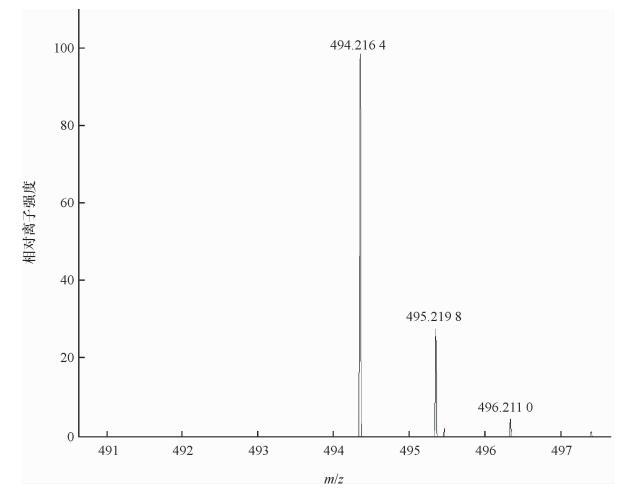
598.242 9 0.125 0 18 494.216 4 0.015 6 泰妙菌素 0.015 6 1)苯环上不同取代基的苯甲酰氯。  下载: 导出CSV
下载: 导出CSV
-
[1] 冯元璋. 鸡毒支原体感染及危害[J]. 中国家禽, 2008, 15(12): 45-47. http://www.cnki.com.cn/Article/CJFDTOTAL-ZGJQ200812016.htm [2] 陈凤梅,牛钟相,程光民,等. 鸡毒支原体研究进展[J]. 动物医学进展, 2004, 25(3): 56-59. http://www.cnki.com.cn/Article/CJFDTOTAL-LXLX201403010.htm [3] 纪新花,王永强,张新民. 鸡支原体病的特点及防控策略[J]. 养禽与禽病防治, 2010(9): 34-36. http://www.cnki.com.cn/Article/CJFDTOTAL-YQYF201009014.htm [4] 贾志江,刘娣琴,杨红梅. 鸡毒支原体感染的流行病学分析及综合防制[J]. 畜牧兽医杂志, 2010, 29(3):117-118. http://www.cnki.com.cn/Article/CJFDTOTAL-XMSZ201003059.htm [5] KAVANAGH H R. Antibiotic substances from basidiomycetes: VIII: Pleurotus multilus (Fr.) Sacc. and Pleurotus passeckerianus Pilat[J]. P Natl Acad Sci USA, 1951, 37(9): 570-574. doi: 10.1073/pnas.37.9.570
[6] TANG Y Z, LIU Y H, CHEN J X. Pleuromutilin and its derivatives:The lead compounds for novel antibiotics[J]. Mini Rev Med Chem, 2012, 12(1): 53-61. doi: 10.2174/138955712798868968
[7] 邹懿,黄宇琪,胡昌华. 截短侧耳素及其衍生物的研究进展[J]. 中国抗生素杂志, 2009, 34(2): 65-68. http://youxian.cnki.com.cn/yxdetail.aspx?filename=ZKSS20170627000&dbname=CAPJ2015 [8] SHANG R, WANG S, XU X, et al. Chemical synthesis and biological activities of novel pleuromutilin derivatives with substituted amino moiety[J]. PLoS One, 2013, 8(12): e82595. doi: 10.1371/journal.pone.0082595
[9] YI Y, YANG G, ZHANG C, et al. Synthesis and evaluation of novel pleuromutilin derivatives with a substituted pyrimidine moiety[J]. Eur J Med Chem, 2015, 101(13): 179-184. http://www.sciencedirect.com/science/article/pii/S0223523416310005
[10] LING Y, WANG X, WANG H, et al. Design, synthesis, and antibacterial activity of novel pleuromutilin derivatives bearing an amino thiazolyl ring[J]. Arch Pharm, 2012, 345(8): 638-646. doi: 10.1002/ardp.v345.8
[11] WANG X, LING Y, WANG H, et al. Novel pleuromutilin derivatives as antibacterial agents: Synthesis, biological evaluation and molecular docking studies[J]. Bioorg Med Chem Lett, 2012, 22(19): 6166-6172. doi: 10.1016/j.bmcl.2012.08.021
[12] SHANG R, PU X, XU X, et al. Synthesis and biological activities of novel pleuromutilin derivatives with a substituted thiadiazole moiety as potent drug-resistant bacteria inhibitors[J]. J Med Chem, 2014, 57(13): 5664-5678. doi: 10.1021/jm500374c
[13] SHANG R, WANG G, XU X, et al. Synthesis and biological evaluation of new pleuromutilin derivatives as antibacterial agents[J]. Molecules, 2014, 19(11): 19050-19065. doi: 10.3390/molecules191119050
[14] 王波,李健,汤有志,等. 空间位阻对丁胺侧链取代截短侧耳素衍生物鸡毒支原体抑制活性的影响[J]. 中国畜牧兽医, 2012, 39(12): 101-103. doi: 10.3969/j.issn.1671-7236.2012.12.024 [15] DAVIDOVICH C, BASHAN A, AUERBACH-NEVO T, et al. Induced-fit tightens pleuromutilins binding to ribosomes and remote interactions enable their selectivity[J]. P Natl Acad Sci USA, 2007, 104(11): 4291-4296. doi: 10.1073/pnas.0700041104
[16] ZHANG C, YI Y, CHEN J, et al. In vivo efficacy and toxicity studies of a novel antibacterial agent: 14-O-[(2-amino-1,3,4-thiadiazol-5-yl)thioacetyl] mutilin[J]. Molecules, 2015, 20(4): 5299-5312. doi: 10.3390/molecules20045299
-
期刊类型引用(4)
1. 邓文林. 乡土阔叶树种示范林建设——以韶关市国有仁化林场为例. 农村科学实验. 2024(18): 133-135 .  百度学术
百度学术
2. 何潇,雷相东,段光爽,丰庆荣,张逸如,冯林艳. 气候变化对落叶松人工林生物量生长的影响模拟. 南京林业大学学报(自然科学版). 2023(03): 120-128 . 百度学术
3. 康梓杭,倪苗,邓晶,吴庆书. 海口市10种行道树最优生长模型研究. 热带生物学报. 2020(01): 79-83 . 百度学术
4. 曹磊,刘晓彤,李海奎,雷渊才. 广东省常绿阔叶林生物量生长模型. 林业科学研究. 2020(05): 61-67 . 百度学术
其他类型引用(9)