
Construction of LMH cell line overexpressing VIM gene and effect on replication efficiency of GAstV
-
摘要:目的
构建稳定过表达波形蛋白(Vimentin,VIM)基因的鸡肝癌(Leghorn male hepatoma,LMH)细胞系以提高鹅星状病毒(Goose astrovirus,GAstV)增殖效率。
方法从鹅源细胞中通过PCR扩增VIM基因,结合同源重组的方法将其连接至慢病毒基因过表达系统载体pLV-sfGFP (2A) Puro,获得重组慢病毒质粒pLV-sfGFP (2A) Puro-VIM-Flag(pLV-VIM);利用293T细胞包装重组慢病毒颗粒,随后感染LMH细胞,并利用嘌呤霉素进行筛选以获得目标细胞;通过Western blot、RT-qPCR、间接免疫荧光试验和TCID50测定等方法评估VIM对GAstV增殖效率的影响;最后通过抗体阻断试验分析细胞表面VIM对GAstV侵入的影响。
结果经测序和酶切鉴定,成功构建了重组慢病毒载体pLV-VIM;经筛选后获得了过表达VIM基因的LMH细胞系(LMH-VIM)及其对照组细胞系(LMH-NC)。Western blot结果显示,LMH-VIM细胞中波形蛋白的表达水平显著高于LMH-NC。接种病毒后发现,过表达VIM显著上调LMH-VIM细胞中GAstV的mRNA和蛋白表达水平,以及细胞上清液中的病毒滴度。抗体阻断试验表明,细胞表面VIM可促进GAstV侵入细胞。
结论本研究为提高GAstV在体外细胞培养的增殖滴度提供了新的见解和思路,对GAstV感染引起的雏鹅痛风疾病的疫苗研发、生产和该疾病的防控具有重要意义。
Abstract:ObjectiveTo improve the proliferation efficiency of goose astrovirus (GAstV) by constructing a leghorn male hepatoma (LMH) cell line stably overexpressing vimentin (VIM) gene.
MethodThe VIM gene was amplified from goose-derived cells by PCR and subsequently ligated into the lentiviral gene overexpression system vector pLV-sfGFP (2A) Puro by homologous recombination to obtain the recombinant lentiviral plasmid pLV-sfGFP (2A) Puro-VIM-Flag(pLV-VIM). The 293T cells were used to package the recombinant lentivirus particles, which subsequently infected LMH cells and were screened with puromycin to obtain target cells. The effect of VIM on GAstV proliferation efficiency was evaluated by Western blot, RT-qPCR, indirect immunofluorescence and TCID50 assays. Finally, the effect of VIM on GAstV invasion was analyzed by antibody blocking test.
ResultThe recombinant lentiviral vector pLV-VIM was successfully constructed as confirmed by sequencing and restriction enzyme digestion. After screening, LMH-VIM cell line overexpressing the VIM gene (LMH-VIM) and its control group cell line (LMH-NC) were obtained. The results of Western blot showed that the expression level of VIM protein in LMH-VIM cells was significantly higher than that in LMH-NC cells. Overexpression of VIM significantly upregulated GAstV mRNA and protein expression levels in LMH-VIM cells, as well as viral titer in the cell supernatant. Antibody blocking test showed that cell surface VIM could actively promote GAstV invasion into cells.
ConclusionThe study provides a new insight and ideas for improving the proliferation titer of GAstV in the in vitro cell culture and is of great significance for the vaccine research and development, as well as prevention and control of gosling gout disease caused by GAstV infection.
-
新型鹅星状病毒(Goose astrovirus,GAstV)是近年来新发现的一种致雏鹅痛风且高度致死的禽星状病毒[1],感染后主要造成雏鹅内脏器官表面和关节腔内产生大量的尿酸盐沉积,感染率和死亡率分别可高达80%和50%左右[2-6]。GAstV属于星状病毒科禽星状病毒属3型禽星状病毒成员,为单股正链、无囊膜的RNA病毒[7]。
由GAstV造成的致死性雏鹅痛风疾病在我国广泛流行,给养鹅业造成了严重的经济损失[8-9];且该疾病疫苗研发的进展尤其缓慢,尚无疫苗可用,防控形势十分严峻。目前适用于GAstV增殖的可传代细胞系通常为鸡肝癌(Leghorn male hepatoma,LMH)细胞[10],但其培养的病毒滴度较鹅胚原代肾细胞低,在一定程度上限制了对该病毒致病机制的深入研究,同时也限制了该疾病疫苗研发的高效性和便利性。基于此,提高GAstV在LMH细胞中的增殖滴度对疫苗研发、生产和该疾病致病机制的研究具有重要意义。
波形蛋白(Vimentin,VIM)作为一种高度保守的中间纤维蛋白,最初发现于间叶细胞内,是细胞骨架的重要组成成分之一。VIM可影响细胞黏附与迁移,参与细胞的信号传导,调节细胞凋亡和增殖,并具有警报素功能[11-12]。VIM在细胞内高丰度表达,其在细胞表面及胞外的存在直到近些年才被发现,并逐渐有研究报道VIM在新型冠状病毒(SARS-CoV-2)[13-15]、猪繁殖与呼吸综合征病毒[16]、寨卡病毒[17]等多种病毒的侵入和复制过程中发挥重要作用。同时,病毒感染能诱导细胞内VIM重排,加强其与病毒蛋白的互作并促进病毒复制[18-20]。
目前关于VIM在GAstV中的作用的研究报道还较少。本研究在前期工作中利用鹅胚原代肾细胞及LMH细胞研究GAstV的感染机制,其中,以GAstV VP70为诱饵蛋白,通过GST pull-down和Co-IP等方法从鹅胚肾细胞的膜蛋白中捕获到众多关键蛋白分子,VIM便是其中之一;基于此,本研究推测VIM是潜在影响GAstV侵入和感染的关键调控因子之一。因此,研究VIM与GAstV病毒复制之间的关系,对于防控GAstV所致雏鹅痛风疾病具有重要意义。本研究拟通过慢病毒基因过表达系统构建可稳定过表达VIM基因的LMH细胞系,以期显著促进GAstV的增殖水平,为提高GAstV的增殖滴度提供新的见解和思路。
1. 材料与方法
1.1 材料
1.1.1 细胞、病毒与载体
LMH细胞、人肾上皮细胞系293T细胞、GAstV GD-ZJ-21-01病毒株由广东省农业科学院动物卫生研究所/农业农村部禽流感等家禽重大疾病防控重点实验室分离鉴定并保存;慢病毒基因过表达载体pLV-sfGFP (2A) Puro(货号:VL3408)及其辅助载体pH1和pH2均为北京英茂盛业生物科技有限公司产品。
1.1.2 主要试剂和抗体
高保真酶Phanta® Max Super-Fidelity DNA Polymerase(货号:P505)、无缝克隆试剂盒ClonExpress® II One Step Cloning Kit(货号:C112)为南京诺唯赞生物科技有限公司产品;内切酶Xba I(货号:R1045)和Xho I(货号:R1046)为New England Biolabs公司产品;总RNA提取试剂盒(货号:
220011 )为上海飞捷生物科技有限公司产品;DNA体外转染试剂PolyJet(货号:SL100688)为SignaGen公司产品;凝胶回收试剂盒DNA Gel Extraction Kit(货号:D2500)、无内毒素质粒提取试剂盒Endo-free Plasmid Midi Kit(货号:D6929)为Omega公司产品;反转录试剂盒PrimeScript RT Master Mix Real Time(货号:RR036A)为TaKaRa公司产品;荧光定量PCR酶Hieff® qPCR SYBR Green Master Mix(货号:11201ES08)为上海翌圣生物科技有限公司产品。鼠抗Flag-tag单克隆抗体(货号:F1804)为Sigma公司产品;兔抗GAPDH抗体(货号:ab181602)为Abcam公司产品;鼠抗Vimentin单克隆抗体(货号:sc-80975)为Santa公司产品;HRP标记的羊抗兔IgG抗体(货号:A0208)、HRP标记的羊抗鼠IgG抗体(货号:A0216)、Alexa Fluor 488标记羊抗兔IgG抗体(货号:A0423)均为上海碧云天生物技术有限公司产品;GAstV-2 VP27多克隆抗体由广东省农业科学院动物卫生研究所制备并惠赠[21]。1.2 方法
1.2.1 VIM基因的扩增及质粒构建
参照试剂盒说明书,从鹅源组织细胞中提取细胞总RNA并反转录为cDNA。参考NCBI中鹅源VIM基因序列(Genebank:XM_048061750.1),按照表1中自行设计的引物(VIM-F、VIM-R),以cDNA为模板通过PCR扩增VIM基因(1 380 bp)。以回收的VIM基因为模板,利用引物(VIM-F2、VIM-R-Flag)通过PCR进一步在VIM基因末端添加Flag标签与VIM基因融合表达(VIM-Flag)以便于检测;回收目的基因VIM-Flag,以其作为模板,利用同源臂引物(VIM-pLV-F、VIM-pLV-R)进行PCR扩增,使得VIM-Flag基因两端分别添加位于pLV-sfGFP (2A) Puro载体Xba I和Xho I酶切位点两侧至少长为20 bp左右的序列(VIM-Flag-pLV),以便其与Xba I和Xho I双酶切后的慢病毒载体进行同源重组。将上述带有同源臂的VIM-Flag-pLV基因与线性化的pLV-sfGFP (2A) Puro载体进行连接,并将连接产物转化至大肠埃希菌TOP10感受态细胞,即获得重组慢病毒表达质粒pLV-sfGFP (2A) Puro-VIM-Flag,下文简称pLV-VIM(图1)。
表 1 VIM基因扩增的PCR引物Table 1. PCR primers for VIM gene amplification引物名称
Primer name引物序列(5′→3′)1)
Primer sequence退火温度/℃
Annealing temperatureVIM-F ATGAGCATCAGCAGCAAGAA 54 VIM-R CTCCAAGTCATCATGGTGC VIM-F2 ATGAGCATCAGCAGCAAGAACTCCTCGTACC 64 VIM-R-Flag ttacttatcgtcgtcatccttgtaatcCTCCAAGTCATCATGG VIM-pLV-F ctcagatctcgaatttctagaATGAGCATCAGCAGCAAGAACTC 66 VIM-pLV-R gggcccgggttcgaactcgagTTACTTATCGTCGTCATCCTTGTAATC 1)小写字母区域为同源臂序列,下划线区域为Flag标签序列。
1) The lowercase letter area represents the homologous arm sequence, and the underlined area represents the Flag tag sequence.1.2.2 VIM基因重组慢病毒包装
在直径为10 cm的细胞培养皿中,待293T细胞单层融合至90%时进行转染。取无菌1.5 mL离心管(A管),加入500 μL DMEM培养基,随后加入20 μL的DNA体外转染试剂PolyJet并充分混匀;另取一无菌1.5 mL离心管(B管),加入500 μL DMEM培养基,随后加入待包装的慢病毒基因过表达质粒pLV-VIM或pLV-NC(对照)5 μg、慢病毒包装辅助载体pH1 3.75 μg及pH2 1.25 μg并充分混匀。将A管中已稀释好的转染试剂加入B管中,立刻充分混匀,于室温静置孵育10~15 min。在孵育等待时,用胰酶消化293T细胞,用含10%(φ)FBS的DMEM培养基制备成约6×105 mL−1的细胞悬液。根据需要制备的病毒量,将细胞悬液放入15 mL或50 mL离心管备用;将每1 mL转染混合液加入10 mL细胞悬液,轻轻吹吸细胞混匀;将混匀细胞悬液加入10 cm细胞培养皿,于37 ℃、5%(φ)CO2条件下培养24 h;去除含有转染试剂的培养基,清洗2遍细胞,加入10 mL慢病毒培养基[(DMEM培养基+10%(φ)FBS+1 mmol/L丙酮酸钠]。48 h后于4 ℃、500 r/min离心10 min去除细胞碎片,收集细胞培养液上清液;上清液中即为含有待表达基因的重组慢病毒颗粒,可直接用于靶细胞感染。
1.2.3 稳定过表达VIM基因的LMH细胞系构建及鉴定
取状态良好的LMH细胞均匀接种至24孔板中培养,待细胞长至70%~80%时弃培养液,加入pLV-VIM重组慢病毒悬液于37 ℃培养箱中孵育(同时设置空载体对照组);孵育2 h后弃病毒液,加入新的培养基继续培养传代;每次传代培养时加入终浓度为2 µg/mL的嘌呤霉素进行抗性筛选,得到的阳性多克隆株即为慢病毒介导的过表达VIM基因的LMH细胞系,命名为LMH-VIM,对照组为LMH-NC。筛选过程中,通过荧光显微镜观察自发绿色荧光的阳性细胞的占比,直至观察到几乎所有细胞均为阳性时,通过流式细胞术进行筛选验证。将筛选好的细胞系传代培养(含嘌呤霉素)10代后,提取细胞总蛋白,分别通过Flag抗体和VIM抗体进行Western blot检测,分析2种细胞系中VIM蛋白的表达水平。
1.2.4 流式细胞术
首先将未做任何处理的LMH细胞用胰酶消化后制备成细胞悬液(每毫升不超过108个细胞),随后上机至流式细胞仪,选取488 nm为激发波长,以此为阴性对照将试验参数调整到位。同理,将已制备好的自发绿色荧光的LMH-VIM和LMH-NC单细胞悬液上机至流式细胞仪,试验参数不变,对LMH-VIM和LMH-NC细胞中的绿色阳性细胞进行筛选分析,明确阳性细胞的比例;并将分选出的阳性细胞收集至离心管中,继续扩大培养,用于后续试验。
1.2.5 过表达VIM基因的LMH细胞对GAstV增殖效率的影响
以MOI=0.1的GAstV分别感染LMH-VIM和LMH-NC细胞,并置于37 ℃、5%(φ)CO2培养箱吸附2 h。吸附结束后弃病毒液,用PBS缓冲液清洗细胞3次,更换为含1%(φ)FBS的培养基进行培养。于感染24、48、72 h后收取细胞,提取细胞蛋白及RNA;通过Western blot及RT-qPCR分析GAstV在细胞内的复制水平,同时在72 h时收取细胞上清液进行TCID50测定。
1.2.6 RT-qPCR
参考总RNA提取试剂盒说明书提取细胞RNA,并将其反转录成cDNA。参考如下引物(GAstV ORF2:5′-GGGTGATCCGCAAGGAAATA-3′、5′-AAGTTTCGCCAGGGTTAGAG-3′;GAPDH:5′-GGAAAGTCATCCCTGAGCTG-3′、5′-GGTCAACAACAGAGACATTGG-3′)对目的基因进行RT-qPCR检测。反应体系:Hieff® qPCR SYBR Green Master Mix 10 μL、上下游引物(10 μmol/L)各1 μL、cDNA 1 μL、RNase Free ddH2O补足至20 μL。扩增条件:95 ℃预变性5 min;95 ℃变性15 s,60 ℃退火35 s,共40个循环。以GAPDH基因作为内参,按照2−△△Ct方法计算各个基因mRNA的相对表达水平。
1.2.7 Western blot
按照SDS-PAGE凝胶配制试剂盒说明书配制好凝胶;取出变性后的蛋白质,每孔加入等量蛋白质样品。在80 V恒压条件下垂直电泳直至溴酚蓝指示剂接近分离胶底部,随后在100 V恒压下将蛋白胶转印至硝酸纤维膜。取出硝酸纤维膜,用含5%(w)脱脂奶粉的TBST缓冲液封闭2 h;洗涤3次后加入一抗,于4 ℃孵育12~18 h;用TBST缓冲液洗涤3次,加入二抗,于室温孵育1~2 h,洗涤3次后通过凝胶成像系统观察并记录结果。
1.2.8 抗体封闭试验
为了明确细胞表面VIM对GAstV侵入细胞是否有影响,本研究进行了抗体封闭试验。将VIM单克隆抗体与LMH细胞于37 ℃孵育2 h(以IgG为对照),弃上清液,用PBS缓冲液清洗3次;随后加入GAstV病毒液(MOI=0.1)于37 ℃孵育2 h,弃上清液并清洗细胞3次,加入新鲜的DMEM完全培养基继续培养24 h;提取细胞蛋白及RNA,通过Western blot及RT-qPCR分析细胞内GAstV的复制水平;同时利用4%(w)多聚甲醛溶液固定细胞,进行GAstV的间接免疫荧光试验。
1.2.9 间接免疫荧光试验
将培养皿中的细胞用PBS缓冲液清洗3次;用4%(w)多聚甲醛溶液固定15 min、PBS缓冲液清洗3次后加入通透液孵育20 min;用PBS缓冲液清洗3次,加入封闭液于室温孵育30 min;弃封闭液,加入一抗于4 ℃孵育12~18 h;用PBST缓冲液清洗3次,弃废液,加入绿色荧光二抗,于37 ℃避光孵育1 h,PBST缓冲液清洗3次后在荧光显微镜下观察采集图像。
1.2.10 数据分析
相关试验至少进行3次独立重复,统计学分析使用GraphPad Prism软件,试验数据以平均值±标准差呈现,统计学差异按照t检验进行评估,P<0.05表示具有显著差异。
2. 结果与分析
2.1 VIM基因扩增
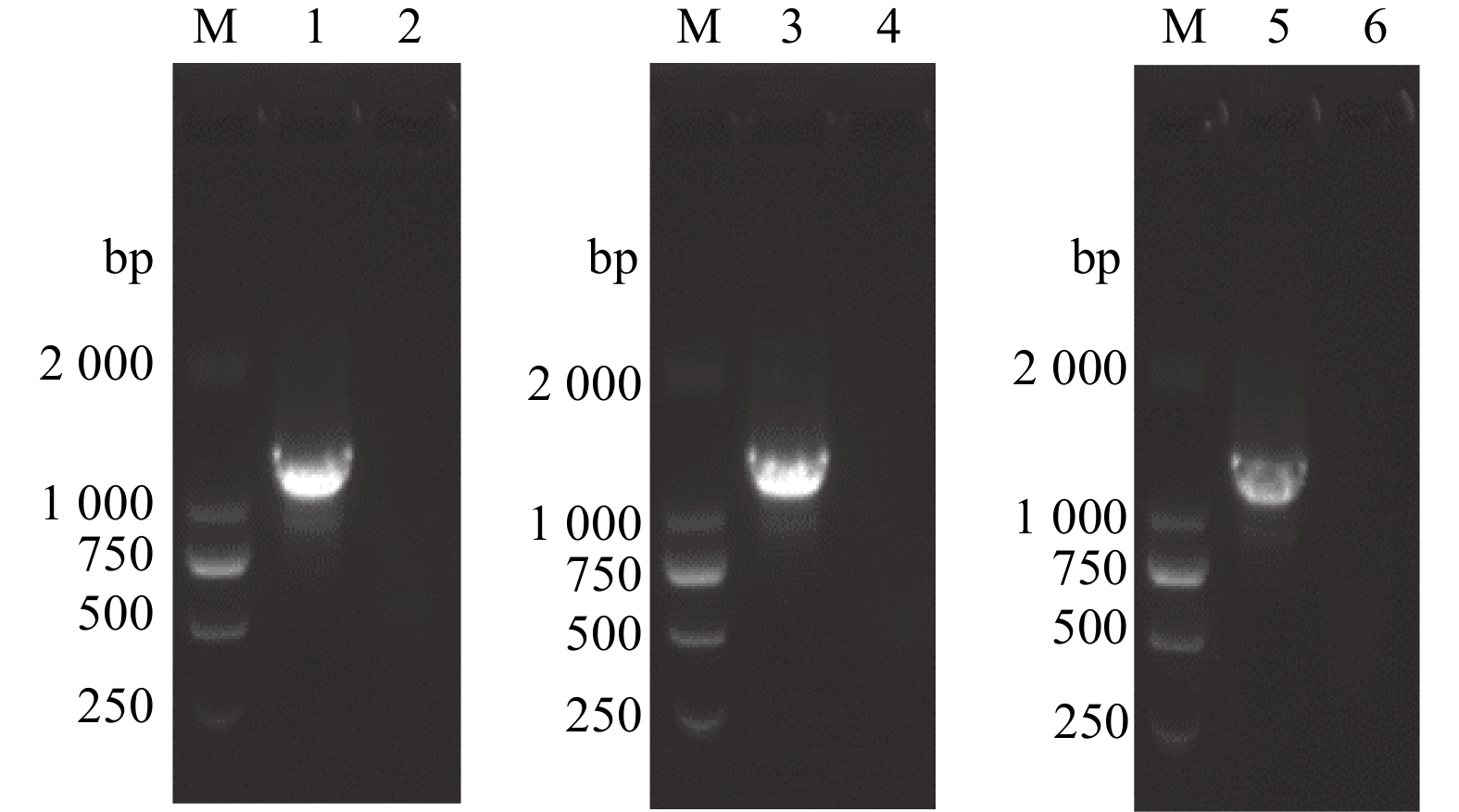
PCR扩增VIM基因后,通过10 g/L琼脂糖凝胶电泳观察到约1 383 bp的目的条带(VIM),与预期相符;回收目的基因后继续通过PCR在VIM基因末端添加Flag标签基因,经10 g/L琼脂糖凝胶电泳观察到约1 407 bp的目的条带(VIM-Flag);回收目的基因后继续通过PCR在VIM基因两端融合目标载体的同源臂,经10 g/L琼脂糖凝胶电泳观察到约1 449 bp的目的条带(VIM-Flag-pLV)(图2)。回收并纯化目的基因,测序结果显示其基因序列正确无误。
![]() 图 2 VIM基因的扩增M:DNA marker DL2000;1:以引物VIM-F、VIM-R进行扩增(VIM);3:以引物VIM-F2、VIM-R-Flag进行扩增(VIM-Flag);5:以引物VIM-pLV-F、VIM-pLV-R进行扩增(VIM-Flag-pLV);2、4、6:阴性对照。Figure 2. Amplification of VIM geneM: DNA marker DL2000; 1: Amplification with primers VIM-F and VIM-R (VIM); 3: Amplification with primers VIM-F2 and VIM-R-Flag (VIM-Flag); 5: Amplification with primers VIM-pLV-F and VIM-pLV-R (VIM-Flag-pLV); 2, 4, 6: Negative control.
图 2 VIM基因的扩增M:DNA marker DL2000;1:以引物VIM-F、VIM-R进行扩增(VIM);3:以引物VIM-F2、VIM-R-Flag进行扩增(VIM-Flag);5:以引物VIM-pLV-F、VIM-pLV-R进行扩增(VIM-Flag-pLV);2、4、6:阴性对照。Figure 2. Amplification of VIM geneM: DNA marker DL2000; 1: Amplification with primers VIM-F and VIM-R (VIM); 3: Amplification with primers VIM-F2 and VIM-R-Flag (VIM-Flag); 5: Amplification with primers VIM-pLV-F and VIM-pLV-R (VIM-Flag-pLV); 2, 4, 6: Negative control.2.2 重组慢病毒VIM基因过表达质粒的鉴定
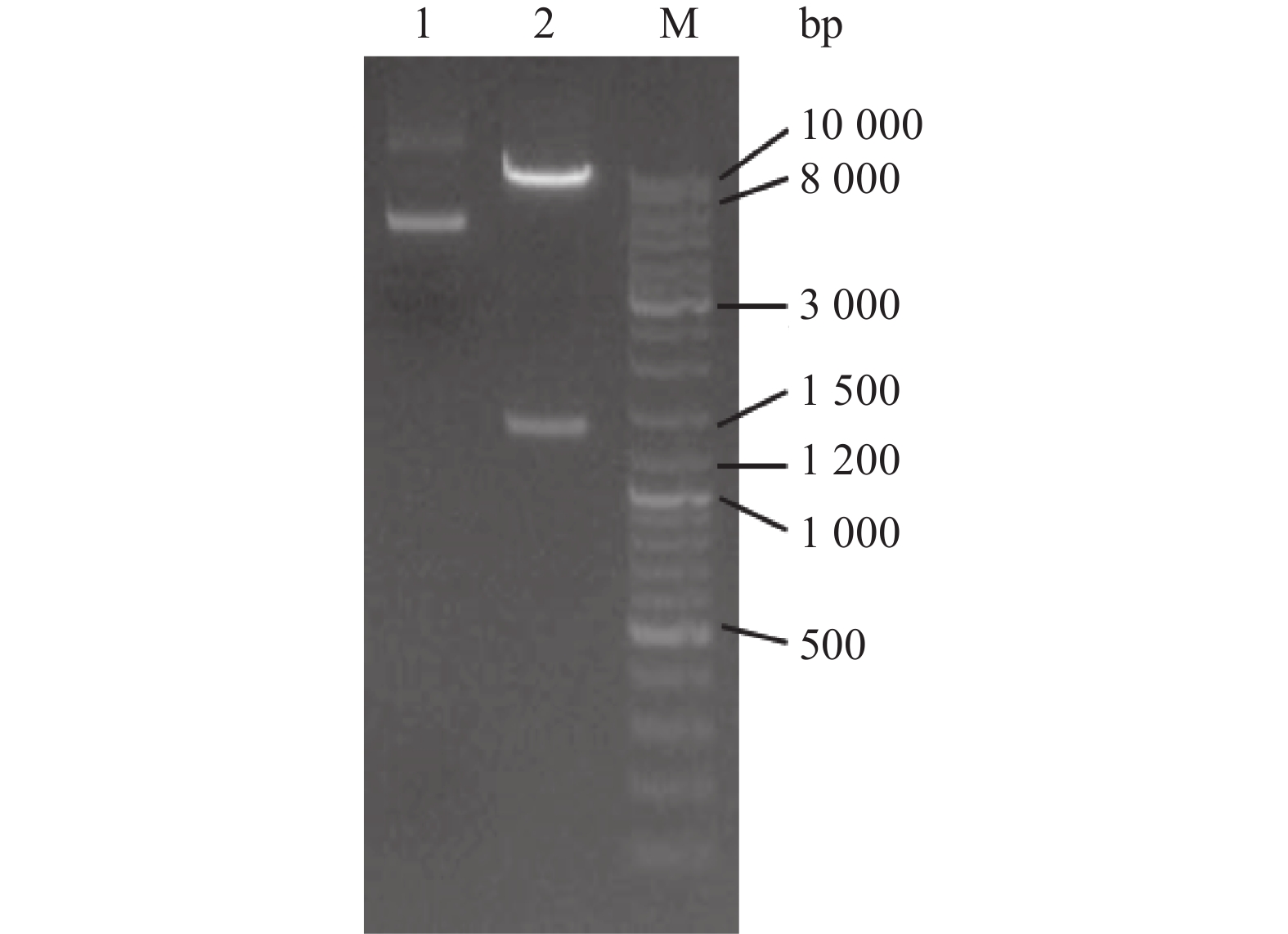
将构建好的pLV-VIM质粒进行双酶切(Xba I和Xho I),酶切后的产物进行琼脂糖凝胶电泳。结果(图3)显示,酶切产物经电泳后可见2条明显的条带,与预期1 395 bp(VIM基因)和9 000 bp(线性载体)保持一致,说明酶切验证无误。测序分析进一步表明该重组质粒成功构建。
![]() 图 3 pLV-VIM质粒双酶切后的琼脂糖凝胶电泳图1:未酶切的pLV-VIM质粒;2:经Xba I和Xho I双酶切的pLV-VIM质粒;M:DNA marker DL10000。Figure 3. Agarose gel electrophoresis graph of pLV-VIM plasmid after double digestion1: Undigested pLV-VIM plasmid; 2: pLV-VIM plasmid digested with Xba I and Xho I; M: DNA marker DL10000.
图 3 pLV-VIM质粒双酶切后的琼脂糖凝胶电泳图1:未酶切的pLV-VIM质粒;2:经Xba I和Xho I双酶切的pLV-VIM质粒;M:DNA marker DL10000。Figure 3. Agarose gel electrophoresis graph of pLV-VIM plasmid after double digestion1: Undigested pLV-VIM plasmid; 2: pLV-VIM plasmid digested with Xba I and Xho I; M: DNA marker DL10000.2.3 稳定过表达VIM基因的LMH细胞系的筛选及鉴定
将重组慢病毒颗粒感染LMH细胞后,在第1代细胞时仅有少量可自发绿色荧光的阳性细胞。由于嘌呤霉素可将阴性细胞杀死使得阳性细胞继续增殖,随着细胞的不断传代培养,阳性细胞逐渐增多,至第12代时,几乎所有细胞均为阳性细胞(图4),这一点也通过流式细胞术得到了验证(图5)。Western blot结果(图6A)显示,VIM-Flag融合蛋白成功在LMH-VIM细胞中表达,而对照组LMH-NC细胞中则未检测到该融合蛋白表达。另一方面,利用VIM单克隆抗体进行检测也能发现LMH-VIM细胞中VIM的表达水平显著高于对照组LMH-NC(图6B),表明稳定过表达VIM的LMH细胞系构建成功。
![]() 图 4 通过荧光显微镜观察绿色阳性细胞Figure 4. Observation of green positive cells through fluorescence microscope
图 4 通过荧光显微镜观察绿色阳性细胞Figure 4. Observation of green positive cells through fluorescence microscope![]() 图 5 通过流式细胞术筛选绿色阳性细胞Figure 5. Screening of green positive cells through flow cytometry
图 5 通过流式细胞术筛选绿色阳性细胞Figure 5. Screening of green positive cells through flow cytometry![]() 图 6 通过Western blot检测VIM蛋白的表达水平Mr 表示相对分子质量。Figure 6. Detection of VIM protein expression level through Western blotMr indicates relative molecular mass.
图 6 通过Western blot检测VIM蛋白的表达水平Mr 表示相对分子质量。Figure 6. Detection of VIM protein expression level through Western blotMr indicates relative molecular mass.2.4 过表达VIM基因的LMH细胞系对GAstV增殖效率的影响
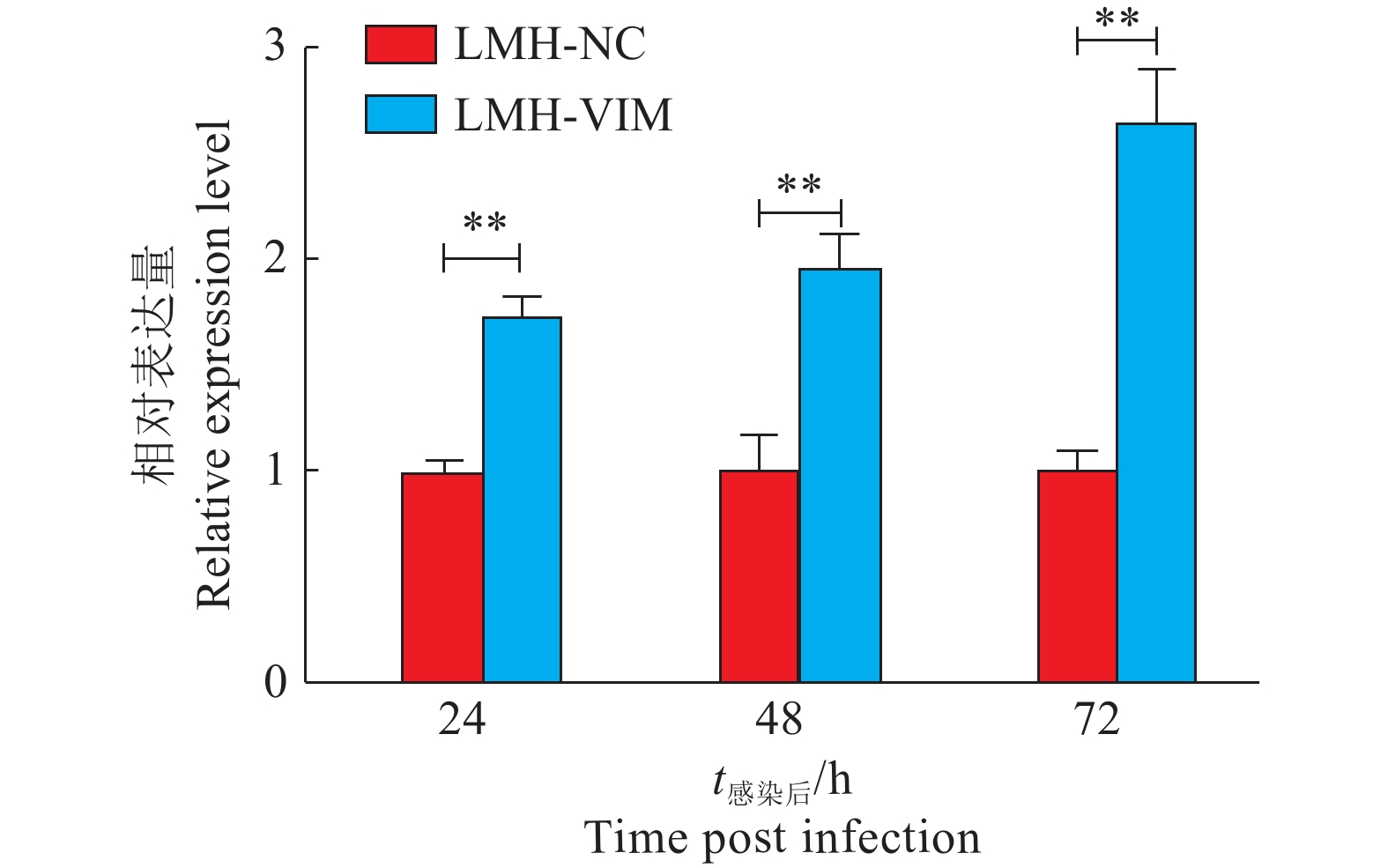
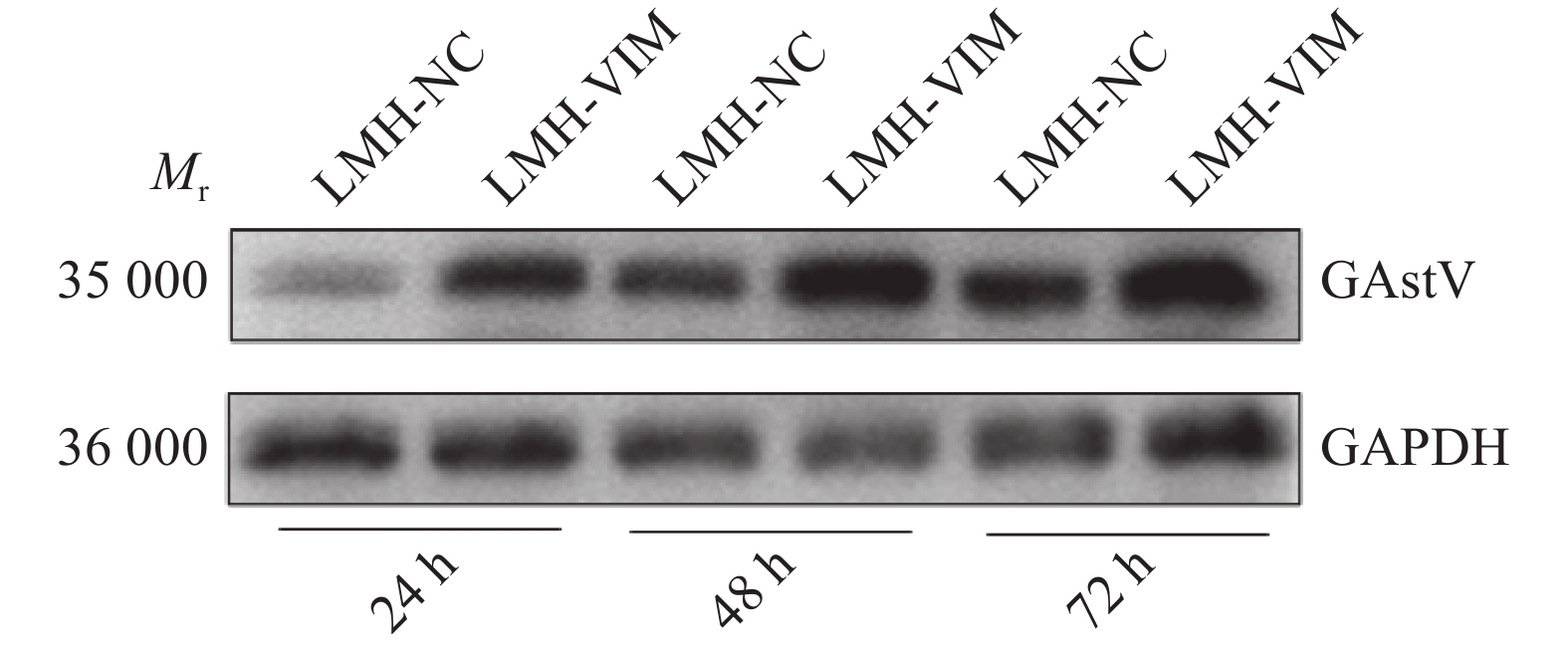
在过表达VIM基因的LMH细胞系(LMH-VIM)中,不论是病毒的mRNA表达水平抑或蛋白表达水平,均显著高于LMH-NC细胞(P<0.01,图7、8)。除此之外,收集感染72 h后的细胞上清液进行病毒TCID50的测定,LMH-VIM细胞上清液中的病毒滴度显著高于对照组LMH-NC(P<0.001,图9)。以上结果表明,过表达VIM的LMH细胞系对GAstV的增殖具有显著的促进作用。
![]() 图 7 感染VIM基因对GAstV mRNA表达水平的影响“**”表示在P<0.01水平差异显著(t检验)。Figure 7. Effect of VIM gene infection on GAstV mRNA expression level“**” indicates significant differences at P<0.01 (t test).
图 7 感染VIM基因对GAstV mRNA表达水平的影响“**”表示在P<0.01水平差异显著(t检验)。Figure 7. Effect of VIM gene infection on GAstV mRNA expression level“**” indicates significant differences at P<0.01 (t test).![]() 图 8 不同VIM基因感染时间对GAstV蛋白表达水平的影响Mr表示相对分子质量。Figure 8. Effect of different VIM gene infection time on GAstV protein expression levelMr indicates relative molecular mass.
图 8 不同VIM基因感染时间对GAstV蛋白表达水平的影响Mr表示相对分子质量。Figure 8. Effect of different VIM gene infection time on GAstV protein expression levelMr indicates relative molecular mass.![]() 图 9 病毒TCID50测定“***”表示在P<0.001水平差异显著(t检验)。Figure 9. Virus TCID50 determination“***” indicates significant difference at P<0.001 (t test)
图 9 病毒TCID50测定“***”表示在P<0.001水平差异显著(t检验)。Figure 9. Virus TCID50 determination“***” indicates significant difference at P<0.001 (t test)2.5 细胞表面VIM促进GAstV侵入细胞
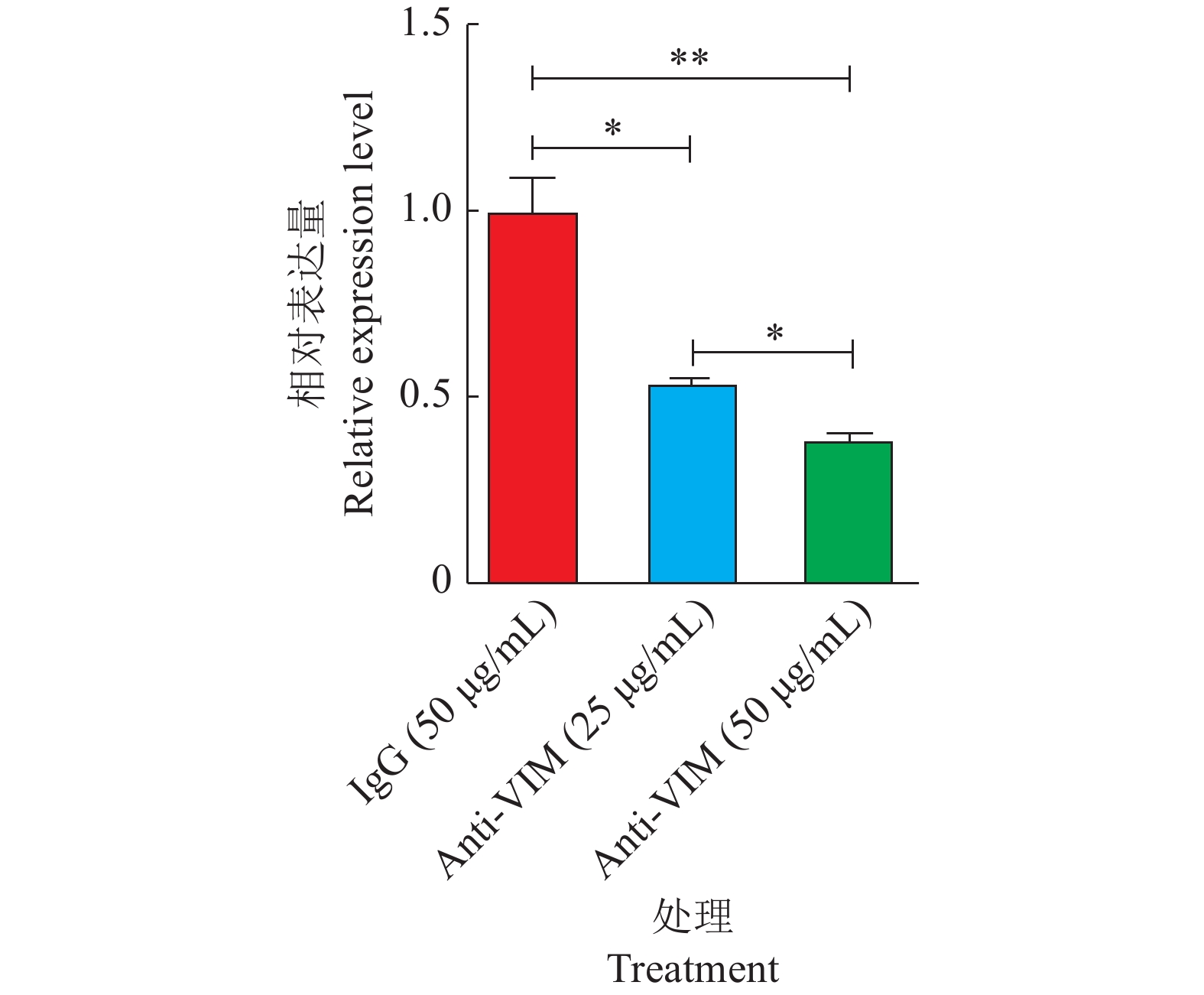
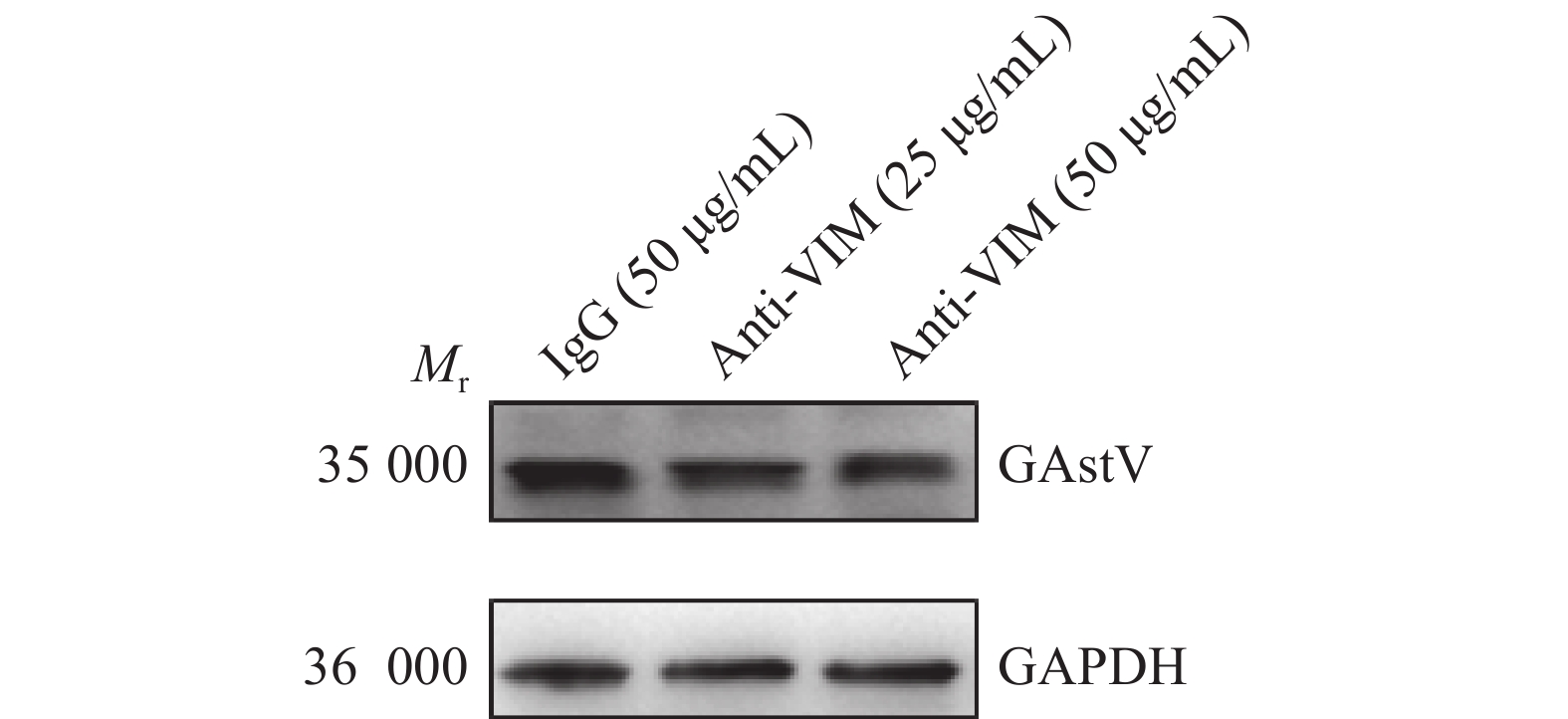
当利用VIM单克隆抗体阻断细胞表面VIM时,GAstV侵入细胞内的效率降低(图10),其在LMH细胞中的mRNA和蛋白表达水平均显著下降(P<0.05,图11、12)。由以上结果可知,VIM单克隆抗体抑制GAstV侵入细胞具有浓度依赖性;当抗体浓度越高,阻止病毒侵入细胞的作用越显著(P<0.05)。以上结果提示VIM可能是GAstV的辅助受体之一,进而影响GAstV的复制。
![]() 图 10 GAstV VP27间接免疫荧光试验结果Figure 10. Indirect immunofluorescence experimental result of GAstV VP27
图 10 GAstV VP27间接免疫荧光试验结果Figure 10. Indirect immunofluorescence experimental result of GAstV VP27![]() 图 11 阻断细胞表面VIM对GAstV mRNA表达水平的影响“*”“**”分别表示在P<0.05和P<0.01水平差异显著(t检验)。Figure 11. Effect of blocking cell surface VIM on GAstV mRNA expression level“*” and “**” indicate significant differences at P<0.05 and P<0.01, respectively (t test).
图 11 阻断细胞表面VIM对GAstV mRNA表达水平的影响“*”“**”分别表示在P<0.05和P<0.01水平差异显著(t检验)。Figure 11. Effect of blocking cell surface VIM on GAstV mRNA expression level“*” and “**” indicate significant differences at P<0.05 and P<0.01, respectively (t test).![]() 图 12 阻断细胞表面VIM对GAstV蛋白表达水平的影响Mr表示相对分子质量。Figure 12. Effect of blocking cell surface VIM on GAstV protein expression levelMr indicates relative molecular mass.
图 12 阻断细胞表面VIM对GAstV蛋白表达水平的影响Mr表示相对分子质量。Figure 12. Effect of blocking cell surface VIM on GAstV protein expression levelMr indicates relative molecular mass.3. 讨论与结论
3.1 稳定过表达VIM基因的LMH细胞株的构建
慢病毒基因过表达系统是目前应用最广泛的构建稳定表达外源蛋白细胞系的方法,该病毒包装系统的基本原理是将携带目的基因的慢病毒载体整合进宿主细胞基因组,以实现目的基因在宿主细胞中的持续表达[22],可用于研究某种蛋白的功能及其对病毒复制的影响;如稳定表达人HDAC6基因的Vero细胞系的构建及其对狂犬病病毒增殖的影响[23],非洲猪瘟病毒的D1133L蛋白在MA-104细胞中的稳定过表达[24],稳定过表达人TMPRSS2基因的BHK细胞系的构建及其对新城疫病毒增殖效果的影响[25]等。
外源目的基因导入细胞有多种方式,但大部分方法不能将外源基因整合入基因组,且导入的目的基因容易在筛选后的传代培养过程中丢失和突变;而慢病毒载体可以将携带的外源基因导入宿主细胞中,通过反转录将目的基因整合到细胞基因组,并稳定表达,具有操作简便、扩增周期短、转染效率高、能长期稳定表达的特点,且能够感染分裂和不分裂细胞,适用于难转染的原代细胞,在研究中被广泛使用[26]。因此,本研究基于慢病毒基因过表达系统,利用VIM基因过表达的重组慢病毒颗粒感染LMH细胞,通过嘌呤霉素进行初步筛选;筛选至第12代时,在显微镜下观察到几乎所有细胞均自发荧光,表明VIM在绝大多数细胞中成功过表达。为明确是否所有细胞均过表达VIM基因,本研究通过流式细胞术对其进行分选,将分选得到的阳性细胞继续扩大培养,并进行VIM蛋白表达分析;结果显示VIM蛋白成功过表达,即得到过表达VIM基因的LMH细胞系。这为寻找更优的GAstV增殖系统提供了研究思路,也为GAstV致病机制的深入研究及抗病毒策略的探索提供了理论基础。除此之外,在后续的研究中,可利用该系统将细胞永生化相关基因整合至鹅源原代细胞(如肾细胞、肝细胞等)中,以尝试构建鹅源可传代细胞系,将为GAstV的高效增殖提供更佳的解决方案。
3.2 VIM对GAstV复制的影响
VIM是中间丝蛋白家族中广泛表达和高度保守的蛋白之一,主要表达于中胚层来源的细胞中,广泛存在于成纤维细胞、内皮细胞和免疫系统细胞中;其由4个α−螺旋片段组成的杆状蛋白组成,头部为非螺旋体的氨基端,尾部为羧基端,在荧光显微镜下显示为网状海绵结构,与微管、微丝一起构成细胞网格[27]。作为骨架蛋白,VIM自身很少引起研究者的关注,但除了发挥骨架蛋白应有的物理功能外,VIM还参与多种病原体的入侵过程,并在多种病毒的感染过程中发挥重要作用。Stefanovic等[20]研究发现,非洲猪瘟病毒感染导致细胞中VIM重排,在病毒装配位点的周围形成“笼状”结构,并且这一结构的形成与病毒晚期结构蛋白的出现同步,说明病毒晚期蛋白的表达可能需要VIM的参与。Zheng等[28]对传染性法氏囊病病毒感染鸡胚成纤维细胞的蛋白质组学进行研究,发现该病毒显著下调VIM的表达水平,并且VIM的网状结构被破坏,推测其可能与病毒粒子的释放有关。Zhang等[29]研究表明,蓝舌病毒衣壳蛋白VP2与VIM的相互作用可促进病毒的释放。Zhang等[30]研究表明,VIM与猪传染性胃肠炎病毒的N蛋白存在直接互作,沉默VIM抑制TGEV的释放。对猪繁殖与呼吸障碍综合症病毒的研究发现,VIM是该病毒受体复合物的组成成分,VIM抗体孵育后的Marc-145细胞能阻断病毒侵入[31]。
本研究发现,稳定过表达VIM基因的LMH细胞可显著促进GAstV的mRNA和蛋白表达水平,并提高细胞上清液中GAstV的病毒滴度。在此过程中,细胞表面VIM蛋白在病毒侵入细胞的过程中发挥重要角色,即当利用VIM单克隆抗体阻断细胞表面VIM蛋白时,显著抑制GAstV侵入细胞的效率,但并无法完全阻断病毒侵入细胞;这表明VIM并不是GAstV的主要功能性受体,可能是其辅助受体之一,但还需要进一步的佐证,譬如进行VIM基因的敲除试验,或在非易感细胞中进行受体重建试验。另一方面,Xiang等[32]研究发现,VIM是GAstV结构蛋白VP70的宿主结合伴侣之一,VIM可通过与VP70相互作用以正向调节GAstV的复制,完整的VIM网格结构及VIM-VP70互作是GAstV高效复制所必需的;这便揭示了VIM促进GAstV复制的分子机理。同时,本研究也正不断探索VIM调控GAstV复制的深层次原因,以期为GAstV的科学防控提供新的理论基础。
3.3 结论
本研究利用慢病毒基因过表达系统,成功构建了稳定过表达VIM基因的LMH细胞系(LMH-VIM),其可显著促进GAstV复制,提高病毒增殖滴度。本研究初步探究了VIM与GAstV增殖的相关性,为深入阐明VIM调控GAstV复制的分子机制提供了理论基础,也为GAstV的防控和抗病毒研究提供了潜在的候选靶点。
-
![]()
图 2 VIM基因的扩增
M:DNA marker DL2000;1:以引物VIM-F、VIM-R进行扩增(VIM);3:以引物VIM-F2、VIM-R-Flag进行扩增(VIM-Flag);5:以引物VIM-pLV-F、VIM-pLV-R进行扩增(VIM-Flag-pLV);2、4、6:阴性对照。
Figure 2. Amplification of VIM gene
M: DNA marker DL2000; 1: Amplification with primers VIM-F and VIM-R (VIM); 3: Amplification with primers VIM-F2 and VIM-R-Flag (VIM-Flag); 5: Amplification with primers VIM-pLV-F and VIM-pLV-R (VIM-Flag-pLV); 2, 4, 6: Negative control.
![]()
图 3 pLV-VIM质粒双酶切后的琼脂糖凝胶电泳图
1:未酶切的pLV-VIM质粒;2:经Xba I和Xho I双酶切的pLV-VIM质粒;M:DNA marker DL10000。
Figure 3. Agarose gel electrophoresis graph of pLV-VIM plasmid after double digestion
1: Undigested pLV-VIM plasmid; 2: pLV-VIM plasmid digested with Xba I and Xho I; M: DNA marker DL10000.
![]()
图 4 通过荧光显微镜观察绿色阳性细胞
Figure 4. Observation of green positive cells through fluorescence microscope
![]()
图 5 通过流式细胞术筛选绿色阳性细胞
Figure 5. Screening of green positive cells through flow cytometry
![]()
图 6 通过Western blot检测VIM蛋白的表达水平
Mr 表示相对分子质量。
Figure 6. Detection of VIM protein expression level through Western blot
Mr indicates relative molecular mass.
![]()
图 7 感染VIM基因对GAstV mRNA表达水平的影响
“**”表示在P<0.01水平差异显著(t检验)。
Figure 7. Effect of VIM gene infection on GAstV mRNA expression level
“**” indicates significant differences at P<0.01 (t test).
![]()
图 8 不同VIM基因感染时间对GAstV蛋白表达水平的影响
Mr表示相对分子质量。
Figure 8. Effect of different VIM gene infection time on GAstV protein expression level
Mr indicates relative molecular mass.
![]()
图 9 病毒TCID50测定
“***”表示在P<0.001水平差异显著(t检验)。
Figure 9. Virus TCID50 determination
“***” indicates significant difference at P<0.001 (t test)
![]()
图 10 GAstV VP27间接免疫荧光试验结果
Figure 10. Indirect immunofluorescence experimental result of GAstV VP27
![]()
图 11 阻断细胞表面VIM对GAstV mRNA表达水平的影响
“*”“**”分别表示在P<0.05和P<0.01水平差异显著(t检验)。
Figure 11. Effect of blocking cell surface VIM on GAstV mRNA expression level
“*” and “**” indicate significant differences at P<0.05 and P<0.01, respectively (t test).
![]()
图 12 阻断细胞表面VIM对GAstV蛋白表达水平的影响
Mr表示相对分子质量。
Figure 12. Effect of blocking cell surface VIM on GAstV protein expression level
Mr indicates relative molecular mass.
表 1 VIM基因扩增的PCR引物
Table 1 PCR primers for VIM gene amplification
引物名称
Primer name引物序列(5′→3′)1)
Primer sequence退火温度/℃
Annealing temperatureVIM-F ATGAGCATCAGCAGCAAGAA 54 VIM-R CTCCAAGTCATCATGGTGC VIM-F2 ATGAGCATCAGCAGCAAGAACTCCTCGTACC 64 VIM-R-Flag ttacttatcgtcgtcatccttgtaatcCTCCAAGTCATCATGG VIM-pLV-F ctcagatctcgaatttctagaATGAGCATCAGCAGCAAGAACTC 66 VIM-pLV-R gggcccgggttcgaactcgagTTACTTATCGTCGTCATCCTTGTAATC 1)小写字母区域为同源臂序列,下划线区域为Flag标签序列。
1) The lowercase letter area represents the homologous arm sequence, and the underlined area represents the Flag tag sequence. 下载: 导出CSV
下载: 导出CSV
-
[1] ZHANG X, REN D, LI T, et al. An emerging novel goose astrovirus associated with gosling gout disease, China[J]. Emerging Microbes & Infections, 2018, 7(1). doi: 10.1038/s41426-018-0153-7.
[2] XU J, GAO L, ZHU P, et al. Isolation, identification, and pathogenicity analysis of newly emerging gosling astrovirus in South China[J]. Frontiers in Microbiology, 2023, 14: 1112245. doi: 10.3389/fmicb.2023.1112245.
[3] LIU C, LI L, DONG J, et al. Global analysis of gene expression profiles and gout symptoms in goslings infected with goose astrovirus[J]. Veterinary Microbiology, 2023, 279: 109677. doi: 10.1016/j.vetmic.2023.109677.
[4] ZHANG X, DENG T, SONG Y, et al. Identification and genomic characterization of emerging goose astrovirus in central China, 2020[J]. Transboundary and Emerging Diseases, 2022, 69(3): 1046-1055. doi: 10.1111/tbed.14060
[5] 王昊锋. 新型鹅星状病毒引起组织损伤特性研究及感染靶细胞的鉴定[D]. 泰安: 山东农业大学, 2022. [6] 黎洁玉. 新型鹅星状病毒的分离鉴定与致病性分析[D]. 长沙: 湖南大学, 2021. [7] CHEN Q, XU X, YU Z, et al. Characterization and genomic analysis of emerging astroviruses causing fatal gout in goslings[J]. Transboundary and Emerging Diseases, 2020, 67(2): 865-876. doi: 10.1111/tbed.13410
[8] ZHANG F, LI H, WEI Q, et al. Isolation and phylogenetic analysis of goose astrovirus type 1 from goslings with gout in Jiangxi province, China[J]. Poultry Science, 2022, 101(7): 101800. doi: 10.1016/j.psj.2022.101800.
[9] PENG Z, GAO D, SONG X, et al. Isolation and genomic characterization of one novel goose astrovirus causing acute gosling gout in China[J]. Scientific Reports, 2023, 13(1): 10565. doi: 10.1038/s41598-023-37784-9.
[10] REN D, LI T, ZHANG X, et al. OASL triggered by novel goose astrovirus via ORF2 restricts its replication[J]. Journal of Virology, 2020, 94(24): e01767-20.
[11] SALDANHA R, THANH M T H, KRISHNAN N, et al. Vimentin supports cell polarization by enhancing centrosome function and microtubule acetylation[J]. Journal of the Royal Society Interface, 2024, 21(215): 20230641. doi: 10.1098/rsif.2023.0641.
[12] MONTEIRO-REIS S, MIRANDA-GONÇALVES V, GUIMARÃES-TEIXEIRA C, et al. Vimentin epigenetic deregulation in Bladder Cancer associates with acquisition of invasive and metastatic phenotype through epithelial-to-mesenchymal transition[J]. International Journal of Biological Sciences, 2023, 19(1): 1-12. doi: 10.7150/ijbs.77181
[13] SUPREWICZ Ł, SWOGER M, GUPTA S, et al. Extracellular vimentin as a target against SARS-CoV-2 host cell invasion[EB/OL]. bioRxiv: 2021.01. 08.425793 (2021-03-18) [2024-07-10]. doi: 10.1002/smll.202105640.
[14] LALIOTI V, GONZÁLEZ-SANZ S, LOIS-BERMEJO I, et al. Cell surface detection of vimentin, ACE2 and SARS-CoV-2 Spike proteins reveals selective colocalization at primary cilia[J]. Scientific Reports, 2022, 12(1): 7063. doi: 10.1038/s41598-022-11248-y.
[15] AMRAEI R, XIA C, OLEJNIK J, et al. Extracellular vimentin is an attachment factor that facilitates SARS-CoV-2 entry into human endothelial cells[J]. Proceedings of the National Academy of Sciences of the United States of America, 2022, 119(6): e2113874119.
[16] ZHENG X, LI R, QIAO S, et al. Vimentin rearrangement by phosphorylation is beneficial for porcine reproductive and respiratory syndrome virus replication in vitro[J]. Veterinary Microbiology, 2021, 259: 109133. doi: 10.1016/j.vetmic.2021.109133.
[17] ZHANG Y, ZHAO S, LI Y, et al. Host cytoskeletal vimentin serves as a structural organizer and an RNA-binding protein regulator to facilitate Zika viral replication[J]. Proceedings of the National Academy of Sciences of the United States of America, 2022, 119(8): e2113909119.
[18] ZHU Z, LI W, ZHANG X, et al. Foot-and-mouth disease virus capsid protein VP1 interacts with host ribosomal protein SA to maintain activation of the MAPK signal pathway and promote virus replication[J]. Journal of Virology, 2020, 94(3): e01350-19.
[19] GLADUE D P, O’DONNELL V, BAKER-BRANSTETTER R, et al. Foot-and-mouth disease virus modulates cellular vimentin for virus survival[J]. Journal of Virology, 2013, 87(12): 6794-6803. doi: 10.1128/JVI.00448-13
[20] STEFANOVIC S, WINDSOR M, NAGATA K, et al. Vimentin rearrangement during African swine fever virus infection involves retrograde transport along microtubules and phosphorylation of vimentin by calcium calmodulin kinase II[J]. Journal of Virology, 2005, 79(18): 11766-11775. doi: 10.1128/JVI.79.18.11766-11775.2005
[21] 向勇, 李林林, 张俊勤, 等. 2型鹅星状病毒VP27蛋白的原核表达及多克隆抗体制备[J]. 中国家禽, 2024, 46(9): 187-193. [22] PARK F, KAY M A. Modified HIV-1 based lentiviral vectors have an effect on viral transduction efficiency and gene expression in vitro and in vivo[J]. Molecular Therapy, 2001, 4(3): 164-173. doi: 10.1006/mthe.2001.0450
[23] 殷娟斌, 张志雄, 王莎莎, 等. 过表达HDAC6基因的Vero细胞系建立及其对狂犬病病毒增殖效率评价[J]. 中国兽医科学, 2023, 53(2): 150-155. [24] 陈玲玲, 张婷, 郝雨, 等. 非洲猪瘟病毒D1133L蛋白增加宿主波形蛋白磷酸化而促进病毒在猪巨噬细胞中的复制[J]. 微生物学报, 2024, 64(3): 720-732. [25] 刘伟, 李梦娇, 郭佩东, 等. 稳定表达人TMPRSS2基因BHK细胞系的建立及其对新城疫弱毒增殖效率评价[J]. 中国兽医科学, 2021, 51(5): 594-600. [26] KLIMPEL M, TERRAO M, CHING N, et al. Development of a perfusion process for continuous lentivirus production using stable suspension producer cell lines[J]. Biotechnology and Bioengineering, 2023, 120(9): 2622-2638. doi: 10.1002/bit.28413
[27] RAMOS I, STAMATAKIS K, OESTE C L, et al. Vimentin as a multifaceted player and potential therapeutic target in viral infections[J]. International Journal of Molecular Sciences, 2020, 21(13): 4675. doi: 10.3390/ijms21134675.
[28] ZHENG X, HONG L, SHI L, et al. Proteomics analysis of host cells infected with infectious bursal disease virus[J]. Molecular & Cellular Proteomics, 2008, 7(3): 612-625.
[29] ZHANG X, BOYCE M, BHATTACHARYA B, et al. Bluetongue virus coat protein VP2 contains sialic acid-binding domains, and VP5 resembles enveloped virus fusion proteins[J]. Proceedings of the National Academy of Sciences of the United States of America, 2010, 107(14): 6292-6297.
[30] ZHANG X, SHI H, CHEN J, et al. Identification of the interaction between vimentin and nucleocapsid protein of transmissible gastroenteritis virus[J]. Virus Research, 2015, 200: 56-63. doi: 10.1016/j.virusres.2014.12.013
[31] KIM J K, FAHAD A M, SHANMUKHAPPA K, et al. Defining the cellular target(s) of porcine reproductive and respiratory syndrome virus blocking monoclonal antibody 7G10[J]. Journal of Virology, 2006, 80(2): 689-696. doi: 10.1128/JVI.80.2.689-696.2006
[32] XIANG Y, LI L, HUANG Y, et al. Cellular vimentin interacts with VP70 protein of goose astrovirus genotype 2 and acts as a structural organizer to facilitate viral replication[J]. Poultry Science, 2024, 103(10): 104146. doi: 10.1016/j.psj.2024.104146.
-
期刊类型引用(5)
1. 张乐,于小航,王义鹤,杨佳美,赵丽丽,段铭. 2019—2020年河南与河北部分地区犬瘟热流行病学调查及病毒分离鉴定. 广东农业科学. 2023(02): 95-103 .  百度学术
百度学术
2. 罗世民,李中波. 怀化地区犬瘟热病毒H、F基因遗传多样性及其遗传进化分析. 中国畜牧兽医. 2023(08): 3248-3257 . 百度学术
3. 张建普,马丽利,周利锋. 犬瘟热病毒YD株的分离鉴定及其H基因序列分析. 畜牧与兽医. 2021(04): 69-75 . 百度学术
4. 蒋梅,陈武,翟俊琼,卜婉迪,谢逸伦,刘灿彬,单芬,罗满林. 小熊猫源犬瘟热病毒株H、F基因的克隆及序列分析. 畜牧兽医学报. 2021(06): 1670-1676 . 百度学术
5. 刘健,白艺兰,鞠厚斌,潘子豪,俞向前,杨显超,朱晓英,葛菲菲,葛杰,于讳茹,陶田谷晟,王建,赵洪进. 犬瘟热病毒SH202003株全基因组序列分析. 中国畜牧兽医. 2021(07): 2559-2568 . 百度学术
其他类型引用(1)

计量
- 文章访问数: 50
- HTML全文浏览量: 1
- PDF下载量: 19
- 被引次数: 6
