Effects of applying compound fertilizer on yield, nutrient absorption and oligosaccharide accumulation of Morinda officinalis
-
摘要:目的
研究复合肥施用量对巴戟天产量、养分吸收和主要有效成分寡糖累积量的影响,为巴戟天的施肥管理提供理论依据。
方法以巴戟天为研究对象,通过盆栽试验,设置每株施用复合肥0、2、4、6、8和10 g (CK、T1、T2、T3、T4和T5)共6个处理,测定巴戟天在不同施肥处理下的产量、养分含量和寡糖含量,分析各指标间的相关性,并运用主成分分析方法进行综合评价。
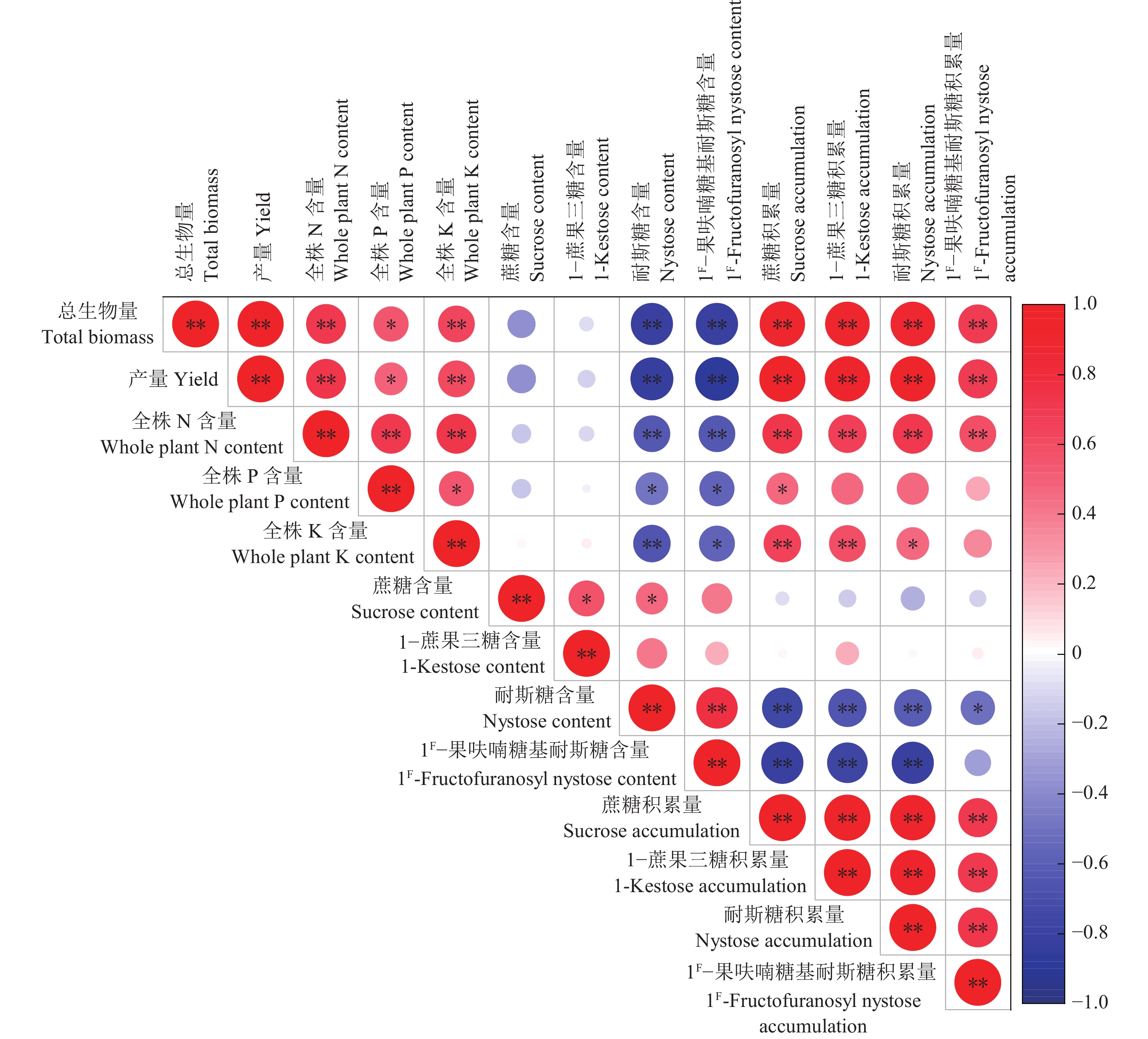
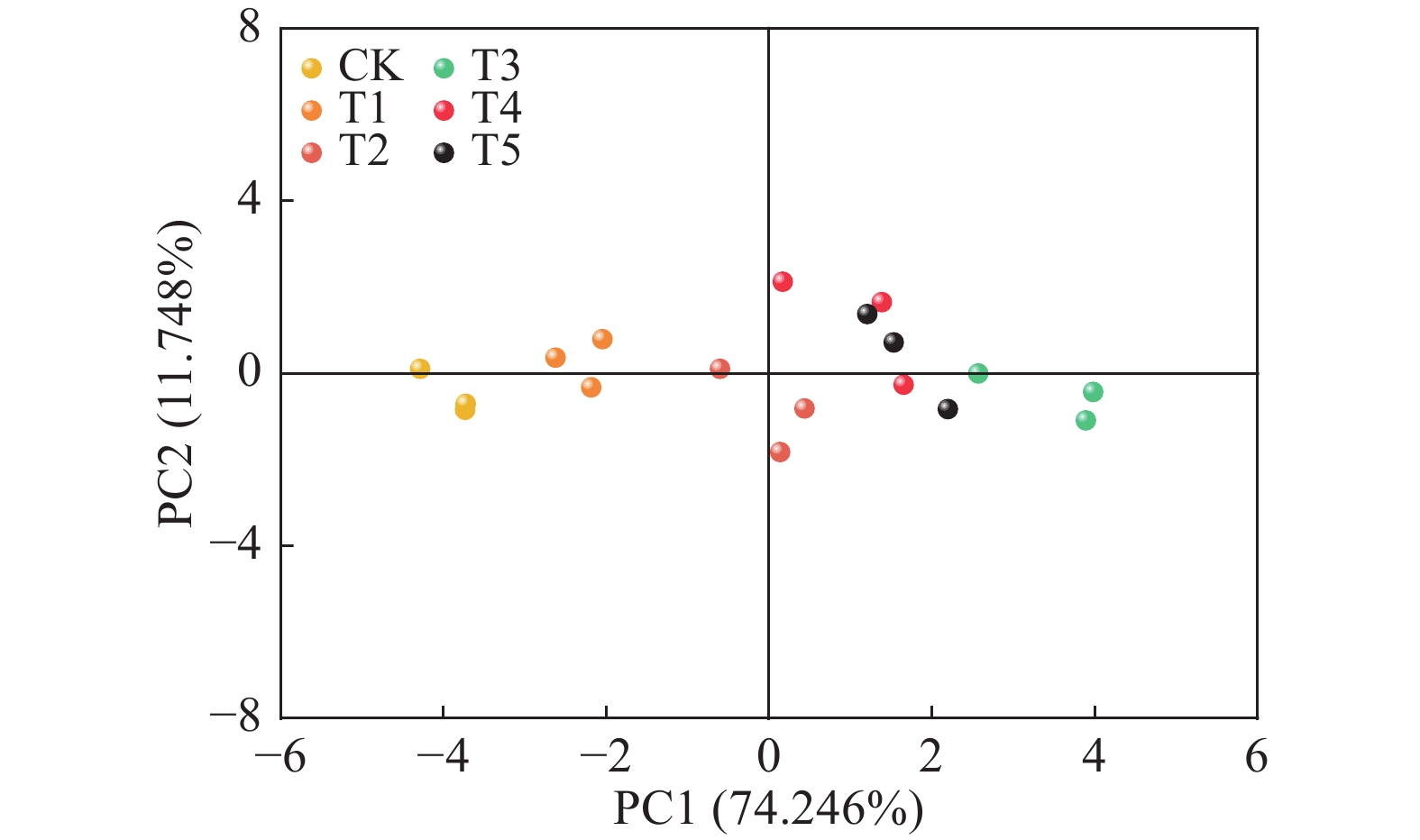
结果施用复合肥可以显著提高巴戟天产量和N、P、K含量,并有利于提高寡糖的累积量。其中,单株施肥量为6 g处理的巴戟天产量和寡糖总累积量最高,与CK相比,分别提升114.47%和75.36%。施肥在一定程度上降低了巴戟天的寡糖含量,与CK相比,降幅最高达35.65%。巴戟天对复合肥中养分的利用效率表现为N > K > P,其中T3处理N、P、K累积量均高于其他处理。影响巴戟天寡糖累积量的最关键因素是产量,巴戟天产量与全株N/P/K含量、4种寡糖累积量呈显著或极显著正相关,但耐斯糖、1F−果呋喃糖基耐斯糖含量与产量、全株N/P/K含量呈显著或极显著负相关。主成分分析的综合得分由高到低为T3 > T5 > T4 > T2 > T1 > CK,T3处理最佳。
结论建议巴戟天的种植以N肥为主,适量补充K和P肥,适当施肥可显著提高巴戟天药材产量和寡糖累积量。
Abstract:ObjectiveTo study the effects of compound fertilizer application amount on yield, nutrient absorption and accumulation of main effective component oligosaccharides of Morinda officinalis, and provide a theoretical basis for the fertilization management of M. officinalis.
MethodUsing M. officinalis as the research object, through the pot experiment, a total of six treatments were set up to apply 0, 2, 4, 6, 8 and 10 g compound fertilizer (CK, T1, T2, T3, T4 and T5) per plant. The yield, nutrient content and oligosaccharide content of M. officinalis under different fertilization treatments were determined. Then we analyzed the correlation between the indicators, and comprehensively evaluated them by principal component analysis.
ResultThe application of compound fertilizer significantly improved the yield of M. officinalis, increased the N, P and K contents, and contributed to the increase in the accumulation of oligosaccharides. The highest yield and total oligosaccharide accumulation were achieved in the treatment of 6 g compound fertilizer per plant (T3), compared with no fertilizer (CK), with increases of 114.47% and 75.36% respectively. Fertilizer application reduced the oligosaccharide content of M. officinalis to some extent, with a maximum decrease of 35.65% compared to CK. The utilization efficiency of nutrient in compound fertilizer by M. officinalis showed N > K > P, and the accumulations of N, P and K in T3 treatment were higher than those of other treatments. The yield was the most critical factor affecting the accumulation of oligosaccharides in M. officinalis. The yield of M. officinalis was significantly or highly significantly positively correlated with the whole plant N, P, K contents and the accumulation of four oligosaccharides, but the contents of nystose and 1F-fructofuranosyl nystose were significantly or highly significantly negatively correlated with the yield and the whole plant N, P, K contents. The comprehensive score of principal component analysis from high to low was T3>T5>T4>T2>T1>CK, with T3 treatment being the best.
ConclusionIt is suggested that the cultivation of M. officinalis should focus on N fertilizer, and supplement K and P fertilizer appropriately. Proper fertilization can significantly increase the yield of M. officinalis and the accumulation of oligosaccharides.
-
Keywords:
- Morinda officinalis /
- Compound fertilizer /
- Yield /
- Oligosaccharide /
- Nutrient absorption
-
山羊Capra hircus繁殖力影响养羊业的经济效应,受遗传、营养、管理水平等因素影响,是复杂的数量性状。母羊的产羔数是山羊繁殖性能之一,由排卵数决定,主要受遗传因素影响,与卵泡发育息息相关[1]。卵泡发育是一个极其复杂的过程,在发育过程中绝大多数卵泡会闭锁,只有少数卵泡能成熟并排卵[2-3]。
目前,关于整个卵巢的研究越来越多[4-5],但卵巢作为一个动态发育的生殖器官,包含不同发育阶段的卵泡,如原始卵泡、生长卵泡、成熟卵泡等。因此,在研究卵巢时,有必要对其细分后再开展研究。Terenina等[6]对猪卵巢健康小卵泡和闭锁小卵泡转录组的研究发现CCDC80、GADD45A、ZNF628等基因在闭锁小卵泡中的表达量超过健康小卵泡5倍,认为可能是卵泡闭锁的标志基因。Zhang等[7]在猪健康卵泡和早期闭锁卵泡的转录组研究中发现450个显著差异表达基因。李鹏飞等[8-9]对牛的优势卵泡和从属卵泡进行转录组测序发现差异显著的PRSS35、PTGRF和ARID4B等基因与卵泡发育相关。Hatzirodos等[10]在牛健康大卵泡和健康小卵泡的转录组研究中发现,在大卵泡中STAT4和XBP1基因表达上调,MGEA5基因下调;而KIT、IHH和MEST基因在小卵泡中最为活跃。吴阳升等[11]通过分析绵羊小卵泡和中卵泡转录组发现,在中卵泡表达上调的基因有IGFBP1、INHBA和TIMP1等,下调的有C-FOS、EGR1、FOSB等。综上,猪、牛和绵羊不同发育阶段卵泡的基因表达模式不同,然而山羊的卵泡转录组研究以及卵巢基质和卵泡之间的基因表达模式鲜见报道。
川中黑山羊是我国优秀的地方山羊品种[12],经过长期的自然和人工选择,繁殖性能突出,遗传性能稳定,母羊平均产羔率为236.78%,羔羊成活率为91%[13]。目前对川中黑山羊繁殖性能的研究较少。转录组包括编码RNA和非编码RNA,指在某一特定条件下细胞或组织内所有转录产物的总和,从整体水平研究基因结构和功能[14]。转录组测序是通过高通量测序技术,对生物体全部基因的测序,反映生命体处于不同环境条件下的不同基因表达水平及调控模式[15]。本研究以发情期的川中黑山羊为研究对象,应用高通量测序技术对卵巢基质、大卵泡和小卵泡进行转录组测序,比较分析三者的转录水平差异,探索卵巢基质、大卵泡和小卵泡间的mRNA表达模式。旨在从转录组水平揭示卵巢不同部位的遗传差异,丰富山羊卵巢基质、大卵泡和小卵泡的转录组数据信息,筛选出的基因可为山羊卵泡发育提供一定的基因资源和新的研究方向。
1. 材料与方法
1.1 试验动物和样品采集
选用南方某种羊场10头健康的川中黑山羊母羊,年龄3.5~4.5岁。采用前列腺素1次注射法对每只母羊注射氯前列烯醇0.1 mg,24 h后第1次发情,18 d后确认第2次发情,采用公羊试情法确认母羊是否发情,以母羊摇尾、站立并接受爬跨视为发情[16-19]。发情后24 h内进行屠宰并采集卵巢。
卵巢用PBS缓冲液洗3次,分离小卵泡(Small follicle,SF,d<3 mm)、大卵泡(Large follicle,LF,d>1 cm)和卵巢基质,各6个重复组,其中1个小卵泡样本包含8~10个小卵泡,1个大卵泡样本仅包含1个大卵泡,连同分离后的卵巢基质放入冻存管,立即置于液氮中,带回实验室于−80 ℃冰箱保存,用于后续试验。试验所得的卵巢基质样品中有少量d<3 mm的小卵泡,所有d>1 cm的大卵泡已分离出来。
1.2 总RNA抽提、cDNA文库构建和转录组测序
将卵巢基质、大卵泡和小卵泡完全研磨,用Trizol试剂提取总RNA(Invitrogen,美国)。通过DanoDrop ND-2000分光度计(Thermo Science,美国)、10 g/L琼脂糖凝胶分别检测RNA的浓度和纯度、完整性。每个样品取3 μL总RNA,构建cDNA文库,利用Illumina HiSeq测序平台进行双末端测序。整个流程委托上海派森诺生物科技股份公司完成。
1.3 转录组数据分析
对测序数据采用Cutadapt软件去除3′端的接头,去除的部分与接头有至少10 bp重叠(AGATCGGAAG),允许20%碱基错配;去除平均占比低于Q20(碱基识别准确率在99%以上的碱基所占百分比)的Reads。通过Bowtie2软件(GCF_001704415.1_ARS1_genomic; http://www.ensembl.org/)建立参考基因组索引,使用Tophat2软件的microexon-search和library-type=fr-firststrand参数将过滤后的Reads比对到参考基因组上,默认Reads和参考基因组序列的碱基错配在2个之内即为比对成功。采用R语言DESeq程序包对差异基因进行筛选,条件为:|log2表达差异倍数|>1,显著性P<0.05,至少有4个样品的FPKM(每百万Reads中来自某一基因每千碱基长度的Reads数目,Fragments per kilobase per million of reads)大于1。通过R语言Pheatmap软件包对所有基因和样品进行双向聚类分析。
1.4 差异基因功能注释
利用Gene ontology(GO)数据库对所有差异基因进行注释,通过Kyoto encyclopedia of genes and genomes(KEGG)数据库对差异基因进行通路分析,采用超几何分布计算差异基因显著富集的GO条目和KEGG通路,P<0.05为显著富集。
1.5 qRT-PCR验证分析
从差异基因中随机选择5个基因,利用qRT-PCR进行转录组表达谱验证分析,PCR反应体系10 μL:5 μL SYBR® Green Master Mix,上下游引物各0.5 μL,1 μL cDNA和3 μL ddH2O。反应条件:95 ℃,1 min;95 ℃,30 s,58 ℃,30 s,72 ℃,1 min,34个循环。引物设计参考NCBI的山羊相关基因序列,β-actin作为内参基因,用Primer 5.0软件设计,并送Sangon Biotech合成引物(表1)。
表 1 用于qRT-PCR检测的基因引物Table 1. Gene primers for qRT-PCR detection基因
Gene引物序列(5′→3′)
Primer sequence扩增长度/bp
Product lengthCYP19A1 F: CCGAAGTTGTGCCTATTGC
R: GCTGGGACCTGGTATTGAG101 INHBA F: CATGTGGGAAAAGTGGGGGA
R: TCAAAGTGCAGCGTCTTCCT146 KITLG F: CATTTATCTTCAACTGCTCCTA
R: CCACCATCTCGCTTATCC191 SERP1 F: GTCTTCCTGTGCTCGCTCTTC
R: CGATTATGGCTTCCGATTTCA268 GPNMB F: CCTTGTCCTTTGCCTTCAC
R: AAATCCACCAGGGAGTCGT230 β-actin F: TGCTTCTAGGCGGACTGATT
R: TACAATCAAAGTCCTCGGCCAC106 1.6 数据统计分析
试验数据用SPSS 22.0统计软件分析,通过独立样本t检验来进行差异显著性分析。
2. 结果与分析
2.1 测序数据质控
将本研究所得的测序数据经过质量控制,对Clean reads进行碱基含量分布和碱基质量分布统计,Q30(碱基识别准确率在99.9%以上的碱基所占百分比)在91%以上,测序数据与山羊的数据库序列相似性在83%以上(表2)。综合表明测序数据质量好,满足后续分析的要求。
表 2 测序数据质控Table 2. Quality control of sequencing data样品1)
Sample有效测序量
No. of clean reads有效测序的占比/%
Ratio of clean readsQ302)/% 样品1)
Sample有效测序量
No. of clean reads有效测序的占比/%
Ratio of clean readsQ302)/% O1 105 953 656 99.68 94.28 LF1 104 768 836 99.62 94.23 O2 102 197 046 99.34 92.71 LF2 100 396 168 99.70 94.85 O3 105 678 158 99.41 92.86 LF3 104 879 676 99.48 94.15 O4 101 844 278 99.44 92.86 LF4 102 209 432 99.37 92.57 O5 107 096 856 99.32 91.31 LF5 100 346 902 99.75 93.86 O6 104 231 700 99.57 92.09 LF6 101 449 700 99.50 92.26 SF1 102 014 414 99.69 94.88 SF4 105 756 846 99.19 91.52 SF2 102 993 890 99.45 94.24 SF5 103 645 956 99.58 93.79 SF3 106 653 668 99.77 94.11 SF6 106 529 902 99.45 92.38 1) O:卵巢基质;LF:大卵泡;SF:小卵泡;2) 碱基识别准确率在99.9%以上的碱基所占百分比
1) O: Ovarian stroma; LF: Large follicle; SF: Small follicle; 2) Percentage of bases with base recognition accuracy above 99.9%2.2 差异基因筛选
测序结果分析表明,卵巢基质和大卵泡之间显著差异表达的基因总数为524个,其中显著上调的有238个,显著下调的有286个;卵巢基质和小卵泡之间显著差异表达的基因总数为180个,其中显著上调的有25个,显著下调的有155个;大卵泡和小卵泡之间显著差异表达的基因总数为403个,其中显著上调的有281个,显著下调的有122个。结合近年哺乳动物中与卵泡发育相关的基因的报道[6, 20-33],我们发现本试验中15个差异表达的基因可能与卵泡发育相关,包括INHA、INHBA、CYP19A1、MGST1、BEX2、APOA1、APOA2、MMP15、CITED1、PRSS23、LRP8、ADAMTS1、SPARCL1、TNFRSF1B和TNFRSF19。按照P值大小筛选排名前10的差异表达基因,再根据差异倍数从大到小进行排序[34-35],结果如表3所示。通过聚类图(图1)发现,卵巢基质、大卵泡和小卵泡各样本的基因分别聚类在一起,且同一组几个不同样本聚类在同一个簇中。此外,卵巢基质与小卵泡间差异基因的数量最少,且大部分基因表达量相似(图1),推测小卵泡和卵巢基质间的表达模式较为相似。大卵泡与卵巢基质、小卵泡的差异表达基因数量较多,且表达量相差较大,推测大卵泡与卵巢基质、小卵泡的基因表达模式差异较大。
![图 1 不同卵巢部位间差异表达基因的聚类图]() 图 1 不同卵巢部位间差异表达基因的聚类图O:卵巢基质;LF:大卵泡;SF:小卵泡;图例右侧的数字为基因表达上调或下调的倍数Figure 1. Cluster analysis of differentially expressed genes among different parts of ovaryO: Ovarian stroma; LF: Large follicle; SF: Small follicle; Numbers by the legend are expression fold-changes for up- or down-regulated genes表 3 不同卵巢部位间排名前10位的差异表达基因Table 3. Top ten differentially expressed genes among different parts of ovary
图 1 不同卵巢部位间差异表达基因的聚类图O:卵巢基质;LF:大卵泡;SF:小卵泡;图例右侧的数字为基因表达上调或下调的倍数Figure 1. Cluster analysis of differentially expressed genes among different parts of ovaryO: Ovarian stroma; LF: Large follicle; SF: Small follicle; Numbers by the legend are expression fold-changes for up- or down-regulated genes表 3 不同卵巢部位间排名前10位的差异表达基因Table 3. Top ten differentially expressed genes among different parts of ovary项目1)
Item基因
Genelog2(FC)2) P 项目1)
Item基因
Genelog2(FC)2) P 项目1)
Item基因
Genelog2(FC)2) P O vs LF EVI2A 1.69 3.45×10−6 O vs SF KCTD12 1.57 1.52×10−3 LF vs SF GLI1 2.13 1.22×10−6 IRF2BP2 1.24 5.84×10−7 CFD 1.43 3.02×10−4 SLC25A34 1.57 5.72×10−6 PNRC1 1.16 8.06×10−6 PTCH2 −2.23 2.59×10−3 PRRT2 1.35 2.55×10−5 GPR21 1.06 6.22×10−6 GDF5 −1.84 1.99×10−3 PPM1J 1.34 3.08×10−5 TMEM176B −1.86 1.27×10−5 GLI1 −1.48 3.80×10−4 TNRC6C 1.24 2.21×10−5 RDH11 −1.50 1.84×10−6 EMID1 −1.32 4.01×10−4 EFNB3 1.10 2.00×10−5 MINOS1 −1.37 6.17×10−6 MYCL −1.30 3.35×10−4 EFS 1.07 3.58×10−5 MPDU1 −1.09 1.09×10−5 DPEP3 −1.26 1.47×10−3 ADGRL1 1.04 1.63×10−5 NME1 −1.06 1.01×10−5 DPEP2 −1.23 1.68×10−3 KITLG −1.85 1.34×10−5 ARL2BP −1.05 3.12×10−6 CNRIP1 −1.20 1.81×10−3 TMEM176B −1.49 3.67×10−5 1) O:卵巢基质;LF:大卵泡;SF:小卵泡;2) FC:表达差异倍数
1) O: Ovarian stroma; LF: Large follicle; SF: Small follicle; 2) FC: Expression fold-change2.3 共有的差异表达基因
在卵巢基质vs大卵泡、卵巢基质vs小卵泡、小卵泡vs大卵泡这3组中有4个共有的差异表达基因;卵巢基质vs大卵泡、卵巢基质vs小卵泡之间有73个共有的差异表达基因;卵巢基质vs大卵泡、小卵泡vs大卵泡之间有186个共有的差异表达基因;卵巢基质vs小卵泡、小卵泡vs大卵泡之间有29个共有的差异表达基因。
2.4 差异表达基因的生物学功能分析
基于GO数据库,对差异基因进行功能分类,各功能分类下差异前10位的GO条目如表4、表5和表6所示。在卵巢基质vs大卵泡(表4)中有15个GO条目显著富集(P<0.05),包括催化活性、细胞质、细胞膜、细胞内膜结合细胞器、转移酶活性等。在卵巢基质vs小卵泡(表5)中有25个GO条目显著富集(P<0.05),包括小分子结合、碳水化合物衍生物结合、核苷酸结合、磷酸核苷结合、嘌呤核苷三磷酸结合等。在小卵泡vs大卵泡(表6)中有64个GO条目显著富集(P<0.05),包括细胞过程、细胞内部、线粒体部分、嘌呤核苷酸结合等。
表 4 卵巢基质vs大卵泡部分差异表达基因的GO功能分类Table 4. GO functional classification of partial differentially expressed genes in ovarian stroma vs large follicle分类
Category条目
Term基因数目
Gene countP 生物学过程 Biological process RNA加工过程 RNA processing 5 2.70×10−2 细胞组分 Cellular component 细胞 Cell 66 1.90×10−2 细胞部分 Cell part 65 2.40×10−2 细胞内部 Intracellular 63 1.70×10−3 细胞内的部分 Intracellular part 55 6.00×10−3 细胞质 Cytoplasm 39 9.80×10−4 细胞内膜结合细胞器 Intracellular membrane-bounded organelle 36 2.50×10−2 膜结合细胞器 Membrane-bounded organelle 36 2.50×10−2 细胞质部分 Cytoplasmic part 28 2.80×10−2 分子功能 Molecular function 催化活性 Catalytic activity 51 8.20×10−4 磷酸核苷结合 Nucleotide phosphate binding 22 4.60×10−2 核苷酸结合 Nucleotide binding 22 4.60×10−2 转移酶活性 Transferase activity 21 1.50×10−2 转移酶活性(转移含磷基团)
Transferase activity (Transferring phosphorus-containing groups)12 2.70×10−2 异构酶活性 Isomerase activity 5 2.00×10−2 表 5 卵巢基质vs小卵泡部分差异表达基因的GO功能分类Table 5. GO functional classification of partial differentially expressed genes in ovarian stroma vs small follicle分类
Category条目
Term基因数目
Gene countP 生物学过程 Biological process 初级代谢过程 Primary metabolic process 13 4.70×10−2 细胞对应激的反应 Cellular response to stress 4 4.40×10−2 分子功能 Molecular function 小分子结合 Small molecule binding 12 2.30×10−4 碳水化合物衍生物结合 Carbohydrate derivative binding 11 2.50×10−4 核苷酸结合 Nucleotide binding 11 5.90×10−4 磷酸核苷结合 Nucleoside phosphate binding 11 5.90×10−4 核糖核苷结合 Ribonucleoside binding 10 6.00×10−4 核苷结合 Nucleoside binding 10 6.00×10−4 嘌呤核苷三磷酸结合 Purine ribonucleoside triphosphate binding 10 6.00×10−4 嘌呤核苷结合 Purine nucleoside binding 10 6.00×10−4 嘌呤核糖核苷结合 Purine ribonucleoside binding 10 6.00×10−4 核糖核苷酸结合 Ribonucleotide binding 10 6.70×10−4 表 6 小卵泡vs大卵泡部分差异表达基因的GO功能分类Table 6. GO functional classification of partial differentially expressed genes in small follicle vs large follicle分类
Category条目
Term基因数目
Gene countP 生物学过程
Biological process细胞进程 Cellular process 73 3.40×10−2 有机环状化合物代谢过程 Organic cyclic compound metabolic process 31 1.70×10−2 有机氮化合物代谢过程 Organonitrogen compound metabolic process 17 3.80×10−2 有机磷代谢过程 Organophosphate metabolic process 10 1.60×10−2 含核苷的小分子代谢过程 Nucleobase-containing small molecule metabolic process 9 3.50×10−2 嘌呤核糖核苷代谢过程 Purine ribonucleoside metabolic process 8 2.00×10−2 嘌呤核苷代谢过程 Purine nucleoside metabolic process 8 2.00×10−2 核糖核苷代谢过程 Ribonucleoside metabolic process 8 2.00×10−2 糖基化合物代谢过程 Glycosyl compound metabolic process 8 2.90×10−2 核苷代谢过程 Nucleoside metabolic process 8 2.90×10−2 细胞组分
Cellular component细胞内 Intracellular 64 2.70×10−3 细胞质 Cytoplasm 35 2.80×10−2 细胞质部分 Cytoplasmic part 29 2.30×10−2 线粒体 Mitochondrion 14 9.20×10−4 线粒体部分 Mitochondrial part 10 5.20×10−3 细胞器膜 Organelle membrane 10 2.30×10−2 线粒体膜 Mitochondrial membrane 9 5.70×10−3 细胞器包膜 Organelle envelope 9 9.00×10−3 线粒体包膜 Mitochondrial envelope 9 9.00×10−3 囊膜 Envelope 9 9.00×10−3 分子功能
Molecular function磷酸核苷结合 Nucleoside phosphate binding 28 2.90×10−3 核苷酸结合 Nucleotide binding 28 2.90×10−3 嘌呤核苷酸结合 Purine nucleotide binding 25 2.10×10−3 嘌呤核苷结合 Purine nucleoside binding 24 3.30×10−3 嘌呤核苷三磷酸结合 Purine ribonucleoside triphosphate binding 24 3.30×10−3 嘌呤核糖核苷结合 Purine ribonucleoside binding 24 3.30×10−3 核苷结合 Nucleoside binding 24 3.30×10−3 核糖核苷结合 Ribonucleoside binding 24 3.30×10−3 嘌呤核糖核苷酸结合 Purine ribonucleotide binding 24 4.10×10−3 基于KEGG数据库的通路富集分析(各组对比项目中前10位的信号通路见表7),在卵巢基质vs大卵泡中差异表达基因涉及55条通路,显著富集的通路有46个(P<0.05),包括氧化磷酸化、代谢途径、内质网蛋白质加工、柠檬酸循环、类固醇生物合成等。在卵巢基质vs小卵泡中差异表达基因涉及27条通路,显著富集的通路有17个(P<0.05),包括细胞周期、错配修复、DNA复制、卵母细胞减数分裂、RNA转运、孕酮介导的卵母细胞成熟等。在小卵泡vs大卵泡中差异表达基因涉及42条通路,显著富集的通路有30个(P<0.05),包括氧化磷酸化、蛋白酶体、内质网蛋白质加工等。
表 7 卵巢基质、大卵泡和小卵泡部分差异表达基因的KEGG代谢通路Table 7. KEGG metabolic pathway of partial differentially expressed genes in ovarian stroma, large follicle and small follicle项目1)
Item通路
Pathway数目
CountP O vs LF 代谢途径 Metabolic pathway 298 4.90×10−17 抗生素的生物合成 Antibiotic biosynthesis 79 1.30×10−14 氧化磷酸化 Oxidative phosphorylation 77 1.10×10−22 内质网蛋白质加工 Protein processing in endoplasmic reticulum 70 1.00×10−14 泛素介导的蛋白水解 Ubiquitin mediated proteolysis 34 6.30×10−3 细胞周期 Cell cycle 32 7.30×10−3 甲状腺激素信号通路 Thyroid hormone signaling pathway 29 5.00×10−3 柠檬酸循环 Citrate cycle 17 2.20×10−6 Notch信号通路 Notch signaling pathway 13 4.90×10−2 类固醇生物合成 Steroid biosynthesis 12 1.90×10−4 O vs SF 癌症的途径 Pathway in cancer 28 3.20×10−2 细胞周期 Cell cycle 25 4.50×10−9 抗生素的生物合成 Antibiotic biosynthesis 17 4.00×10−2 RNA转运 RNA transport 16 1.10×10−2 嘌呤代谢 Purine metabolism 15 3.70×10−2 卵母细胞减数分裂 Oocyte meiosis 13 8.30×10−3 嘧啶代谢 Pyrimidine metabolism 11 1.30×10−2 DNA复制 DNA replication 9 2.30×10−4 孕酮介导的卵母细胞成熟 Progesterone-mediated oocyte maturation 9 4.30×10−2 错配修复 Mismatch repair 8 8.90×10−5 SF vs LF 代谢途径 Metabolic pathway 248 5.60×10−4 氧化磷酸化 Oxidative phosphorylation 74 3.80×10−19 内质网蛋白质加工 Protein processing in endoplasmic reticulum 67 6.00×10−12 抗生素的生物合成 Antibiotic biosynthesis 54 7.80×10−4 泛素介导的蛋白水解 Ubiquitin mediated proteolysis 40 2.90×10−4 甲状腺激素信号通路 Thyroid hormone signaling pathway 32 1.40×10−3 AMPK信号通路 AMPK signaling pathway 32 8.70×10−3 蛋白酶体 Proteasome 31 1.10×10−12 Notch信号通路 Notch signaling pathway 15 1.50×10−2 类固醇生物合成 Steroid biosynthesis 9 2.10×10−2 1) O:卵巢基质;LF:大卵泡;SF:小卵泡
1) O: Ovarian stroma; LF: Large follicle; SF: Small follicle2.5 基因筛选及qRT-PCR验证分析
从3组差异基因中筛选5个基因,分别是CYP19A1和INHBA(卵巢基质vs大卵泡)、GPNMB(卵巢基质vs小卵泡)、KITLG和SFRP1(小卵泡vs大卵泡),进行qRT-PCR验证(图2)。这些基因的表达变化趋势与测序结果基本一致,说明测序结果可靠。
![图 2 不同卵巢部位间差异表达基因的qRT-PCR验证]() 图 2 不同卵巢部位间差异表达基因的qRT-PCR验证Figure 2. Quantitative identification of differentially expressed genes among different parts of ovary by qRT-PCR
图 2 不同卵巢部位间差异表达基因的qRT-PCR验证Figure 2. Quantitative identification of differentially expressed genes among different parts of ovary by qRT-PCR3. 讨论
山羊性成熟时,以卵泡波的形式募集卵泡,并使卵泡发育至成熟排卵[36]。在形成有腔卵泡之前,卵泡的发育不受生殖激素调控,发育比较缓慢;在形成腔体时,性腺轴会分泌激素,促进卵泡发育或闭锁[37-38]。山羊是单胎或双胎动物[3],与人类相似,研究卵泡不同发育阶段的作用机制,山羊是较好的动物模型。
本研究采用高通量测序技术,首次对川中黑山羊的卵巢基质、大卵泡和小卵泡的mRNA表达量进行研究。结果表明卵巢基质、大卵泡和小卵泡的表达模式不同,其中卵巢基质和小卵泡的表达模式更相近,可能是因为在分离卵泡时,每个卵巢基质中d>1 cm的大卵泡已分离出来,但不可避免的,卵巢基质中含有少量的d<3 mm的小卵泡,从而造成卵巢基质和小卵泡的基因表达模式更相近。
通过差异基因分析,筛选出与其他哺乳动物卵泡发育或闭锁有关的基因,包括在大卵泡上调的抑制素基因家族、细胞色素P450家族19亚家族成员1(CYP19A1)、MGST1、BEX家族、APO家族、MMP家族、CITED家族、PRSS23、LRP8;在大卵泡下调的ADAMTS1、SPARCL1、TNFRSF家族。在猪[6]和牛[8, 11]的研究中,抑制素基因家族在闭锁卵泡或小卵泡中低表达,通过负反馈调节抑制垂体分泌促卵泡激素,影响卵泡闭锁。INHA和INHBA在牛大卵泡上调[10],有助于卵泡膜产生雄激素并抑制垂体产生卵泡刺激素。CYP19A1的过表达增加大窦状卵泡类固醇合成[6-8],促进卵泡发育。谷胱甘肽代谢MGST1与细胞的代谢和解毒能力有关,其高表达可能表示健康卵泡的解毒能力更好[39]。BEX家族是X染色体连锁基因家族,BEX2在小卵泡低表达,引起线粒体凋亡,使细胞周期阻滞于G1期,从而调控颗粒细胞增殖和凋亡[9, 40-41]。载脂蛋白APOA1在中卵泡中高表达,可能参与了卵泡的选择及发育过程[11]。基质金属蛋白酶(Matrix metallopeptidase,MMP)家族是一类依赖金属锌离子的蛋白水解酶家族,能有效地降解细胞外基质,可能与卵泡体积的快速增大有关[11]。ADAMTS1基因家族与膜外结构域的分裂有关,在小卵泡中表达上调[9],在经产奶牛排卵前的卵泡中上调[42]。SPARCL1是SPARC家族一员,在小卵泡中高表达,阻滞细胞停留在G1期,抑制细胞增殖[9, 43]。但IGFBP5、IGF1、MMP9、PGRMC1等基因的表达量与其他研究报道的不一致。值得注意的是,在本研究中经典颗粒细胞凋亡标志物TNF受体超家族(TNFRSF1B、TNFRSF19)在大卵泡中下调[8],与一些研究报道的无显著差异不同[6-7]。
进一步对差异基因功能分析发现,卵巢基质vs大卵泡、卵巢基质vs小卵泡、小卵泡vs大卵泡三者富集的通路大不相同,其中,卵巢基质vs大卵泡、小卵泡vs大卵泡富集的通路较多,且有大部分是相同的;而卵巢基质vs小卵泡富集的通路较少,与其他2组差异较大。说明可能是大卵泡的生物学功能较为丰富。功能富集表明不同组织表现出不同的富集特征,核苷酸结合和磷酸核苷结合过程在3组中都共同富集,但在卵巢基质vs大卵泡中,特有的GO条目包括RNA加工过程、细胞质、细胞内膜、酶的催化活性等,可能是因为大卵泡阶段代谢过程比较旺盛,大卵泡与卵巢基质之间物质运输比较频繁。在卵巢基质vs小卵泡中特有的GO条目有细胞应激反应、初级代谢过程等,可能是因为小卵泡正处于初级发育阶段。在小卵泡vs大卵泡中特有的GO条目有线粒体和线粒体膜、细胞器内膜等,可能是因为卵泡发育过程主要受到线粒体膜的调控。
与卵巢基质相比,柠檬酸循环和类固醇生物合成信号通路在大卵泡中上调。柠檬酸循环保证机体能量供给和促进物质代谢[44]。类固醇生物合成在卵泡发育中起着重要的作用[45],例如类固醇合成快速调节蛋白(Steroidogenic acute regulatory protein,STAR)可介导胆固醇进入线粒体内膜,调控孕酮和雌激素的合成[46-48],影响卵泡发育;健康卵泡中CYP19A1的上调,促进孕烯醇酮形成,孕烯醇酮是黄体酮(Progesterone,P4)和雌二醇(Estradiol,E2)的底物,可能是促进卵泡发育的主要因素[7]。
在卵巢基质vs小卵泡中,差异基因富集的信号通路与细胞分裂周期有关。有研究[49-50]报道,在小鼠卵泡发育阶段,细胞周期、DNA复制和减数分裂等多个信号通路被激活,与本试验结果一致,可能在小卵泡阶段,卵母细胞发育上调了与细胞分裂相关的信号通路。
在小卵泡vs大卵泡中,差异基因富集的甲状腺激素信号通路、内质网蛋白质加工、AMPK信号通路、类固醇生物合成和Notch信号通路在猪[7]和牛[10]的卵泡转录组研究中均有报道。内质网蛋白质加工和甲状腺激素信号通路的显著富集,可能是因为随着卵泡发育,颗粒细胞分化成熟,甾体激素合成逐渐增强[51-52],物质代谢也因此增强。在人和大鼠的研究中发现[53],类固醇激素(如孕酮)与不同膜受体结合,抑制颗粒细胞的有丝分裂或凋亡,从而抑制卵泡发育或闭锁,维持卵泡的平衡状态。关于绵羊的研究发现,Notch信号通路调节细胞增殖、分化和凋亡,参与卵巢基质卵泡的发育[54]。
4. 结论
卵巢基质、大卵泡、小卵泡的差异mRNA表达模式不相同,其中卵巢基质与小卵泡更相近。就卵泡发育而言,深入研究这些特异性mRNA在卵泡不同发育阶段的作用有着重要意义。本文从生物信息学角度分析卵巢不同组织表达谱,为下一步差异基因的功能验证及其在卵泡发育中的作用提供一定的理论基础。
致谢:感谢练志全帮助分离小卵泡和大卵泡,感谢赵智锋帮助分析转录组测序结果!
-
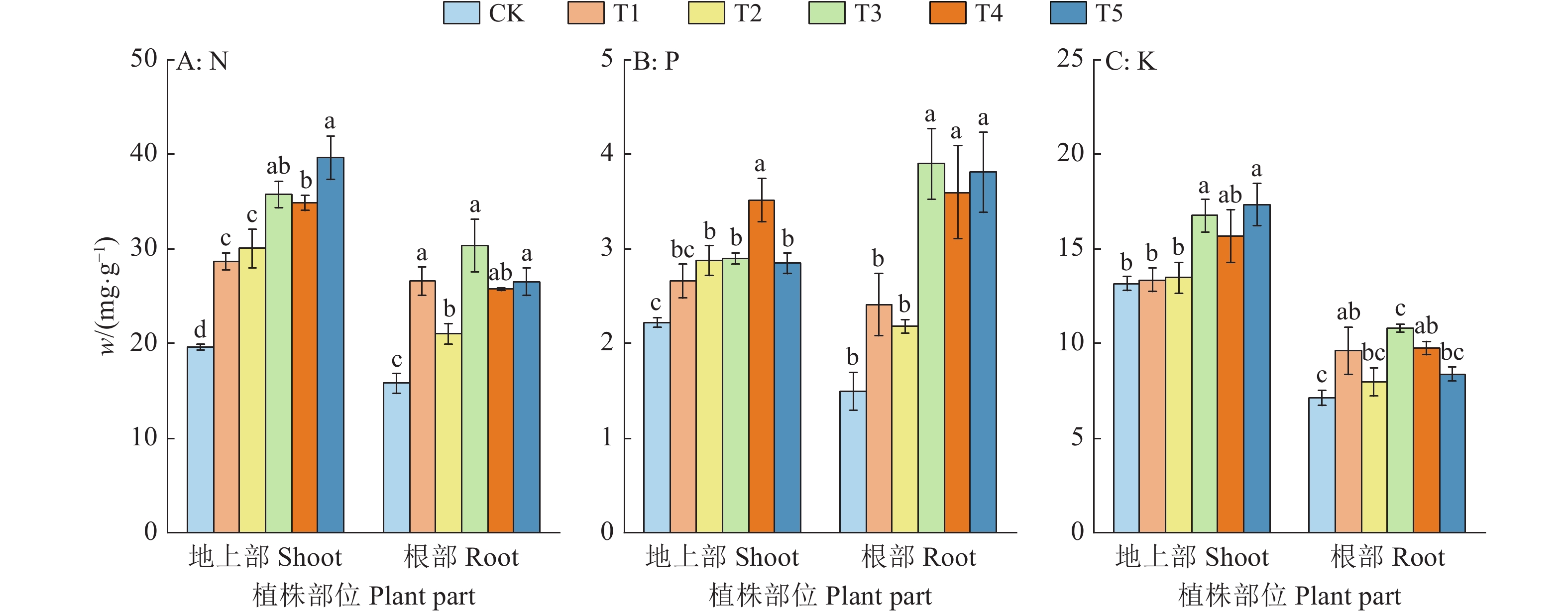
![]()
图 1 复合肥不同施用量对巴戟天不同部位N、P、K含量的影响
各小图中,相同部位柱子上方的不同小写字母表示处理间差异显著(P<0.05,Duncan’s法)
Figure 1. Effects of different application amounts of compound fertilizer on contents of N, P and K in different parts of Morinda officinalis
In each figure, different lowercase letters on the columns of the same part indicate significant differences among different treatments (P<0.05, Duncan’s method)
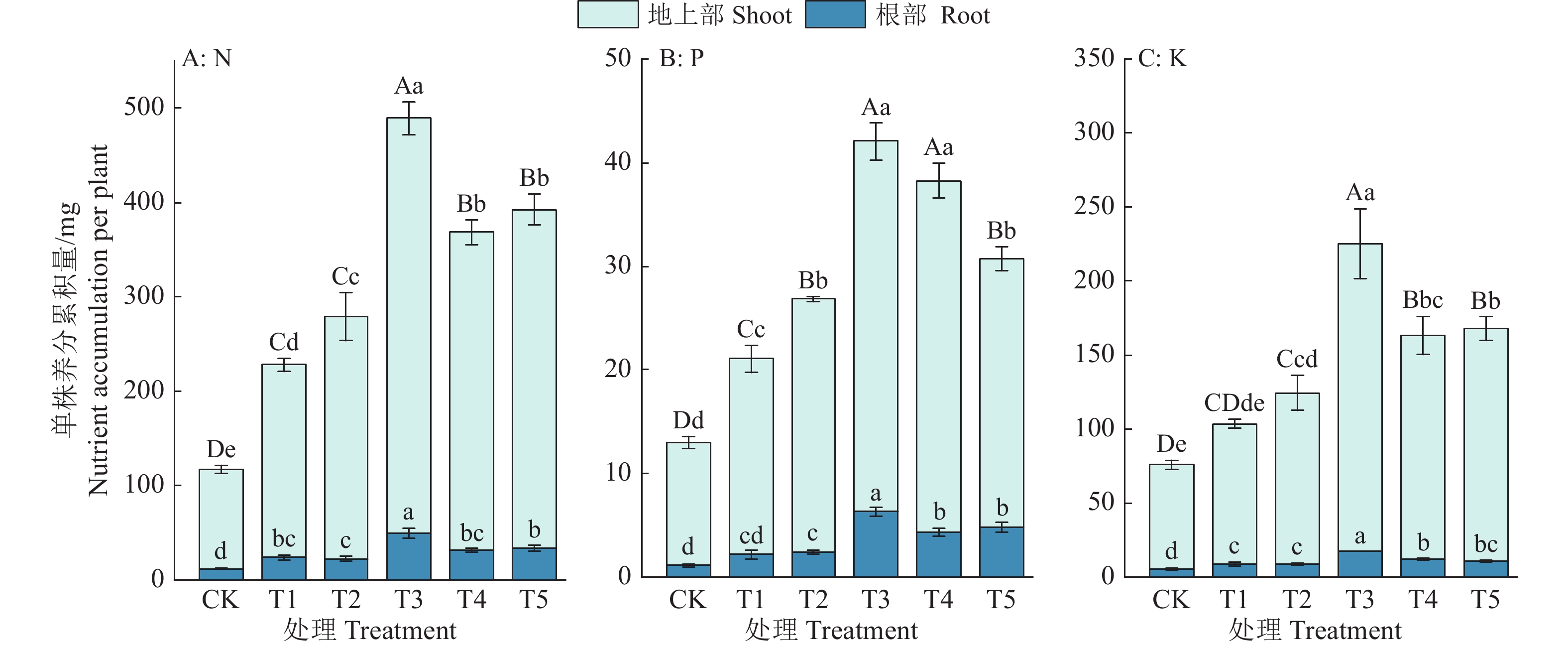
![]()
图 2 复合肥不同施用量对巴戟天各部位N、P、K单株累积量的影响
各小图中,不同小写字母表示处理间地上部、根部养分累积量差异显著,不同大写字母表示处理间全株养分累积量差异显著(P<0.05,Duncan’s法)
Figure 2. Effects of different application amounts of compound fertilizer on the accumulation of N, P and K per plant in different parts of Morinda officinalis
In each figure, different lowercase letters on the columns indicate significant differences in aboveground and root nutrient accumulation among treatments, different capital letters indicate significant differences in whole plant nutrient accumulation among treatments (P < 0.05, Duncan’s method)
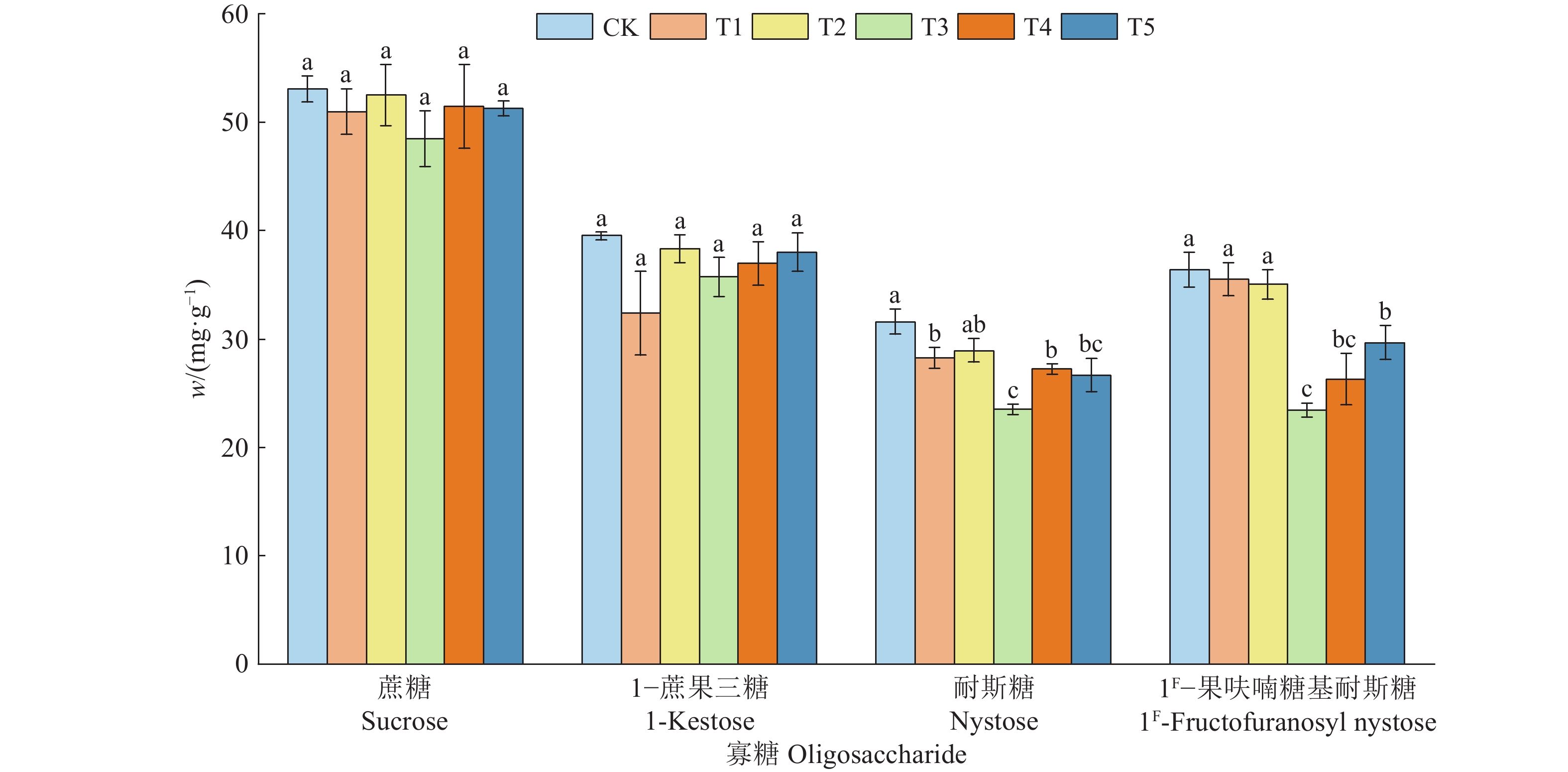
![]()
图 3 复合肥不同施用量对巴戟天4种寡糖含量的影响
相同寡糖柱子上方的不同小写字母表示处理间差异显著(P<0.05,Duncan’s法)
Figure 3. Effects of different application amounts of compound fertilizer on contents of four oligosaccharides in Morinda officinalis
Different lowercase letters on the columns of the same oligosaccharide indicate significant differences among treatments (P<0.05, Duncan’s method)
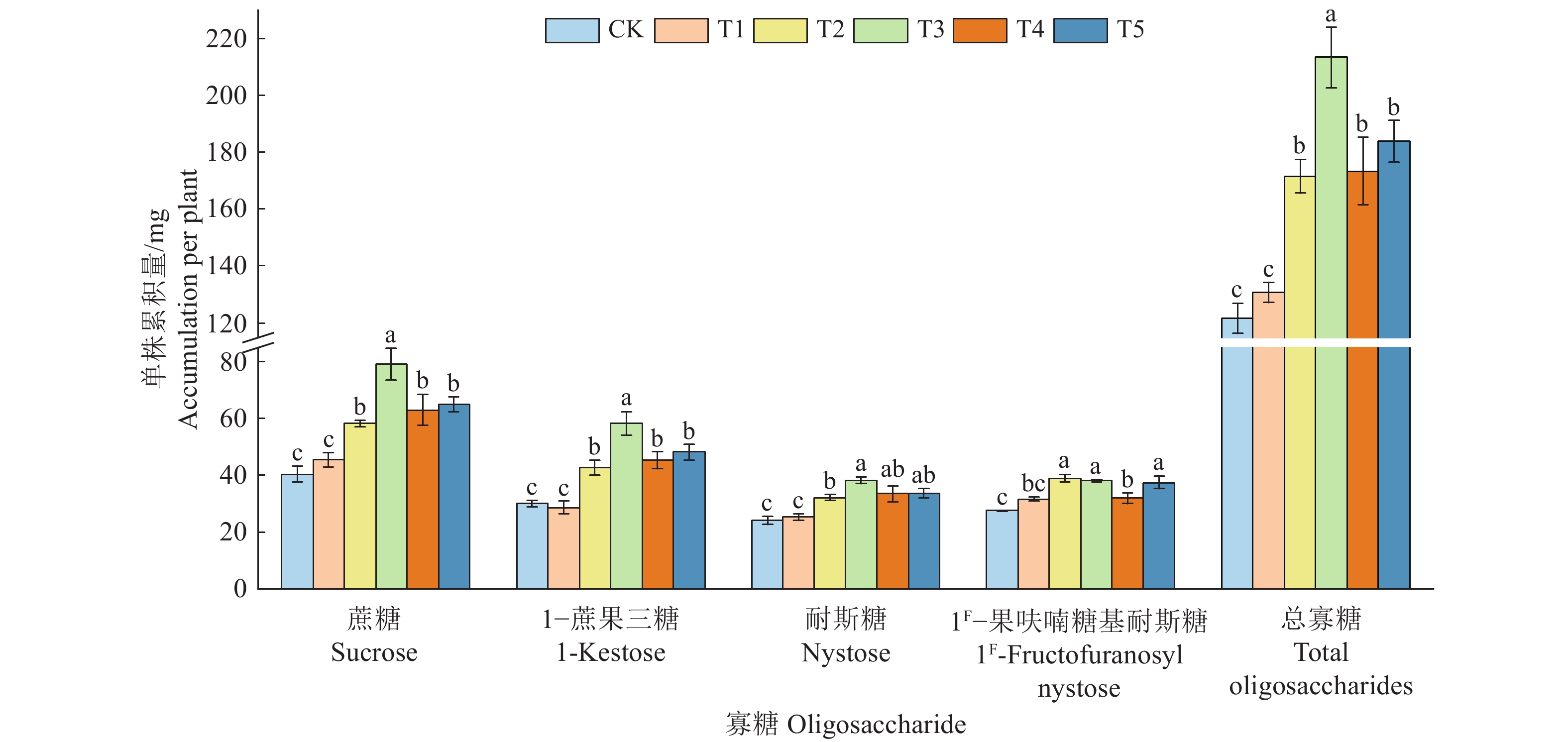
![]()
图 4 复合肥不同施用量对巴戟天4种寡糖单株累积量的影响
相同寡糖柱子上方的不同小写字母表示处理间差异显著(P<0.05,Duncan’s法)
Figure 4. Effects of different application amounts of compound fertilizer on accumulation of four oligosaccharides in Morinda officinalis
Different lowercase letters on the columns of the same oligosaccharide indicate significant differences among treatments (P<0.05, Duncan’s method)
![]()
图 5 巴戟天不同指标的相关性分析
“*”和“**”分别表示在P<0.05和P<0.01水平显著相关(Pearson法)
Figure 5. Correlation analyses of different indicators for Morinda officinalis
“*” and “**” indicate significant correlations at P < 0.05 and P < 0.01 levels, respectively (Pearson method)
![]()
图 6 复合肥不同施用量的主成分分析
PC1:第一主成分,PC2:第二主成分
Figure 6. Principal component analyses of different application amounts of compound fertilizer
PC1: The first principal component, PC2: The second principal component
表 1 复合肥不同施用量对巴戟天生长指标的影响1)
Table 1 Effects of different application amounts of compound fertilizer on growth indexes of Morinda officinalis
处理
Treatment地径/mm
Ground diameter总生物量/g
Total biomass根部产量/g
Root yieldCK 5.19±0.35a 6.12±0.31d 0.76±0.04c T1 5.25±0.14a 8.00±0.14c 0.89±0.05c T2 6.32±0.54a 9.65±0.61bc 1.11±0.08b T3 6.07±0.31a 13.96±0.78a 1.63±0.04a T4 6.22±0.44a 10.88±0.24b 1.22±0.08b T5 5.72±0.25a 10.40±0.82b 1.26±0.05b 1)同列数据后的不同小写字母表示处理间差异显著(P<0.05,Duncan’s法)
1) Different lowercase letters in the same column indicate significant differences among different treatments (P<0.05, Duncan’s method) 下载: 导出CSV
下载: 导出CSV
表 2 复合肥不同施用量的肥料利用效率1)
Table 2 Fertilizer utilization efficiency of different application amounts of compound fertilizer
% 处理 Treatment N P K T1 36.97±1.37a 2.69±0.31ab 9.16±0.53b T2 27.14±4.41b 2.31±0.07b 8.04±2.05b T3 41.35±2.37a 3.23±0.20a 16.62±2.63a T4 20.97±1.29bc 2.11±0.17b 7.27±1.00b T5 18.38±1.20c 1.18±0.10c 6.11±0.56b 1)同列数据后的不同小写字母表示处理间差异显著(P<0.05,Duncan’s法)
1) Different lowercase letters in the same column indicate significant differences among treatments (P<0.05, Duncan’s method)
下载: 导出CSV
表 3 主成分分析的载荷系数
Table 3 Loading coefficients for principal component analysis
评价指标
Evaluation
index第一主成分
The first
principal
component第二主成分
The second
principal
component总生物量 Total biomass 0.952 −0.083 产量 Yield 0.959 −0.154 全株N含量
Whole plant N content0.843 0.353 全株P含量
Whole plant P content0.708 0.475 全株K含量
Whole plant K content0.961 −0.141 蔗糖积累量
Sucrose accumulation0.946 −0.189 1−蔗果三糖积累量
1-Kestose accumulation0.935 −0.232 耐斯糖积累量
Nystose accumulation0.755 −0.372 1F−果呋喃糖基耐斯糖累积量
1F-Fructofuranosyl nystose accumulation0.618 0.655 特征值 Eigenvalue 6.682 1.057 方差贡献率/%
Contribution of variance74.246 11.748 累计贡献率/%
Cumulative contribution rate74.246 85.994
下载: 导出CSV
表 4 复合肥不同施用量的主成分分析综合得分
Table 4 Comprehensive score of principal component analysis of different application amounts of compound fertilizer
处理
Treatment主成分分值
Principal component value综合得分(F)
Comprehensive
score排序
RankingF1 F2 CK −3.92 −0.49 −2.96 6 T1 −2.28 0.27 −1.66 5 T2 −0.01 −0.86 −0.11 4 T3 3.48 −0.52 2.53 1 T4 1.07 1.17 0.93 3 T5 1.65 0.42 1.27 2
下载: 导出CSV
-
[1] 国家药典委员会. 中华人民共和国药典: 1部[M]. 北京: 中国医药科技出版社, 2020: 83. [2] YANG Z, HU J, ZHAO M. Isolation and quantitative determination of inulin-type oligosaccharides in roots of Morinda officinalis[J]. Carbohydrate Polymers, 2011, 83(4): 1997-2004. doi: 10.1016/j.carbpol.2010.11.006
[3] CHEN D, LI N, LIN L, et al. Confocal mirco-Raman spectroscopic analysis of the antioxidant protection mechanism of the oligosaccharides extracted from Morinda officinalis on human sperm DNA[J]. Journal of Ethnopharmacology, 2014, 153(1): 119-124. doi: 10.1016/j.jep.2014.01.021
[4] YANG X, CHEN D, CHEN T, et al. Oligosaccharides from Morinda officinalis slow the progress of aging mice by regulating the key microbiota-metabolite pairs[J]. Evidence-Based Complementary and Alternative Medicine, 2019, 2019: 9306834. doi: 10.1155/2019/9306834.
[5] ZHU J, PENG Q, XU Y, et al. Morinda officinalis oligosaccharides ameliorate depressive-like behaviors in poststroke rats through upregulating GLUT3 to improve synaptic activity[J]. FASEB Journal, 2020, 34(10): 13376-13395. doi: 10.1096/fj.201902546RR
[6] 葛阳, 康传志, 万修福, 等. 生产中氮肥施用及其对中药材产量和质量的影响[J]. 中国中药杂志, 2021, 46(8): 1883-1892. [7] 刘哲, 钟海蓉, 威则日沙, 等. 氮、磷、钾配施对华重楼根茎的产量和有效成分含量的影响[J]. 中草药, 2019, 50(24): 6103-6113. [8] 邹庆军, 汪涛, 郭巧生, 等. N、P、K调控对野菊药用部位性状及活性成分含量的影响[J]. 中草药, 2019, 50(2): 535-540. doi: 10.7501/j.issn.0253-2670.2019.02.036 [9] 苏文华, 张光飞, 王泽明, 等. 氮、磷和钾肥对灯盏花生长和有效成分积累的影响[J]. 中草药, 2009, 40(12): 1963-1966. [10] 苏现明, 王洪庆, 陈若芸, 等. 巴戟天属植物化学成分及药理活性研究进展[J]. 中药材, 2017, 40(4): 986-991. [11] LIANG J, LIANG J, HAO H, et al. The extracts of Morinda officinalis and its hairy roots attenuate dextran sodium sulfate-induced chronic ulcerative colitis in mice by regulating inflammation and lymphocyte apoptosis[J]. Frontiers in Immunology, 2017, 8: 905. doi: 10.3389/fimmu.2017.00905.
[12] 代文豪. 发酵法炮制巴戟天工艺优化及其活性成分变化研究[D]. 广州: 华南农业大学, 2018. [13] 李倩. 巴戟天炮制过程中化学成分变化规律的研究[D]. 广州: 广州中医药大学, 2015. [14] 朱露林, 沈燚, 易晓薇, 等. 不同产地及种植年限巴戟天中6种寡糖类成分的定量分析[J]. 中草药, 2021, 52(14): 4348-4352. [15] 冼丽铧, 吴道铭, 隆曼迪, 等. 土壤养分、种植年限和种植方式对巴戟天寡糖含量的影响[J]. 华南农业大学学报, 2021, 42(3): 75-85. [16] 邵玲, 李美映, 黎学荣, 等. 肇庆巴戟天两种种植模式的植物学性状及药效成分研究[J]. 中药材, 2019, 42(11): 2480-2485. [17] 隆曼迪. 种植环境及栽培管理措施对巴戟天生长和品质的影响[D]. 广州: 华南农业大学, 2019. [18] 吴礼树. 土壤肥料学[M]. 北京: 中国农业出版社, 2011. [19] 邵镪钎, 李丹, 蒋攀, 等. 氮、磷、钾配施对川明参产量和品质的影响[J]. 中草药, 2018, 49(16): 3926-3932. doi: 10.7501/j.issn.0253-2670.2018.16.029 [20] 梁琴, 陈兴福, 李瑶, 等. 化肥与有机肥配施对川芎产量的影响[J]. 中药材, 2015, 38(10): 2015-2020. [21] 陈晓玉, 贺超, 闫滨滨, 等. N、P、K配施对丹参产量和质量的影响[J]. 中草药, 2019, 50(3): 722-730. [22] 范巧佳, 刘灵, 郑顺林, 等. 春季施氮时期和数量对川芎产量、生物碱和阿魏酸的影响[J]. 四川农业大学学报, 2013, 31(2): 136-139. [23] 贾袭伟, 晋小军, 李慧, 等. 磷酸二氢钾施肥量对款冬生长及产量、质量的影响[J]. 中成药, 2022, 44(1): 314-317. [24] DIMKPA C O, FUGICE J, SINGH U, et al. Development of fertilizers for enhanced nitrogen use efficiency: Trends and perspectives[J]. Science of the Total Environment, 2020, 731: 139113. doi: 10.1016/j.scitotenv.2020.139113.
[25] 洪丕征, 刘世荣, 王晖, 等. 正常和环割条件下不同形态氮素添加对红椎幼苗光合特性的影响[J]. 生态学报, 2016, 36(14): 4485-4495. [26] 潘超美, 詹若挺, 丁平, 等. 不同有机肥对巴戟天生长及矿质养分吸收的影响[J]. 中药材, 2002, 25(10): 699-701. doi: 10.3321/j.issn:1001-4454.2002.10.002 [27] 漆小雪, 韦霄, 陈宗游, 等. 黄花蒿干物质的积累及青蒿素与N、P、K量的动态变化研究[J]. 中草药, 2011, 42(12): 2541-2544. [28] 董娟娥, 张康健, 梁宗锁. 植物次生代谢与调控[M]. 杨凌: 西北农林科技大学出版社, 2009. [29] 冯嘉仪, 谢姗宴, 吴道铭, 等. 氮磷钾配施对银杏果实和外种皮产量及品质的影响[J]. 生态学杂志, 2021, 40(6): 1650-1659. [30] 胡佳栋, 毛歌, 张志伟, 等. 不同施肥处理对党参产量和次生代谢物含量的影响研究[J]. 中国中药杂志, 2017, 42(15): 2946-2953. [31] 卢挺, 杨全, 唐晓敏, 等. 氮磷钾配比施肥对广金钱草产量及质量的影响[J]. 广西植物, 2014, 34(3): 426-430. [32] 吴向维. 基于巴戟天寡糖的喘可治注射剂质量评价研究[D]. 广州: 广州中医药大学, 2014. [33] 邓秋林, 杨正明, 陈雨, 等. 氮磷钾配施对瓦布贝母产量及总生物碱质量分数的影响[J]. 西北农业学报, 2019, 28(7): 1138-1146. [34] 鲁泽刚, 朱永全, 卢迎春, 等. 氮、磷、钾施用对灯盏花产量和主要提取物收获量的影响[J]. 植物科学学报, 2019, 37(1): 55-62.