
Effect of ethylene and 1-MCP on the terpenoid fragrance of Hedychium coronarium and the expressions of related genes
-
摘要:目的
研究乙烯和1-MCP对白姜花萜类香气物质合成释放的影响,探究相关基因的表达变化。
方法以白姜花Hedychium coronarium为材料,分别用10 μL·L–1的乙烯和4 μL·L–1的乙烯抑制剂1-MCP处理8 h,随后采用顶空固相微萃取和气相色谱–质谱(GC-MS)技术进行挥发性香气物质的测定,所得到的离子图用NIST 08数据库进行定性及定量分析。通过荧光定量PCR分析相关基因的表达情况。
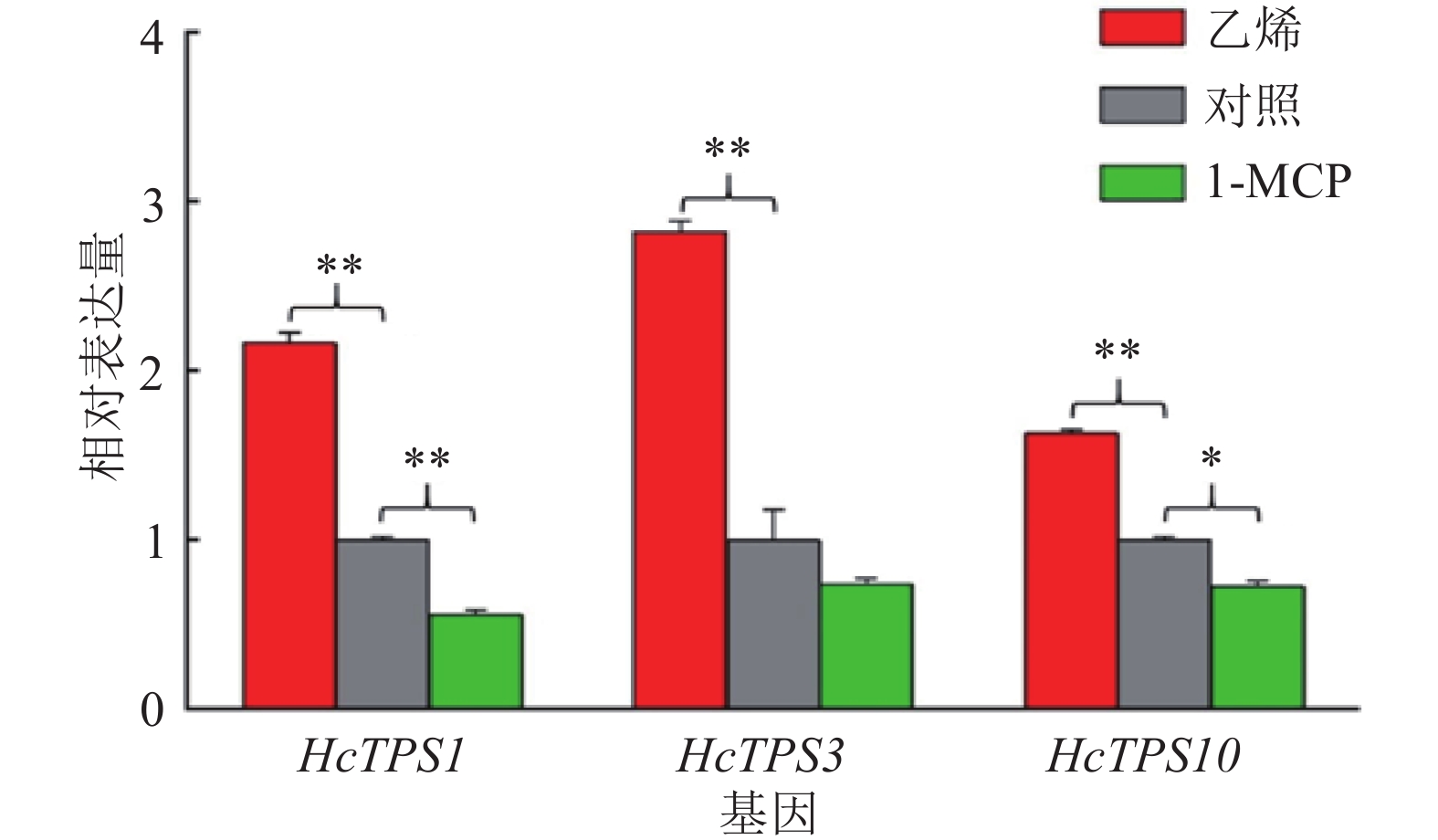
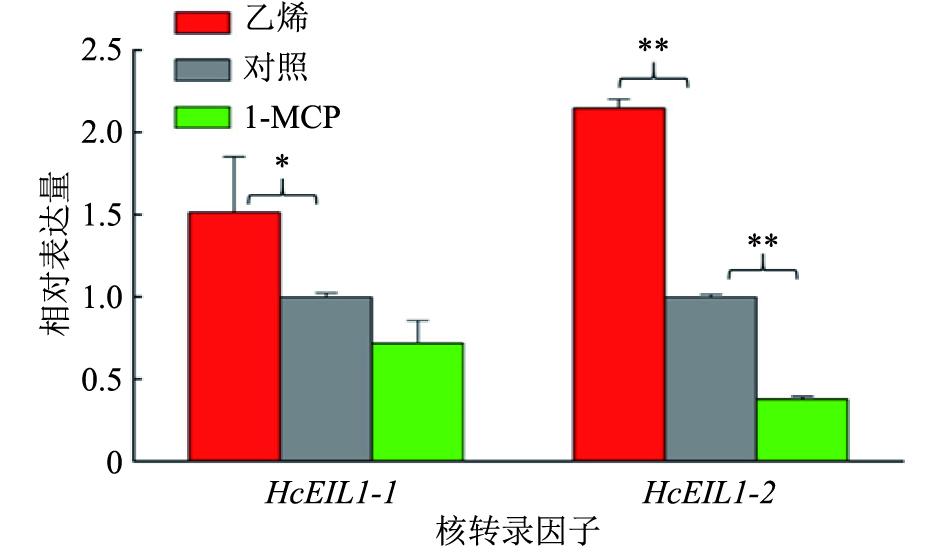
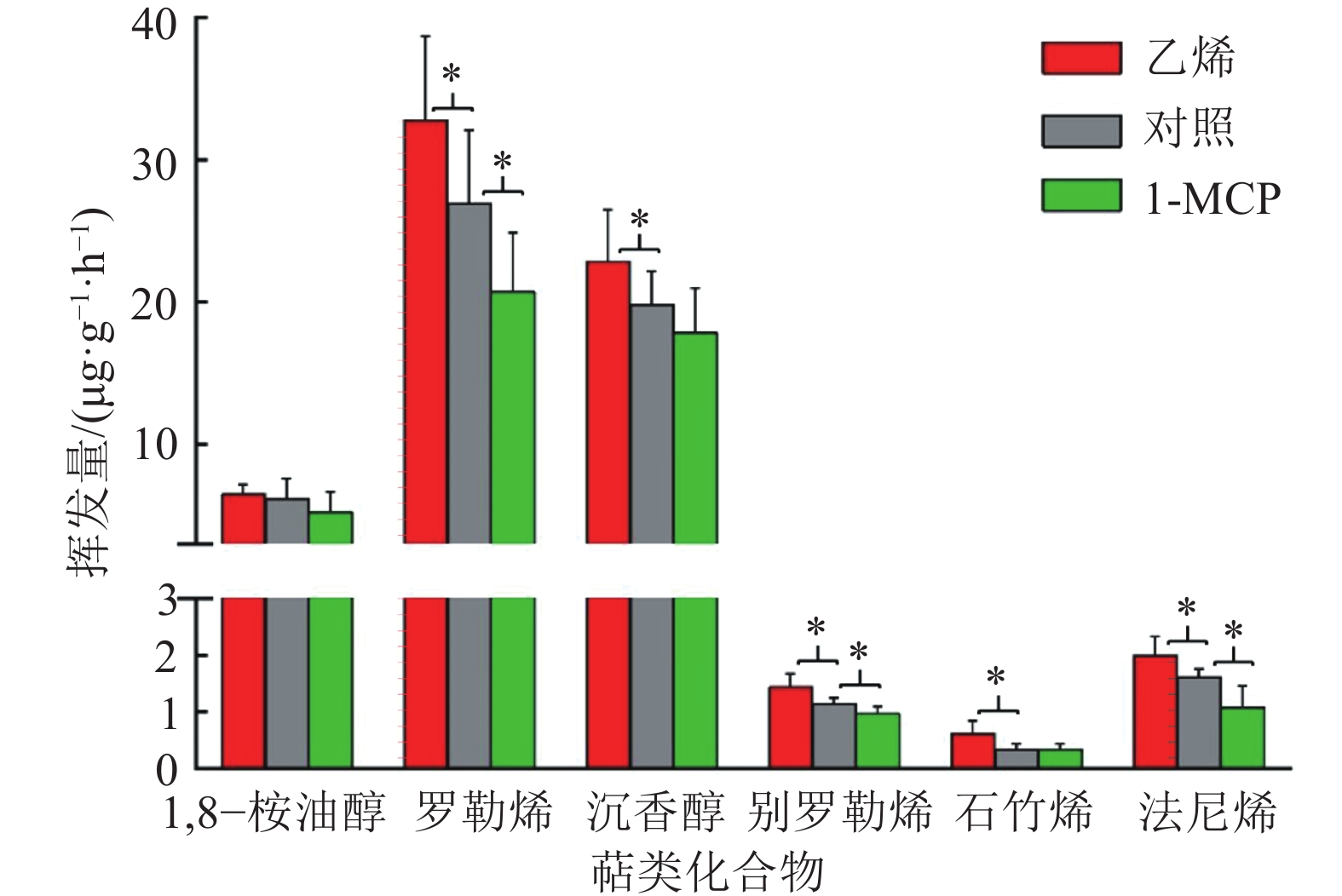
结果乙烯处理后白姜花香气成分罗勒烯、沉香醇、别罗勒烯、石竹烯和法尼烯的挥发量显著上升,相关香气萜类合成酶基因HcTPS1、HcTPS3和HcTPS10的表达量分别上升了116%、182%和63%;而用1-MCP处理后,罗勒烯、别罗勒烯和法尼烯的挥发量显著下降,HcTPS1、HcTPS3和HcTPS10基因的表达量分别降低了45%、30%和30%。乙烯处理后,核转录因子HcEIL1-1和HcEIL1-2的表达量分别上升了51%和114%;1-MCP处理后,HcEIL1-1和HcEIL1-2表达量分别下降了30%和60%。
结论外源乙烯可以提升白姜花萜类花香物质的释放量,白姜花乙烯信号通路中的核转录因子HcEIL1-1和HcEIL1-2与萜类花香合成酶基因的表达模式一致。
Abstract:ObjectiveTo investigate the effect of ethylene and 1-methylcyclopropene (1-MCP) treatment on the terpenoid fragrance of Hedychium coronarium and analyze the expressions of related genes.
MethodH. coronarium was treated separately with 10 μL·L–1 ethylene and 4 μL·L–1 ethylene inhibitor 1-MCP for 8 h. Volatile scent compounds were measured by headspace solid-phase microextraction and gas chromatography-mass spectrometry (GC-MS), and the obtained ion chromatograms were analyzed using NIST 08 database. The expressions of related genes were analyzed by fluorescence quantitative PCR.
ResultAfter ethylene treatment, the emissions of volatile scent compounds ocimene, linalool, allo-ocimene, β-caryophyllene and α-farnesene increased significantly, and the expressions of related terpene synthase genes HcTPS1, HcTPS3 and HcTPS10 increased by 116%, 182% and 63% respectively. After 1-MCP treatment, the emissions of ocimene, allo-ocimene and α-farnesene decreased significantly, and the expressions of HcTPS1, HcTPS3 and HcTPS10 decreased by 45%, 30% and 30% respectively. After ethylene treatment, the expressions of nuclear transcription factors HcEIL1-1 and HcEIL1-2 increased by 1.51 and 2.14 times respectively. After 1-MCP treatment, the expressions of HcEIL1-1 and HcEIL1-2 decreased by 30% and 60% respectively.
ConclusionExogenous ethylene can increase the emission of terpenoids in H. coronarium. The expression patterns of HcEIL1-1 and HcEIL1-2, the nuclear transcription factors in ethylene signaling pathway, were consistent with the expression pattern of terpenoid synthase genes.
-
Keywords:
- Hedychium coronarium /
- fragrance /
- ethylene /
- 1-MCP /
- gene expression /
- terpenoid
-
-
![]()
图 1 乙烯和1-MCP处理对白姜花挥发性萜类香气物质的影响
“*”表示与对照差异达到0.05的显著水平(单因素方差分析)
Figure 1. Effects of ethylene and 1-MCP treatments on volatile terpene components of Hedychium coronarium
![]()
图 2 乙烯和1-MCP处理对白姜花萜类合成酶相关基因表达的影响
“*”和“**”分别表示与对照差异达到0.05和0.01的显著水平(单因素方差分析)
Figure 2. Effects of ethylene and 1-MCP treatments on the expressions of terpene synthase related genes in Hedychium coronarium
![]()
图 3 乙烯和1-MCP处理对HcEIL1-1和HcEIL1-2表达的影响
“*”和“**”分别表示与对照差异达到0.05和0.01的显著水平(单因素方差分析)
Figure 3. Effects of ethylene and 1-MCP treatments on the expressions of HcEIL1-1 and HcEIL1-2
-
[1] 李瑞红, 范燕萍. 白姜花不同开花时期的香味组分及其变化[J]. 植物生理学通讯, 2007, 43(1): 176-180. [2] 岳跃冲, 范燕萍. 植物萜类合成酶及其代谢调控的研究进展[J]. 园艺学报, 2011, 38(2): 379-388. [3] ARC E, SECHET J, CORBINEAU F, et al. ABA crosstalk with ethylene and nitric oxide in seed dormancy and germination[J]. Front Plant Sci, 2013, 4: 63. doi: 10.3389/fpls.2013.00063
[4] WANG Q, ZHANG W, YIN Z, et al. Rice CONSTITUTIVE TRIPLE-RESPONSE2 is involved in the ethylene-receptor signalling and regulation of various aspects of rice growth and development[J]. J Exp Bot, 2013, 64(16): 4863-4875.
[5] IQBAL N, KHAN N A, FERRANTE A, et al. Ethylene role in plant growth, development and senescence: Interaction with other phytohormones[J]. Front Plant Sci, 2017, 8: 475. doi: 10.3389/fpls.2017.00475
[6] BARRY C S, GIOVANNONI J J. Ethylene and fruit ripening[J]. J Plant Growth Regul, 2007, 26(2): 143-159.
[7] NEGRE F, KISH C M, BOATRIGHT J, et al. Regulation of methylbenzoate emission after pollination in snapdragon and petunia flowers[J]. Plant Cell, 2003, 15(12): 2992-3006.
[8] UNDERWOOD B A, TIEMAN D M, SHIBUYA K, et al. Ethylene-regulated floral volatile synthesis in Petunia corollas[J]. Plant Physiol, 2005, 138(1): 255-266.
[9] SEXTON R, STOPFORD A P, MOODIE W T, et al. Aroma production from cut sweet pea flowers (Lathyrusodoratus): The role of ethylene[J]. Physiol Plantarum, 2005, 124(3): 381-389.
[10] 邹晶晶. 乙烯对桂花花色花香成分的影响[C]//张启翔. 中国观赏园艺研究进展2017. 成都: 中国林业出版社, 2017: 458-462. [11] LI G, JIA H, LI J, et al. Effects of 1-MCP on volatile production and transcription of ester biosynthesis related genes under cold storage in ‘Ruanerli’ pear fruit (Pyrusussuriensis Maxim.)[J]. Postharvest Biol Tec, 2016, 111: 168-174.
[12] 王宝春, 颉敏华, 王学喜, 等. 1-MCP处理对冷藏期间花牛苹果香气成分和果实品质的影响[J]. 食品工业科技, 2017, 38(7): 331-339. [13] 马婷, 任亚梅, 张艳宜, 等. 1-MCP处理对‘亚特’猕猴桃果实香气的影响[J]. 食品科学, 2016, 37(2): 276-281. [14] 范燕萍, 余让才, 黄蕴, 等. 姜花挥发性成分的固相微萃取:气相色谱质谱分析[J]. 园艺学报, 2003, 30(4): 475-475. [15] 刘洋. 百合COP1和SPA对光的响应及在花香形成中的作用[D].广州: 华南农业大学, 2016. [16] CHEN H, XUE L, CHINTAMANANI S, et al. ETHYLENE INSENSITIVE3 and ETHYLENE INSENSITIVE3-LIKE1 repress SALICYLIC ACID INDUCTION DEFICIENT2 expression to negatively regulate plant innate immunity in Arabidopsis[J]. Plant Cell, 2009, 21(8): 2527-2540.
[17] SCHADE F, LEGGE R L, THOMPSON J E. Fragrance volatiles of developing and senescing carnation flowers[J]. Phytochemistry, 2001, 56(7): 703-710.
[18] BORDA A M, CLARK D G, HUBER D J, et al. Effects of ethylene on volatile emission and fragrance in cut roses: The relationship between fragrance and vase life[J]. Postharvest Biol Tec, 2011, 59(3): 245-252.
[19] CHANG KN, SHAN Z, WEIRAUCH M T, et al. Temporal transcriptional response to ethylene gas drives growth hormone cross-regulation in Arabidopsis[J]. Elife, 2013, 2(11): 1-20. CHANG KN, SHAN Z, WEIRAUCH M T, et al. Temporal transcriptional response to ethylene gas drives growth hormone cross-regulation in Arabidopsis[J]. Elife, 2013, 2(11): 1-20.
[20] WAWRZYNSKA A, SIRKO A. EIN3 interferes with the sulfur deficiency signaling in Arabidopsis thaliana through direct interaction with the SLIM1 transcription factor[J]. Plant Sci, 2016, 253: 50-57.
[21] MBEGUIE-A-MBEGUIE D, HUBERT O, FILS-LYCAON B, et al. EIN3-like gene expression during fruit ripening of Cavendish banana (Musa acuminata cv. Grande naine)[J]. Physiol Plantarum, 2008, 133(2): 435-448.
[22] LI X, PAN Y, CHANG B, et al. NO promotes seed germination and seedling growth under high salt may depend on EIN3 protein in Arabidopsis[J]. Front Plant Sci, 2016, 6: 1203. doi: 10.3389/fpls.2015.01203
[23] ZHU Z, AN F, FENG Y, et al. Derepression of ethylene-stabilized transcription factors (EIN3/EIL1) mediates jasmonate and ethylene signaling synergy in Arabidopsis[J]. P Natl Acad Sci USA, 2011, 108(30): 12539-12544.
[24] PENG J, LI Z, WEN X, et al. Salt-induced stabilization of EIN3/EIL1 confers salinity tolerance by deterring ROS accumulation in Arabidopsis[J]. PLoS Genet, 2014, 10(10): e1004664.
-
期刊类型引用(3)
1. 孙栋栋,陈丹,徐哲壮,章汉林. 基于US-FMT*算法的机械臂路径规划. 电子测量技术. 2025(04): 16-24 .  百度学术
百度学术
2. 袁雷,贾小林,顾娅军,徐正宇. 融合椭圆约束的快速行进树路径规划算法. 计算机应用研究. 2024(12): 3595-3599 . 百度学术
3. 余晓云,陶明霞,叶剑标. 基于BIM技术的建筑钢筋混凝土框架梁柱构件数字化建模研究. 黑龙江工业学院学报(综合版). 2024(12): 152-156 . 百度学术
其他类型引用(1)
 下载:
下载:
计量
- 文章访问数: 1582
- HTML全文浏览量: 9
- PDF下载量: 1639
- 被引次数: 4
