
Extraction of pig contour based on fully convolutional networks
-
摘要:目的
实现猪舍场景下非接触、低成本的生猪轮廓高效提取。
方法以真实养殖环境下的生猪个体为研究对象,提出一种基于VGG16 与UNET相结合的全卷积神经网络模型(VGG-UNET模型)。该模型采用批处理方法,迁移学习VGG16模型参数,通过在模型中构建复制通道深度融合图像深层抽象特征与浅层特征,实现对图像语义级别分割。在30头长白生猪的1 815张数据集上进行模型验证,通过设置不同批大小对比试验,并选取其中具有最佳效果的3组探讨批大小与评价指标值变化趋势间的关系。
结果测试集上的对比试验结果表明,VGG-UNET模型在像素精度与均交并比方面分别达到94.32% 和86.60%,比单独采用UNET模型分别高出0.89%和1.67%。不同指标值变化情况与批大小间的关系不尽相同。在本文试验环境下,批大小对模型收敛速度的影响不明显。不同批大小条件下PA及MIoU指标值变化综合分析得出,VGG-UNET模型具有较强稳定性和较高鲁棒性;批大小为8 的情况下VGG-UNET模型效果最佳。
结论本文提出的生猪轮廓提取方法(VGG-UNET模型)是有效的,能实现精确、稳定的生猪轮廓提取,且分割结果较为完整,同时模型具有较高鲁棒性,可为后续生猪个体识别研究提供参考。
Abstract:ObjectiveTo realize non-contacting and low-cost pig contour extraction under the piggery scene.
MethodWe took individual pig in the real culture environment as the research object, and proposed a full convolutional neural network model based on the combination of VGG16 and UNET (VGG-UNET model). We adopted the batch processing method in this model to transfer and learn the parameters of VGG16 model. We achieved semantic level segmentation of the image by combining the deep abstract feature and shallow feature in depth via building the duplicate channel. The model was verified on 1 815 datasets of 30 Large White× Landrace pigs. Comparison experiments of different batch sizes were performed, and three groups with the best results were selected to explore the relationship between batch size and the evaluation index.
ResultThrough comparison experiments on datasets, the pixel accuracy and mean intersection-over-union of VGG-UNET model were 94.32% and 86.60% respectively, which were 0.89% and 1.67% higher than those of the UNET model. The experiments showed different relationship between the change of different index values and batch size. Batch size had no obvious impact on the convergence rate of the model under this experimental environment. Through comprehensive analysis of PA and MIoU index values under different batch sizes, the VGG-UNET model showed the highest stability and robustness, and it was found to be the best when the batch size was 8.
ConclusionThe VGG-UNET model is effective for accurate and stable extraction of pig contour. Such segmentation result is relatively complete and the model has higher robustness, which can provide a reference for follow-up identification of individual pigs.
-
水稻Oryza sativa L.是重要的粮食作物之一,预计到2030年水稻产量需增加40%才能适应人口增长的需求[1]。2050年世界人口将增至90多亿,如何养活日益增加的人口是全世界面临的共同问题[2]。培育高产品种是世界粮食安全的重要保障之一。我国约60%人口以稻米为主食,然而,水稻生产面临着人口不断增加、耕地面积逐渐减少、生长环境恶劣等严峻问题,要实现“中国人的饭碗任何时候都要牢牢端在自己手中,饭碗主要装中国粮”这一目标,不断提高水稻产量、品质和抗性是保障我国粮食安全的重大战略需求。
20世纪60年代以来,矮化育种、杂种优势利用促进水稻产量有了质的飞跃,极大满足了我国人口增长和生活水平提高对粮食增长的需求,这一巨大成就得益于半矮秆基因、野败型细胞质雄性不育基因的发现与应用。半矮秆基因资源有利于水稻品种(或组合)高产,却导致过度施用化肥、农药和灌溉水,引发了水稻生产与生态环境的矛盾。为了缓解资源趋紧、环境污染严重、生态系统退化的严峻问题,“绿色超级稻”新理念主张以功能基因组研究的成果为基础,大力培育环境友好型的“绿色超级稻”新品种(组合),因此亟需更多有利的稻种基因资源[3]。然而,近几十年来,水稻育种目标主要以提高商业经济效益为主,多数是在推广的优良品种上进行改良,致使栽培稻遗传基础变窄,种内遗传多样性降低,对未来水稻安全生产构成一定的潜在风险。野生稻或地方品种在自然或栽培条件下经长期自然或人工选择,保留了较为丰富的遗传变异,可用于拓宽栽培稻遗传基础,具有较高的利用潜力和生产应用价值。
我国水稻工作者很早就意识到稻种资源的重要性,并开展了考察和收集工作,目前我国保存的稻种资源数量超过9万份。我国的“稻作之父”丁颖先生早在20世纪20—30年代就开始广泛收集稻种资源,从全国20个省,以及朝鲜、日本、菲律宾、巴西、西里伯岛、爪哇、澳洲、越南共8个国家和地区,收集到7 000多份地方水稻品种,其中多数是农家种,遗传多样性十分丰富,是广东省乃至全国收集最早、极具研究和利用价值的珍贵稻种资源。为了纪念丁颖,卢永根将这批稻种资源命名为“丁氏收集稻种资源(Ting’s rice germplasm collection)”[4]。丁颖在我国还首次利用普通野生稻自然杂交后代于1927—1933年间选育出水稻新品种‘中山一号’,他先后培育出100多个水稻新品种,推广时间超过半个世纪,影响十分深远;并且利用野生稻与栽培稻杂交(印度野生稻和栽培稻‘旱银占’杂交)培育出千粒穗的水稻,引起国内外关注。丁颖先生去世后,卢永根传承保护了丁氏收集稻种资源,并组织在我国南方地区广泛收集野生稻资源,同时还利用不同的渠道从国外引进和收集不同种的野生稻,数量超过2 000份。近20多年来,卢永根带领研究团队,利用已有的资源开展优异基因挖掘和种质创新等研究工作,从中发掘许多优异基因;创制了大规模的水稻染色体单片段代换系(Single segment substitution line, SSSL)和一大批同源四倍体水稻(Autotetraploid rice),并培育出高育性的新型四倍体水稻(Neo-tetraploid rice),解决了同源四倍体水稻育性偏低的“瓶颈”问题。考虑到以上资源具有明显的特色(其中有许多材料是独有的),为了更好地保护与利用这批珍贵的稻种资源,我们将丁颖收集的稻种资源、卢永根组织收集的野生稻及所创制的同源四倍体水稻(含新型四倍体水稻)等种质合并,统一命名为“丁氏稻种资源(Ting’s rice germplasm)”,并以此为基础,对其近20多年来的研究进展,特别是有利基因资源的挖掘、功能基因研究和创新等方面的进展进行全面总结,为在水稻生物育种上利用该宝贵资源提供参考。
1. 丁氏稻种资源研究进展
1.1 丁氏收集稻种资源核心种质的构建与耐逆优异基因资源的挖掘
核心种质能最大程度涵盖较大群体全部遗传背景,具有量小、代表性和多样性等特点,用于优异基因挖掘和遗传改良,可节约人力和物力成本,提高工作效率。李自超等[5]利用国家品种资源库编目入库的50 526份中国地方稻种资源的基本数据,以丁颖分类体系分组、按平方根或对数比例在组内随机取样的策略,构建初级核心种质,总数达4 000份。广东省植物分子育种重点实验室(本室)于2003年开始利用丁氏收集稻种资源中性状数据较齐全的2 262份材料,基于48个表型数据和多种统计方法,构建了“丁氏收集稻种资源”的核心种质库(以下简称“丁氏核心种质”),数量为150份,该群体最大化地保留了原群体的遗传多样性和遗传结构[6]。Zhang等[7]利用274对SSR分子标记对150份丁氏核心种质进行群体结构与连锁不平衡分析,揭示此核心种质资源可用于关联分析。Zhang等[8]利用基因组重测序数据对丁氏核心种质12个重要农艺性状进行全基因组关联分析,鉴定到多个已知关键基因,并筛选出新的关联基因位点。Song等[9]利用SLAF-seq测序,对丁氏核心种质苗期耐冷性进行关联分析,定位了22个QTLs(表1),从新鉴定的QTL区间内筛选出候选新基因Os01g0620100。Fu等[10]对丁氏核心种质纹枯病抗性进行全基因组关联分析,筛选出13个关联显著的QTLs,与前人研究结果一致,典型粳稻‘Early pradifice’对纹枯病抗性最好。Zhang等[11]利用274对SSR分子标记对丁氏核心种质耐铝毒进行关联分析,鉴定出23对标记与耐铝毒相关联。Zhao等[12]利用SLAF-seq测序标记的高密度SNP位点,将丁氏核心种质群体分为2个亚群,利用GWAS方法定位到25个与耐铝相关的QTLs(表1),其中5个QTLs与前人报道的QTLs或克隆的耐铝毒基因共定位。Zhang等[13]对丁氏核心种质进行基因组重测序产生SNP,并鉴定它们的耐铝毒特性,全基因关联分析鉴定到69个候选基因,并利用转录组测序进行验证,其中8个基因与耐铝毒相关(表1)。Zhong等[14]对丁氏核心种质的耐锌毒特性进行全基因组关联分析,以根长为指标,鉴定到7个QTLs,以相对根长为指标,鉴定到19个QTLs,24个QTLs是新发现的(表1)。由上可见,丁氏种质资源具有丰富的优异基因资源,是珍贵的育种材料,可为生物育种和种业创新提供材料基础。
表 1 从丁氏收集稻种资源中鉴定的潜在优异基因资源Table 1. Potential and superior gene resources identified from Ting’s rice germplasm collection基因/QTL1)
Gene/QTL染色体
Chromosome耐逆性
Stress
resistance基因/QTL1)
Gene/QTL染色体
Chromosome耐逆性
Stress
resistance基因/QTL1)
Gene/QTL染色体
Chromosome耐逆性
Stress
resistanceQTL1~QTL3a Chr1 冷害 Os01g57350c Chr1 铝毒 qRE7d Chr11 锌毒 QTL4~QTL13a Chr2 冷害 Os01g57360c Chr1 铝毒 qRRE1~qRRE2d Chr1 锌毒 QTL14~QTL21a Chr3 冷害 Os01g74200c Chr1 铝毒 qRRE4d Chr2 锌毒 QTL22a Chr5 冷害 Os03g30060c Chr3 铝毒 qRRE4~qRRE6d Chr3 锌毒 qALT1.1~qALT1.6b Chr1 铝毒 Os03g30060c Chr3 铝毒 qRRE7d Chr4 锌毒 qALT2.1~qALT2.3b Chr2 铝毒 Os07g03050c Chr7 铝毒 qRRE8~qRRE9d Chr6 锌毒 qALT3.1~qALT3.7b Chr3 铝毒 Os09g33550c Chr9 铝毒 qRRE10d Chr7 锌毒 qALT4.1~qALT4.2b Chr4 铝毒 Os11g03110c Chr11 铝毒 qRRE11~qRRE12d Chr8 锌毒 qALT6.1~qALT6.2b Chr6 铝毒 qRE1~qRE2d Chr2 铝毒 qRRE13d Chr9 锌毒 qALT7.1~qALT7.2b Chr7 铝毒 qRE3~qRE4d Chr6 铝毒 qRRE14d Chr10 锌毒 qALT9.1b Chr9 铝毒 qRE5d Chr8 锌毒 qRRE15~qRRE17d Chr11 锌毒 qALT11.1~qALT11.2b Chr11 铝毒 qRE6d Chr9 锌毒 qRRE18~qRRE19d Chr11 锌毒 1) a~d分别表示鉴定方法为GWAS (SLAF)[9]、GWAS (SLAF)[12]、GWAS (re-squencing、transcriptome)[13]和GWAS (SLAF)[14];QTL5、QTL17分别与OsFAD2、OsMYB2共定位[9],qALT1.6与OsFRDL4、qALT2.2与Os02g49790、qALT3.1与OsApx1、qALT6.2与STAR1共定位[12],qRRE1、qRRE15分别与OsMTI-3a、qZRTDW11共定位[14]
1) a−d indicate the identification methods were GWAS (SLAF)[9], GWAS (SLAF)[12], GWAS (re-squencing, transcriptome)[13] and GWAS (SLAF)[14], respectively; QTL5 and OsFAD2[9], QTL17 and OsMYB2[9], qALT1.6 and OsFRDL4[12], qALT2.2 and Os02g49790[12], qALT3.1 and OsApx1[12], qALT6.2 and STAR1[12], qRRE1 and OsMTI-3a[14], qRRE15 and qZRTDW11[14], are co-located1.2 丁氏收集稻种资源育性相关基因研究进展
水稻籼粳亚种间杂种具有强大的生物学优势,但杂种F1普遍高度不育,难以直接利用。籼粳亚种间杂种F1不育主要表现为花粉和胚囊部分不育,花粉育性“中性基因”和胚囊“广亲和基因”可以部分克服杂种不育性。Shahid等[15]通过测交和分子标记鉴定发现,2份栽培稻(DN18和DN75)携带San和Sbn双中性基因,而DN22有San、Sbn和Scn 3个中性基因。杨有新等[16]利用国家水稻微核心种质和丁氏稻种资源等,通过胚囊中性基因S5n特异性引物扩增,结合测序鉴定发现,‘毫补卡’‘小红谷’‘老造谷’‘三磅七十箩’‘木邦谷’‘魔王谷内杂’‘饭毫皮’‘飞蛾糯2’‘包协−7B’和‘特青选恢’10个品种携带S5n中性基因。李宏岩等[17]将携带S5n的水稻种质分别与‘台中65’及携带花粉不育基因的一套近等基因系组配F2群体,利用Sa、Sb和Sc座位紧密连锁的分子标记进行基因分型,发现‘灰背子’和‘Madhukar’同时携带S5n、San和Sbn中性基因,‘饭毫皮’‘秕五升’和‘粤泰B’携带S5n和Sbn中性基因,‘Jackson’携带S5n和Scn中性基因。这些材料是克服籼粳杂种F1不育性的重要基因来源。
2. 丁氏稻种资源有利基因的挖掘
2.1 野生稻耐逆优异基因资源挖掘
丁氏稻种资源中的野生稻资源主要是卢永根组织收集的野生稻,其中国内的普通野生稻主要来自广东高州、增城、遂溪、佛冈、惠来、博罗、从化,江西东乡,海南琼海,湖南茶陵、江泳,广西贵港和玉林等地;药用野生稻主要来自广西;疣粒野生稻主要来自海南。本室对这些野生稻开展了一系列研究发现,它们蕴含抗旱、抗寒、耐铝毒以及克服杂种F1不育性的中性基因等,并从遂溪普通野生稻中克隆了2个调控水稻粒型的基因。
药用野生稻原产中国,蕴含大量抗白叶枯病、稻瘟病等抗病基因,抗褐飞虱、白背飞虱等抗虫基因和耐旱、耐盐、耐热等耐非生物胁迫基因。另外,药用野生稻的外观品质好且蛋白质含量高,是水稻品种改良的重要种质资源[18]。但药用野生稻的基因型为CC组,与栽培稻AA组的亲缘关系相差较大,通过有性杂交方法很难得到杂种。为了有效利用药用野生稻,刘耀光团队构建了世界首个药用野生稻TAC文库,该文库有20 631个克隆,基本覆盖了药用野生稻全基因组,为药用野生稻有利基因的转移和应用提供了重要的基础[19]。汪暖等[20]成功建立了以‘粤香占’‘华粳籼74’为受体的药用稻TAC克隆的籼稻转化体系,为药用野生稻TAC大片段基因成功转移创造了条件。刘蕊等[21]和Liu等[22]以AP2/EREBP、bZIP2和NAC转录因子保守序列为探针,对该药用野生稻文库进行耐旱基因筛选,并将筛选到的克隆以‘华粳籼74’为受体进行遗传转化,获得转化植株,发现其中部分转基因材料具有抗旱性。李培纲等[23]从药用野生稻中克隆到OoADF1基因,并证明OoADF1对高盐胁迫具有一定的响应能力。
高州野生稻是广东境内分布面积最广的普通野生稻,具有丰富的遗传多样性,是水稻育种的宝贵资源[24]。王兰等[25]以极不耐寒的品种‘三百粒’为对照,通过自然低温处理以及光照培养箱变温处理鉴定到高州野生稻2个株系GZW5和GZW18具有强耐寒性。刘向东利用普通野生稻自交后代,选育稳定的野生稻品系,先后在江西东乡和广东遂溪普通野生稻自交后代中选育出4个稳定的品系,分别是‘华野1号’‘华野2号’‘华野3号’和‘华野4号’,这些品系均保留了野生稻的高抗性,具有利用价值。Liu等[26]对‘华野1号’和‘华野2号’进行基因组重测序,发现NBS-LRR保守域在第11号染色体上展示更多的变异;在‘华野1号’和‘华野2号’中,NBS比LRR的保守性更强;‘华野1号’的NBS基因比‘华野2号’的具有更高水平的遗传多样性。Yu等[27]利用基因组重测序分析,在‘华野3号’中找到194个NBS-LRR变异编码基因,这些基因分布于水稻12条染色体,其中28个变异NBS-LRR基因对基因功能和表型变异产生明显影响。这些结果为水稻抗性育种提供了重要的基础。
2.2 野生稻胚囊和花粉中性基因挖掘
籼粳杂交组配杂种F1具有强大的生物学优势和增产潜力,但育性偏低,阻碍了其在生产上的进一步利用。广亲和基因S5n和杂种花粉育性中性基因的发现和成功克隆为克服籼粳杂种不育创造了条件[28]。魏常敏等[29]利用S5n功能性标记,对来自国内不同地区的441份普通野生稻进行检测和鉴定,发现其中18份材料含有S5n基因(表2)。Tong等[30]认为S5n广亲和基因在野生稻与栽培稻中平行进化。Peng等[31]对134份栽培稻和139份普通野生稻(来自琼海、高州、佛冈、增城、隧溪等)的S5n基因进行分子进化研究,发现籼稻、部分普通野生稻和‘南京11’(对照)聚成一类,粳稻、其余的普通野生稻和‘巴利拉’(对照)聚成一类,琼海普通野生稻单独聚成一类,研究结果表明S5n是一个非常保守的基因,籼亚种、粳亚种分别起源于不同的野生稻祖先种(表2)。史磊刚等[32]以粳稻‘台中65’及其Sb座位的近等基因系为母本,以141份高州野生稻中的12份材料(GZW005、006、011、019、034、075、087、099、101、124、133、137)为父本,组配杂交组合得到F1和F2,成功鉴定到GZW099中携带Sb座位花粉育性中性基因。Liu等[33]以粳稻‘台中65’及其Sd、Se座位的近等基因系为母本,以141份高州野生稻的13份材料(GZW006、009、011、013、026、054、060、099、101、133、135、136、137)为父本,组配杂交组合得到F1和F2,成功鉴定到GZW054中携带Sd、Se座位花粉育性中性基因(表2)。Li等[34]以普通野生稻为父本,与粳亚种‘台中65’和籼亚种‘广陆矮4号’分别杂交,构建F2群体,检测花粉5个座位(Sa、Sb、Sc、Sd、Se)的育性中性基因,发现普通野生稻中存在Sb、Sd、Se的中性基因(表2)。这些蕴涵中性基因的普通野生稻可为克服籼粳杂种不育提供有价值的基因资源。
表 2 丁氏稻种资源野生稻资源的有利基因资源Table 2. Favorable gene resources from wild rice of Ting’s rice germplasm基因/QTL
Gene/QTL染色体
Chromosome功能
Function来源
Source鉴定方法
Identification methodOoADF1 耐盐 药用野生稻 同源克隆 Sbn Chr5 克服籼粳杂种花粉不育 广东高州普通野生稻以及IRW28 分子标记 Sdn Chr1 克服籼粳杂种花粉不育 广东高州普通野生稻以及IRW28 分子标记 Sen Chr12 克服籼粳杂种花粉不育 广东高州普通野生稻以及IRW28 分子标记 qRRE-6-2 Chr6 耐铝 广东高州普通野生稻 QTL定位 qRRE-7-2 Chr7 耐铝 广东高州普通野生稻 QTL定位 qGL3.5 Chr3 调控粒型 广东遂溪普通野生稻 基因定位与克隆 GSW3 Chr3 调控粒型 广东遂溪普通野生稻 基因定位与克隆 2.3 普通野生稻及其与栽培稻杂种的结实率偏低的细胞学研究
普通野生稻的结实率普遍偏低。杨培周等[35]对广东高州6个地点共141份普通野生稻成熟胚囊和胚囊发育特点等进行研究,发现在141份供试材料中,胚囊异常频率平均11.11%,最高达67.86%,并认为胚囊发育异常是其结实率偏低的原因之一。普通野生稻是栽培稻的直接祖先种,但是普通野生稻与栽培稻杂种的结实率并不高。练子贤等[36]将粳型测验种‘台中65’和籼型测验种‘广陆矮4号’分别与不同编号的高州野生稻进行杂交组配,研究杂种的花粉育性、胚囊育性,对60个杂交组合的6 618个胚囊进行观察,发现胚囊结构正常率平均为64.61%,最高达97.00%;有些组合育性较低,最低仅为20.00%,胚囊育性偏低是影响栽培稻与野生稻杂种的结实率偏低的原因之一。
2.4 非AA组野生稻与栽培稻的杂种不育的细胞学研究
高秆野生稻Oryza alta是水稻抗病育种的一种重要种质资源,为异源四倍体(CCDD),与栽培稻(AA)亲缘关系较远,通过常规有性杂交方式进行有利基因转移很困难。高秆野生稻多倍化后,再与栽培稻杂交可能是克服种间隔离的一种有效方法。刘向东将高秆野生稻进行辐射诱变后,在自交后代选育出育性明显提高、感光性和落粒性显著减弱的稳定品系‘华野5号’。Zhang等[37]利用‘华野5号’建立适合的组培体系,通过秋水仙碱处理愈伤组织,成功获得同源异源八倍体高秆野生稻新种质,为利用高秆野生稻有利基因提供了基础。Fu等[38]通过对高秆野生稻与栽培稻杂种进行研究,发现“杂种不活”(Hybrid inviability)是其种间生殖隔离的主要原因。短花药野生稻Oryza brachyantha(FF)是基因组最小的一个稻种,基因组分化程度高且相对稳定,在稻属进化与分类研究中具有重要的地位,与栽培稻基因组(AA)亲缘关系较远,通过常规的有性杂交途径很难获得杂种。傅雪琳等[39]利用激光扫描共聚焦显微术对栽培稻与短花药野生稻杂种的胚胎、胚乳以及胚囊的发育过程进行了观察,发现四分体异常、单核或二核退化是杂种胚败育的主要原因。为克服栽野杂种的生殖障碍,Fu等[40]对药用野生稻和栽培稻杂种授粉前后进行了深入的细胞学研究,从胚囊发育角度揭示了杂种败育的细胞学机理。
2.5 野生稻有利基因(QTL)定位与克隆
铝是地壳中含量最丰富的金属元素,对植物根系的毒害作用很大。褚绍尉等[41]对高州普通野生稻进行耐铝性鉴定,发现其中8份材料具有很强的耐铝性;然后利用分布于水稻12条染色体上的193个SSR标记对高州普通野生稻的耐铝基因进行QTL定位,初步定位到2个耐铝QTLs(qRRE-6-2与qRRE-7-2),分别位于第6和7号染色体上(表2),对表型的贡献率分别为18.33%与9.18%。王兰等[42]以少分蘖的高秆栽培稻‘南特号’为母本、普通野生稻品系‘华野3号’为父本,构建F2分离群体,共检测到33个与株高相关的QTLs、19个与分蘖相关的QTLs,其中1个与株高相关的主效QTL定位于水稻第1号染色体标记RM30—104之间,来源于‘华野3号’,这些研究结果为水稻株高与分蘖基因的克隆提供了理论基础,为水稻分子育种提供了种质资源。
郑跃滨等[43]以长粒栽培稻‘KJ01’为母本、‘华野3号’(短粒)为父本构建F2分离群体,进行粒长QTL连锁遗传分析,共检测到24个与粒长相关的QTLs,其中位于第3号染色体PSM379—RID24455、RID24455—RM15689和RM571—16238区间的3个主效QTLs对表型的贡献率分别为54.85%、31.02%和7.62%,其中标记PSM379—RID24455的主效QTL为新发现的主效QTL位点,来源于‘华野3号’。Wang等[44]利用BSA-seq对该粒长基因进行重测序定位,把粒长基因定位到第3号染色体340 kb区段内,进一步精细定位将目的基因缩小到24.0 kb范围内,内含目的基因ORF18和ORF17的部分编码区,通过CRISPR/Cas9编辑分析确定ORF18为粒型目的基因,并命名为“qGL3.5”(表2),该基因通过促进籽粒外颖纵向细胞伸长、横向细胞数量增加来增加籽粒大小。Bai等[45]对BSA关联区域内的37个与性状相关的基因进行分析,发现内含1个基因ORF25编码GTPase结合蛋白,利用CRISPR/Cas9对ORF25进行基因功能研究,发现ORF25负调控水稻粒型,并命名为“GSW3”(表2);GSW3在‘华野3号’中过量表达,GSW3Huaye3转化到‘KJ01’,转化植株籽粒变短;2个敲除突变体KO-1和KO-2能增加粒长20.16%和14.05%、增加粒宽6.7%和4.6%,GSW3通过促进籽粒外颖纵向细胞伸长、横向细胞数量增加来调控籽粒大小;GSW3负调控水稻粒型效果明显,在水稻生产上具有较大的利用潜力。
3. 丁氏稻种资源的创新研究
丁氏稻种资源的创新研究主要包括2个方面:一是创建同源四倍体水稻和培育新型四倍体水稻;二是构建水稻染色体SSSL。从1998年开始,本室以丁氏稻种资源栽培稻为基础,通过秋水仙碱处理进行染色体加倍,创建了一大批同源四倍体水稻,一方面利用该四倍体水稻开展原种及其杂种低育性的细胞学和分子遗传学研究;另一方面,通过不同类型同源四倍体水稻杂交,在高代成功选育出高育性的新型四倍体水稻,解决了同源四倍体水稻育性偏低的“瓶颈”问题。
3.1 同源四倍体水稻低育性的细胞学和分子遗传学研究
同源四倍体水稻每个同源组均含有4条染色体,基因剂量增加1倍,基因组原有的协调关系被打破,势必影响生殖发育过程。本室研究表明,同源四倍体水稻及其杂种F1的花粉发育、胚囊发育和双受精等过程均出现不同程度的异常,它们都可能影响育性并导致育性降低[46-49]。影响同源四倍体水稻花粉和胚囊发育异常的主要原因是减数分裂染色体行为异常以及相关基因表达的异常。He等[50]和Wu等[51]研究发现,同源四倍体水稻及其杂种F1花粉母细胞减数分裂前期I均会出现单价体和三价以上的多价体,导致同源染色体异常分离;在中期I和后期I,出现纺锤体位置和形态异常、染色体拖曳和落后等异常;末期I和末期II出现微核和分裂不同步等异常现象;另外,减数分裂期间的微管骨架组织也存在许多异常现象。同源四倍体水稻及其杂种F1胚囊败育类型主要有胚囊退化,雌性生殖单位退化、卵器退化、极核位置和数目异常、小胚囊及其他异常等,这些异常胚囊均无法正常受精,导致结实率下降[52]。
同源四倍体水稻花粉和胚囊发育过程中许多重要基因以及非编码RNA均出现表达异常。Wu等[51]、Chen等[53]和Li等[54]利用不同的同源四倍体水稻材料研究发现,与二倍体水稻原种比较,同源四倍体水稻花粉母细胞减数分裂期间出现不同程度差异表达的基因,其中有些与育性密切相关,如减数分裂相关基因PAIR2和OsDMC1B等(表3)。此外,Li等[55-57]还发现同源四倍体水稻中的基因组DNA碱基变异及甲基化水平异常,miRNAs和lncRNAs异常表达可能也是其育性低的原因。
表 3 同源四倍体水稻减数分裂期间差异表达基因Table 3. Differently expressed genes during meiosis of autotetraploid rice3.2 新型四倍体水稻创制及高育性机理研究
为解决同源四倍体水稻育性偏低的问题,刘向东团队利用不同类型的同源四倍体水稻杂交,并经过多代自交和选择,成功选育出高育性四倍体水稻,并命名为“新型四倍体水稻”[58]。新型四倍体水稻不仅育性高,结实率可达80%,而且,还具备其他优点,包括携带广亲和基因S5n和杂种花粉育性的中性基因(Scn)等多种优异基因,基因组中存在丰富的DNA变异;类型独特,与籼型同源四倍体水稻杂交后代F1的产量优势十分明显,且可保持多代等[59]。目前发现可能与新型四倍体水稻高育性相关的基因至少有7个[59],包括NY1、NY2、MOF1a、HSP101-1、kin7l、bzr3和nrfg4。新型四倍体水稻与同源四倍体水稻杂交强优势的分子机理复杂,可能涉及重要功能基因和表观遗传因子的差异表达,如减数分裂相关基因(DPW和CYP703A3等)、糖代谢和淀粉合酶相关基因(OsBEIIb和OsSSIIIa等)、杂种优势相关基因(GW8、OsGA20ox1、Ghd8、GW6a、LP1、Hd1、IPA1和NAL1)等在F1中均上调表达[60-61]。
3.3 水稻染色体单片段代换系构建与功能基因研究
张桂权团队从1998年开始,以自主培育的水稻新品种‘华粳籼74’为受体,分别以28个不同来源和类型的栽培稻以及8个AA基因组的野生稻为供体构建包含2 360份水稻的SSSL文库[62]。另外,本室还以优良品种‘粤香占’为受体亲本、高州野生稻GZW087为供体亲本,获得了20份高州野生稻种质的9个SSSLs;以‘华粳籼74’为受体亲本、以高州野生稻的18份种质为父本进行杂交和回交,构建了7个SSSLs和3个双片段代换系。利用以上材料克隆了多个优异基因,Lin等[63]利用以展颖野生稻Oryza glumaepatula为供体亲本的SSSL克隆了1个能够显著提升稻米外观品质(粒大小和垩白度)的基因GL9。Zhao等[64]利用以‘美国茉莉稻’(‘American Jasmine’)为供体亲本的SSSL克隆了1个控制水稻籽粒大小和株型的基因GS6.1。Zhan等[65]利用SSSL克隆了控制水稻粒长的基因GL10,该基因编码1个MADS-box家族转录因子OsMADS56,为正调控水稻粒长的新基因。Zhan等[66]利用SSSL克隆了调控水稻籽粒大小和数量的基因GW10。Pei等[67]利用以4种AA基因组野生稻为供体亲本的SSSL鉴定出控制种子低温萌发的10个QTLs。Tan等[68]利用以展颖野生稻为供体亲本的SSSL鉴定获得2个控制水稻柱头外露率的新QTLs。Wang等[69-70]分别克隆了控制水稻粒长和粒宽的基因GW7,以及控制水稻籽粒大小、形状和品质的基因GW8。此外,本室赵杏娟[71]还利用以高州野生稻为供体的SSSL鉴定出7个重要农艺性状的QTLs(表4)。
表 4 利用高州野生稻单片段代换系检出的重要农艺性状QTLsTable 4. QTLs of important agronomic trait detected by SSSL of wild rice in Gaozhou代换系编号
SSSL codeQTL 性状
Trait染色体
Chromosome定位片段
Located segment加性效应
Additive
effect加性效应比例/%
Additive effect
proportion2 Pss-2-2 结实率 Chr2 RM530—RM208—长臂末端 −0.345 37.5 5 Pss-11-1 结实率 Chr11 RM21—PSM366—PSM417 −0.305 33.1 6 Lfl-6-1 剑叶长 Chr6 RM253—RM527—RM539—RM3—PSM138 −8.165 25.3 6 Wfl-6-1 剑叶宽 Chr6 RM253—RM527—RM539—RM3—PSM138 −7.865 24.3 9 Lfl-2-1 剑叶长 Chr2 RM110—RM211—RM279—PSM116 −0.340 16.3 9 Wfl-2-1 剑叶宽 Chr2 RM110—RM211—RM279—PSM116 −0.490 23.6 4. 展望
种子是农业的“芯片”,是国家粮食安全的命脉。中央高度重视种质资源保护与利用工作,2021年中央一号文件提出了“打好种业翻身仗”,中央全面深化改革委员会通过的《种业振兴行动方案》强调,要加强基础性前沿性研究,加快实施农业生物育种重大科技项目,开展种源关键核心技术攻关。水稻作为我国最重要的粮食作物,绿色革命基因和杂种优势利用确保我国稻米产量保持在较高水平。绿色超级稻育种、水稻轻简栽培技术及环境适应型新品种的培育需要挖掘更多的水稻有利基因资源。丁氏稻种资源是我国目前保存的资源中比较有特色的一类,特别是农家种、普通野生稻和新型四倍体水稻,以及以自主品种为受体、野生稻作为供体构建的水稻染色体SSSLs等是独有的,这些珍贵的资源蕴藏丰富的优异基因资源,至今已提供给国内30多个科研单位或高等院校开展研究,取得重要进展,发表一批高水平的学术论文。可以相信,丁氏稻种资源未来将在水稻生物育种中发挥越来越大的作用。本室下一步将结合“岭南水稻种质资源基地库”建设,大力开展丁氏稻种资源氮高效、耐旱、耐盐、耐冷和强抗病虫性等优异基因的发掘,进一步通过遗传学、分子生物学和多组学等技术,揭示优异基因的作用机理,获得基因知识产权,在振兴种业和实现国家未来粮食安全方面作出贡献。
-
![]()
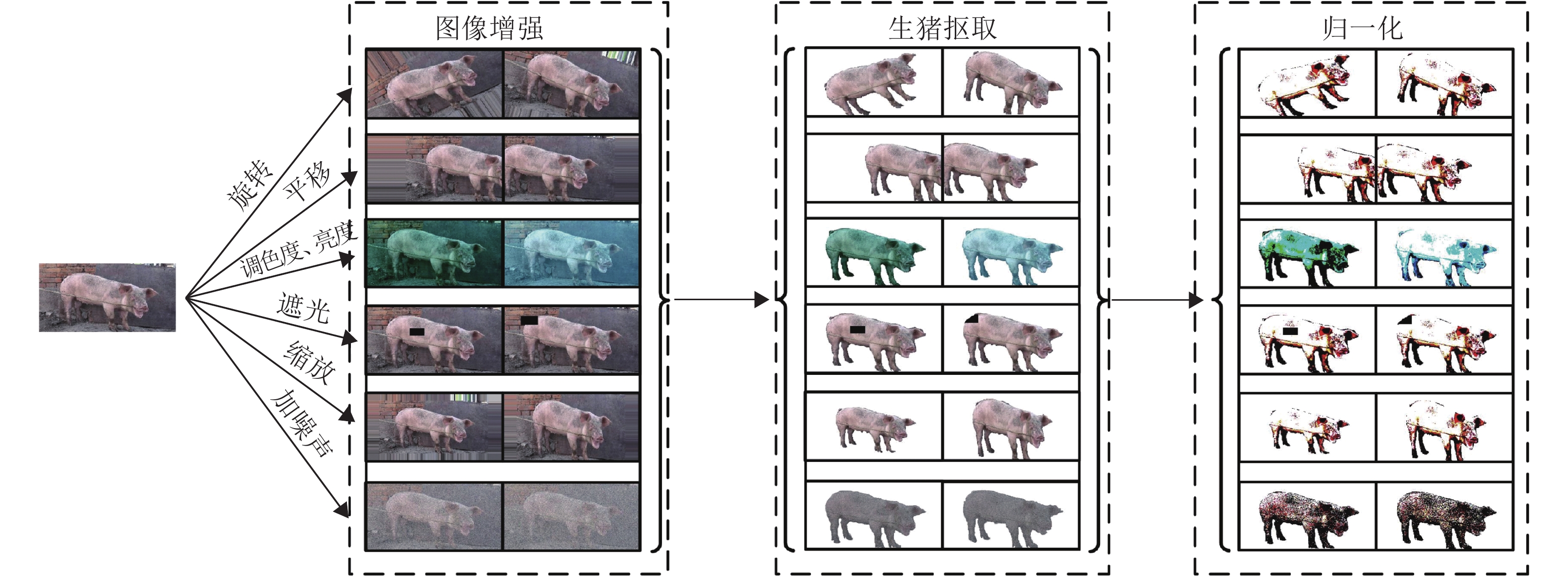
图 1 VGG-UNET模型输入输出数据集创建过程
Figure 1. The process of establishing input and output datasets for the VGG-UNET model
![]()
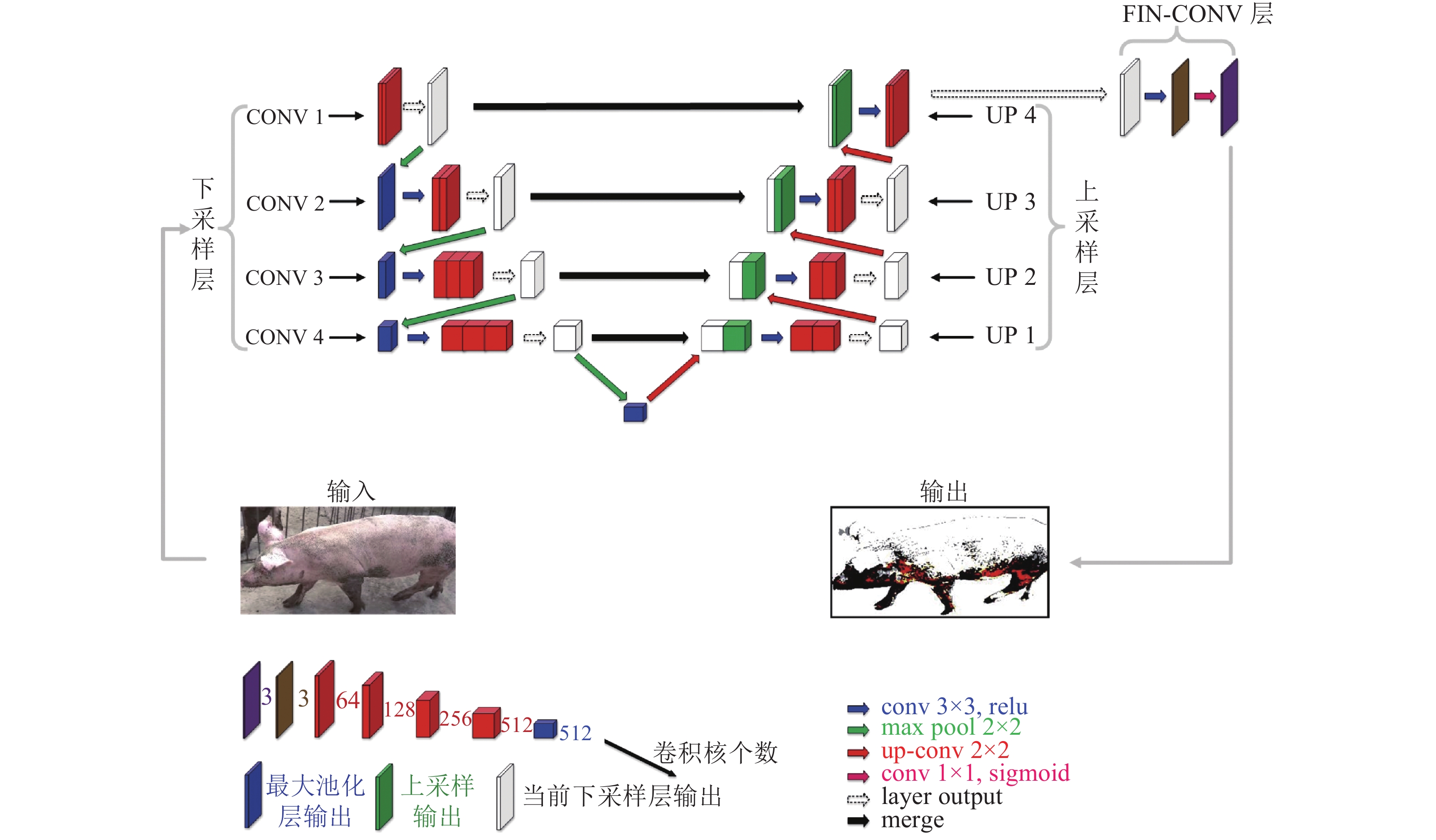
图 2 VGG-UNET网络结构图
图中conv 3×3,relu中conv表示卷积操作,3×3表示卷积核大小,relu表示激活函数;conv 1×1,sigmoid中conv表示卷积操作,1×1表示卷积核大小,sigmoid表示激活函数;up-conv 2×2中up-conv表示反卷积操作,2×2表示反卷积核大小;max pool 2×2中maxpool表示最大池化操作,2×2表示池化核大小;layer output表示当前卷积层级输出;merge表示融合与上采样层处于同一层的下采样层输出操作
Figure 2. Diagram of the VGG-UNET network structure
![]()
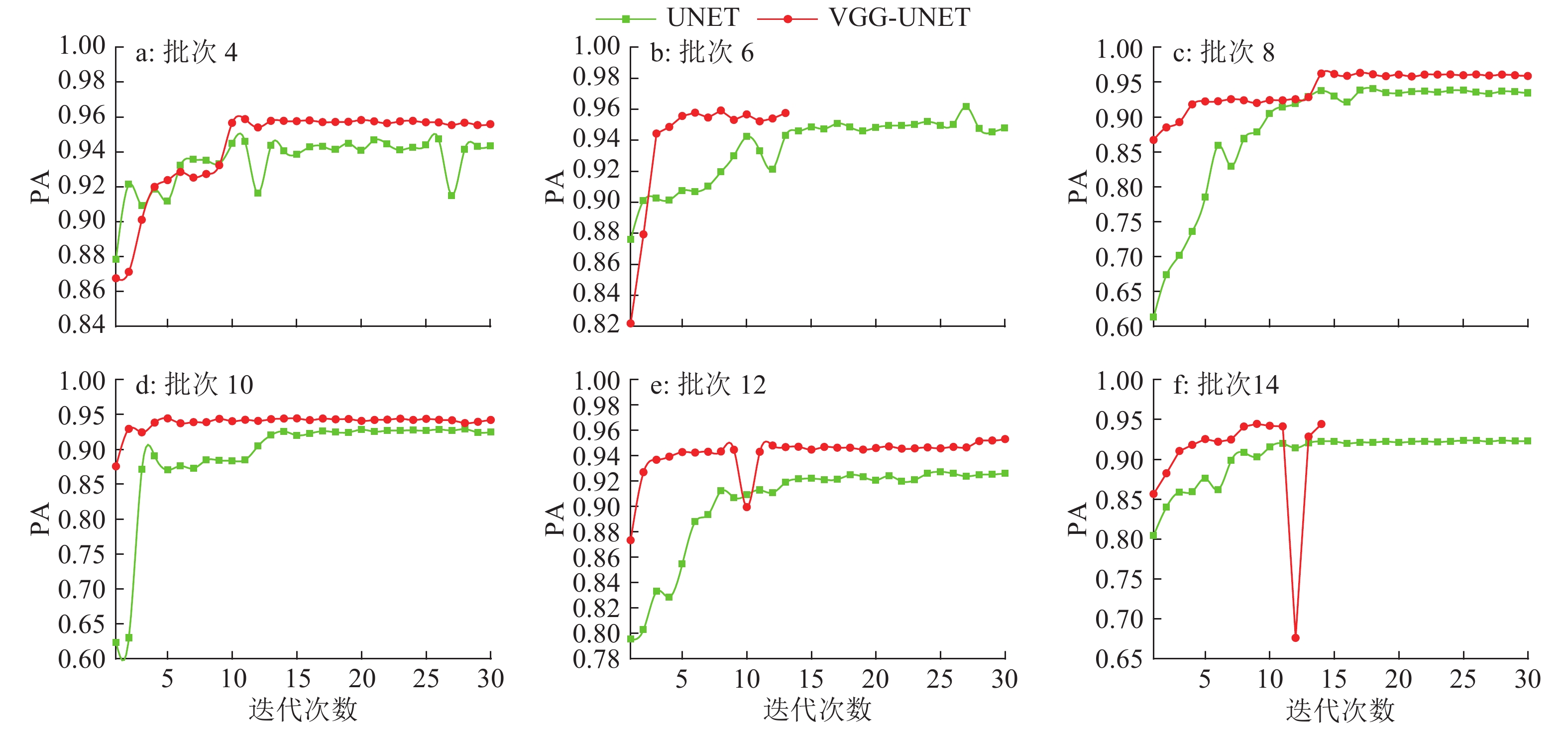
图 3 UNET与VGG-UNET模型不同批大小下每轮PA变化
Figure 3. The variations for each round of PA in different batch sizes of UNET and VGG-UNET models
![]()
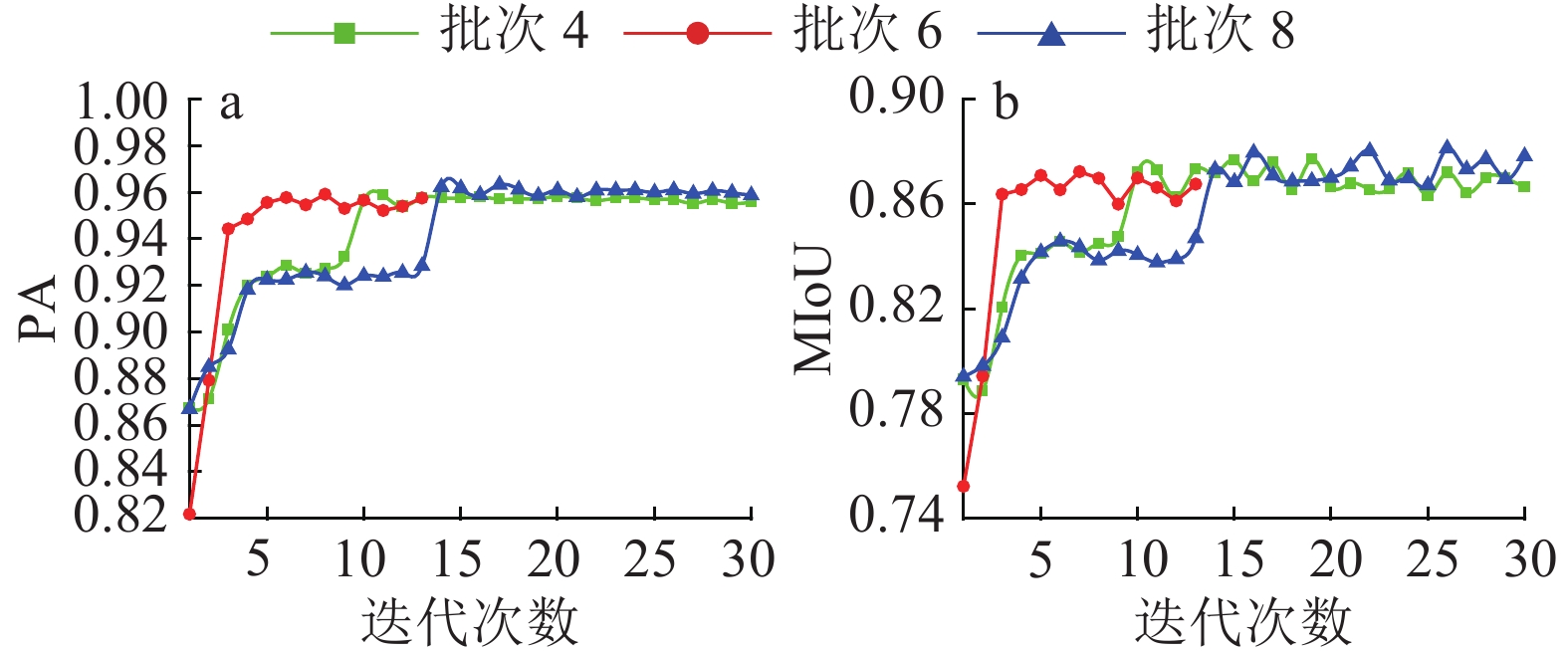
图 4 VGG-UNET模型不同批大小下每轮PA及MIoU变化
Figure 4. The variations for each round of PA and MIoU in different batch sizes of VGG-UNET model
![]()
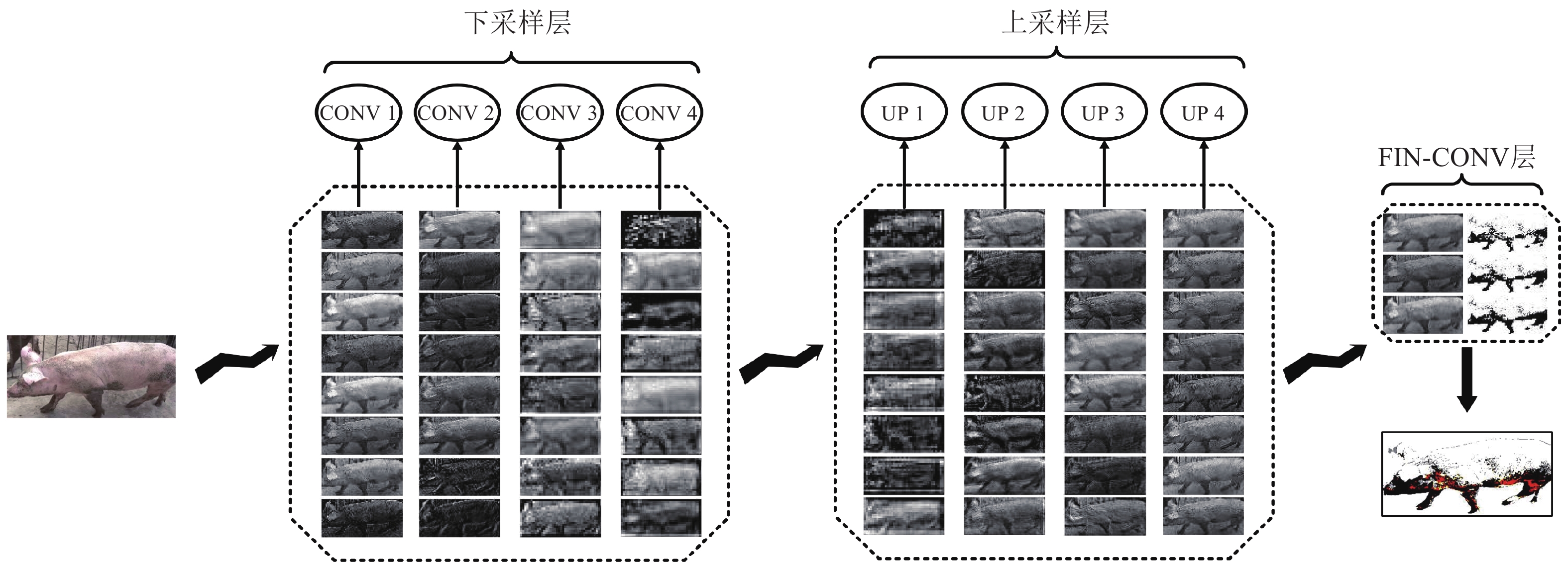
图 5 VGG-UNET模型中间层可视化
上、下采样层中每一列仅展示卷积层或反卷积层中随机的8个卷积核输出
Figure 5. Visualization of the middle layer of the VGG-UNET model
![]()
图 6 VGG-UNET模型预测结果的可视化
a~f组图片分别为不同图片的生猪轮廓提取结果;每组图片的第1列表示模型输入,第2列表示通过预处理后的真实输出,第3列表示批大小为8的VGG-UNET模型预测结果
Figure 6. Visualization of prediction results from the VGG-UNET model
表 1 不同批大小下模型性能比较
Table 1 The performance comparison of models with different batch sizes
模型 批次 PA MPA MIoU FWIoU UNET 4 0.934 3 0.868 0 0.846 4 0.860 7 6 0.932 7 0.863 8 0.849 3 0.858 1 8 0.883 9 0.857 2 0.817 9 0.838 9 VGG-UNET 4 0.940 3 0.877 6 0.864 2 0.866 5 6 0.941 8 0.874 1 0.864 9 0.864 1 8 0.943 2 0.885 1 0.866 0 0.851 5  下载: 导出CSV
下载: 导出CSV
-
[1] 杜晓冬, 滕光辉, 李卓, 等. 典型猪舍光环境下机器视觉图像量化评价及筛选[J]. 农业工程学报, 2017, 33(2): 213-219. [2] 蔡一欣, 马丽, 刘刚. 奶牛隐性乳房炎便携式计算机视觉快速检测系统设计与试验[J]. 农业工程学报, 2017, 33(增刊1): 63-69. [3] 蔡骋, 宋肖肖, 何进荣. 基于计算机视觉的牛脸轮廓提取算法及实现[J]. 农业工程学报, 2017, 33(11): 171-177. doi: 10.11975/j.issn.1002-6819.2017.11.022 [4] 何东健, 刘冬, 赵凯旋. 精准畜牧业中动物信息智能感知与行为检测研究进展[J]. 农业机械学报, 2016, 47(5): 231-244. [5] 肖德琴, 冯爱晶, 杨秋妹, 等. 基于视频追踪的猪只运动快速检测方法[J]. 农业机械学报, 2016, 47(10): 351-357. doi: 10.6041/j.issn.1000-1298.2016.10.045 [6] 刘同海, 李卓, 滕光辉, 等. 基于RBF神经网络的种猪体重预测[J]. 农业机械学报, 2013, 44(8): 245-249. [7] 纪滨, 朱伟兴, 刘波, 等. 基于脊腹线波动的猪呼吸急促症状视频分析[J]. 农业工程学报, 2011, 27(1): 191-195. doi: 10.3969/j.issn.1002-6819.2011.01.031 [8] 闫丽, 沈明霞, 谢秋菊, 等. 哺乳母猪高危动作识别方法研究[J]. 农业机械学报, 2016, 47(1): 266-272. [9] 刘龙申, 沈明霞, 柏广宇, 等. 基于机器视觉的母猪分娩检测方法研究[J]. 农业机械学报, 2014, 45(3): 237-242. [10] CAO X, WEI Y, WEN F, et al. Face alignment by explicit shape regression[J]. Int J Comput Vision, 2012, 107(2): 2887-2894.
[11] WU S, KAN M, HE Z, et al. Funnel-structured cascade for multi-view face detection with alignment-awareness[J]. Neurocomputing, 2017, 221: 138-145. doi: 10.1016/j.neucom.2016.09.072
[12] 张志刚, 周明全, 耿国华. 人脸关键特征点自动标定研究[J]. 计算机工程与应用, 2007, 43(21): 197-198. doi: 10.3321/j.issn:1002-8331.2007.21.060 [13] 孙龙清, 李玥, 邹远炳, 等. 基于改进Graph Cut算法的生猪图像分割方法[J]. 农业工程学报, 2017, 33(16): 196-202. doi: 10.11975/j.issn.1002-6819.2017.16.026 [14] 朱伟兴, 纪滨, 秦锋. 基于伪球算子边缘模型的猪前景帧检测[J]. 农业工程学报, 2012, 28(12): 189-194. doi: 10.3969/j.issn.1002-6819.2012.12.031 [15] GUO Y, ZHU W, JIAO P, et al. Foreground detection of group-housed pigs based on the combination of mixture of Gaussians using prediction mechanism and threshold segmentation[J]. Biosyst Eng, 2014, 125(3): 98-104.
[16] 刘波, 朱伟兴, 霍冠英. 生猪轮廓红外与光学图像的融合算法[J]. 农业工程学报, 2013, 29(17): 113-120. doi: 10.3969/j.issn.1002-6819.2013.17.015 [17] 马丽, 纪滨, 刘宏申, 等. 单只猪轮廓图的侧视图识别[J]. 农业工程学报, 2013, 29(10): 168-174. [18] 王林, 董楠. 基于Gabor特征与卷积神经网络的人体轮廓提取[J]. 南京理工大学学报, 2018, 42(1): 89-95. [19] LONG J, SHELHAMER E, DARRELL T. Fully convolutional networks for semantic segmentation[C]// IEEE. IEEE Conference on Computer Vision and Pattern Recognition. San Francisco: IEEE, 2015: 3431-3440.
[20] KRIZHEVSKY A, SUTSKEVER I, HINTON G E. ImageNet classification with deep convolutional neural networks[J]. Commun ACM, 2017, 60(6): 84-90. doi: 10.1145/3098997
[21] RUSSAKOVSKY O, DENG J, SU H, et al. ImageNet large scale visual recognition challenge[J]. Int J Comput Vision, 2015, 115(3): 211-252. doi: 10.1007/s11263-015-0816-y
[22] SZEGEDY C, LIU W, JIA Y, et al. Going deeper with convolutions[C]//IEEE. IEEE Conference on Computer Vision and Pattern Recognition. San Francisco: IEEE, 2015: 1-9.
[23] 杨阿庆, 薛月菊, 黄华盛, 等. 基于全卷积网络的哺乳母猪图像分割[J]. 农业工程学报, 2017, 33(23): 219-225. doi: 10.11975/j.issn.1002-6819.2017.23.028 [24] 赵凯旋, 何东健. 基于卷积神经网络的奶牛个体身份识别方法[J]. 农业工程学报, 2015, 31(5): 181-187. doi: 10.3969/j.issn.1002-6819.2015.05.026 [25] RONNEBERGER O, FISCHER P, BROX T. U-net: Convolutional networks forbiomedical image segmentation[C]// MICCAI. International Conference on Medical Image Computing and Computer-Assisted Intervention. Munich: MICCAI, 2015: 234-241.
[26] GARCIA-GARCIA A, ORTS-ESCOLANO S, OPREA S, et al. A survey on deep learning techniques for image and video semantic segmentation[J]. Appl Soft Comput, 2018, 70: 41-65. doi: 10.1016/j.asoc.2018.05.018
[27] ISOLA P, ZHU J Y, ZHOU T, et al. Image-to-Image translation with conditional adversarial networks[C]//IEEE. IEEE Conference on Computer Vision and Pattern Recognition. Honolulu: IEEE, 2017: 5967-5976.

计量
- 文章访问数: 1609
- HTML全文浏览量: 2
- PDF下载量: 1811
