
Effects of potassium deficiency on growth and N, P and K balance of banana shoots and roots
-
摘要:目的
明确香蕉苗缺钾的表型症状,探讨缺钾和恢复供钾对香蕉苗生长及氮磷钾营养平衡的影响,为香蕉苗营养诊断和平衡施肥提供依据。
方法通过石英砂培养试验,以正常的完全营养液为对照,观察记录香蕉苗缺钾的表型症状及出现时间,对比研究缺钾和缺钾后恢复供钾条件下香蕉苗的地上部和根系的生物量、根系形态参数、氮磷钾吸收等变化。
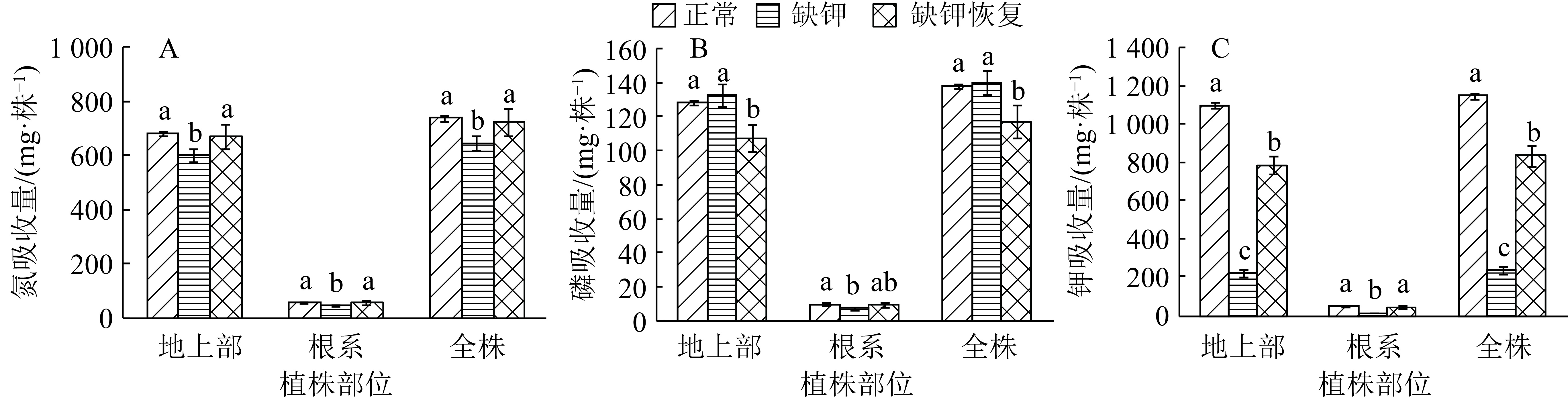
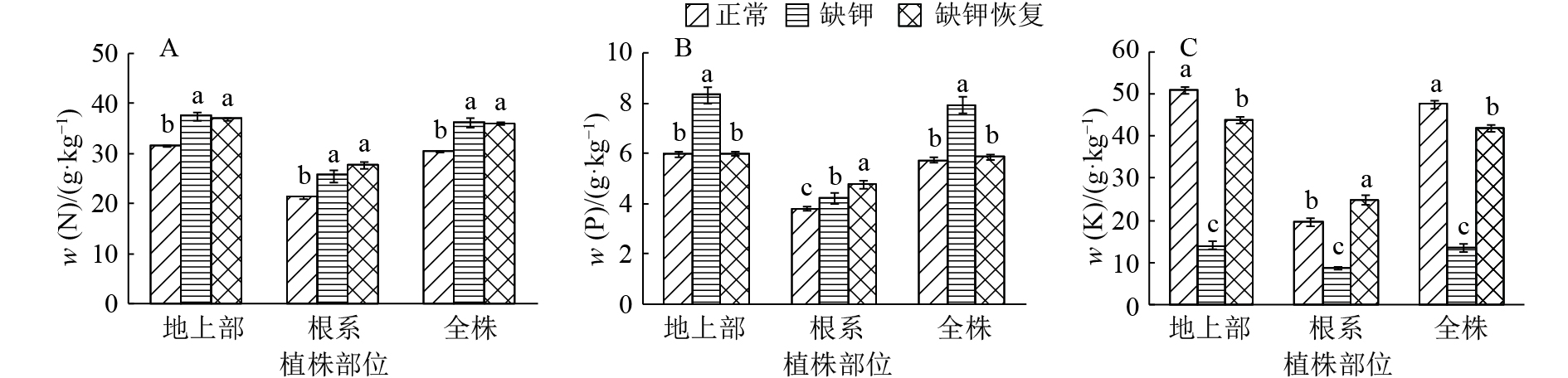
结果缺钾的表型症状为:老叶发黄,中部叶片主叶脉先呈现橙黄色,并逐渐向叶缘蔓延,然后部分叶片叶缘出现近椭圆形小枯斑,叶柄呈现浅红褐色,缺钾严重时,橙黄色叶片近叶脉处的叶肉发黑,进而干枯死亡,枯叶呈褐色,完全展开的新叶不挺立,上部叶片柔弱、下披。缺钾显著抑制香蕉苗生长,缺钾处理较正常处理的地上部和根系生物量降低了31%和39%,其根长、根表面积和根体积减少了30%、31%和34%,且细根减幅大于中根。缺钾同时改变了香蕉苗氮磷钾平衡,其地上部、根系和全株的氮含量分别增加了19%、21%和19%,磷含量分别增加了40%、12%和38%,钾含量分别降低了73%、56%和72%,氮、钾吸收量降低,根系的磷吸收量降低;缺钾处理的地上部、根系和全株的N/K、P/K显著升高,地上部和全株N/P显著降低,根系N/P显著升高。缺钾后恢复供钾,香蕉苗缺钾表型症状消失,但香蕉苗的生物量以及氮、磷、钾含量和吸收量不能同步恢复,香蕉苗生物量仍低于正常处理,各部位的氮含量、根系的磷、钾含量仍高于正常处理,地上部和全株的钾含量和吸收量也未增加至正常水平。供钾后地上部和全株的磷吸收量显著降低,且低于正常处理。
结论缺钾对香蕉苗营养状况的影响显著大于对表型性状的影响,营养状况的恢复滞后于表型性状的恢复,香蕉苗施肥要针对其营养特性施用。
Abstract:ObjectiveTo make clear morphological symptoms of potassium (K) deficiency in banana seedlings, discuss the effects of K deficiency and K resupply on growth and nitrogen (N), phosphorous (P), K balance of banana seedlings and provide a basis for nutrition diagnosis and blanced fertilization of banana seedlings.
MethodThrough quartz sand culture experiment, complete nutrient solution cultivation was marked as the control treatment to observe morphological characteristics and appearance time of K-deficient symptoms. K deficiency and K deficiency-resupply treatments were conducted to study the changes of biomass, root morphological parameters and N, P, K uptake of banana seedlings.
ResultThe K-deficient symptoms were old leaves chlorosis, successive orange-yellowing of main veins and margins of middle leaves, small oval spots in leaf margins and reddish brown petioles. When the seedlings were in severe K deficiency, mesophylls near the veins of orange-yellow leaves were blackening and dried to death, the dead leaves were brown, fully expanding new leaves were not erect, and the upper leaves were soft and drooping. K deficiency significantly inhibited the growth of banana seedlings. Compared with CK, the biomass of shoots and roots in K deficiency treatment decreased by 31% and 39% respectively, the length, surface area and volume of roots decreased by 30%, 31% and 34% respectively, and the decrease ranges of fine roots were more than that of middle roots. K deficiency also changed N, P, K balance of banana seedlings. Compared with CK, N contents of shoots, roots and whole plants in K-deficient treatment increased by 19%, 21% and 19% respectively, P contents increased by 40%, 12% and 38% respectively, K contents decreased by 73%, 56% and 72% respectively, N and K uptake decreased, P uptake of roots decreased, N/K and P/K ratios in shoots, roots and whole plants increased significantly, N/P ratio in shoots and whole plants decreased significantly, and N/P ratio in roots increased significantly. After resupplying K in K deficiency treatment, the K-deficient morphological symptoms of banana seedlings disappeared, but the biomass, N, P and K contents and uptakes of banana seedlings could not recover simultaneously. The biomass of banana seedlings was still lower than that of CK, and N contents of shoots, roots and whole plants and P, K contents of roots were higher than those of CK. K contents and uptakes of shoots and whole plants did not increase to normal levels. P uptakes of shoots and whole plants decreased significantly and was lower than that of CK.
ConclusionThe effects of K deficiency on nutrition status of banana seedlings are significantly greater than that of phenotypic characteristics, and the recovery of nutrition status lags behind phenotypic characters. Banana seedling fertilization should apply to its nutritional characteristics.
-
Keywords:
- potassium deficiency /
- banana seedling /
- shoot /
- root /
- N, P and K balance
-
种群是物种存在、进化及种间关系的基本单位,是连接群落和个体的纽带,也是生态系统的基本组成部分[1]。扩散是几乎所有生物生活史的一个基本特征,生物的扩散过程大大影响生物组织的形成,如:个体的存活、生长和繁殖,种群和群落的组成、结构和动态,物种的维持、进化和地理分布等[2-3]。目前,种群扩散研究日益受到广泛关注,已成为生态学研究中的一个热点领域[4-7]。在自然界中,大部分植物通过花粉和种子扩散来完成种群的扩张和迁移,花粉和种子扩散是关乎植物群落动态的一个重要生态过程[8],它与生境、种源特征和传播者有着密切的关系,因植物生活型和所处生境的不同而存在明显的差别[9-10];同时,它能够直接影响幼苗的命运,进而影响种群的更新动态和分布格局[11]。因此,种群扩散机制的研究,对种群的维持、物种的进化、物种的成种和预测其种群演化趋势等方面具有重要意义。
黄牛木Cratoxylum cochinchinense是亚热带地区的乡土树种[12],其抗逆性好、适生性强,在酸性、砂质等贫瘠土壤中生长良好,是我国华南地区的优良先锋树种[13-14]。黄牛木还是极具潜力的城市景观树种,其黄褐色树干形成的特色林内景观以及春夏花期形成的彩色景观具有较高观赏价值[15]。目前,对黄牛木的研究主要集中于成分分析及药用价值方面[16-17],其种群生态学的研究较少,鲜见有种群扩散方面的研究报道。纵观我国华南珠三角等发达地区,尽管城市内的绿地景观已发展得较为成熟,但城市周边仍有一些生态脆弱的山林地,这些困难立地林相不稳定,景观效果有待提升。黄牛木具备优良的品种特性,可作为该地区植被修复和生态改造工程的优选树种。因此,黄牛木种群扩散动态的研究,不仅可为其种群的天然更新和保护管理提供理论依据,为华南地区乡土树种的种群扩散研究提供参考,同时也可促进这一优良乡土树种的开发和利用,从而为森林群落的生态修复及生态系统的可持续发展作出贡献。
1. 材料与方法
1.1 研究地概况
茶山公园(简称茶山)位于广州天河区的五山街(原五山镇,茶山是五山中的其中一山),北与沙河镇长湴村(现天河区长兴街)相邻,南至广九铁路。地理位置为113°21′~113°22′E和23°9′~23°8′N,低丘地形,海拔93.6 m。本文调研样地选自鸡笼顶,样地土壤为花岗岩、变质石英砂岩、流纹岩和红色砾岩发育的赤红壤。
白云山风景名胜区(简称白云山)座落于广州市东北部,位于北纬23°09′~23°13′,东经113°16′~113°19′,为九连山脉向东南延伸的支脉,属丘陵地形,素有“南粤第一山”之称。白云山景区面积28 km2,最高峰为摩星岭,海拔382 m,属南亚热带季风海洋性气候,年均气温21.4~21.8 ℃,年降水量1 689.3~1 876.5 mm,雨季集中在4—9月份,且降水量占全年降水量的85%以上,年均湿度79%。本文调研样地选自竹园景区,样地土壤为流纹状花岗岩、砂页岩、石英岩及混合岩母质发育而成的酸性赤红壤[18]。
石花山森林公园(简称石花山)位于广东省台山市的东北部,地理位置为22°26′~22°81′N和112°81′~112°9′E,石花山虽高300余m,却融奇、秀、幽于一山。台山属亚热带海洋性季风气候,年平均气温21.8 ℃,年平均日照2 006 h,年均降雨量1 936 mm。公园内随处可见未被人工化的自然群落,本文调研样地选自西严寺东面山顶,样地土壤类型为花岗岩发育的酸性赤红壤。
1.2 研究方法
1.2.1 外业调查
在林分踏查的基础上,采用典型取样法,选择具有代表性的黄牛木天然群落地段作为样地布设区域。于2015年12月至2016年2月期间,在广州茶山公园的鸡笼顶(东南坡)、广州白云山风景名胜区的竹园景区(西北坡)分别选取3 900 m2的样地,并在台山石花山森林公园的西严寺东面选取3 200 m2的样地(其中东南坡2 000 m2,西北坡1 200 m2),总面积11 000 m2。对样地内所有高度≥1.3 m的黄牛木个体的胸径、树高、冠幅、枝下高等生长因子进行测定,高度<1.3 m的黄牛木个体测定除胸径之外的其他生长因子,并记录黄牛木个体之间的相对坐标。
1.2.2 种群径阶划分
参照有关种群大小级结构代替年龄结构的分析方法[19],以“空间替代时间”,将林木的胸径大小作为度量其年龄大小的指标。把树高<1.3 m和胸径<1 cm的黄牛木幼苗记为1径阶;树高≥1.3 m且胸径≥1 cm时,每隔2 cm增加一个径阶,将黄牛木依次划分为若干径阶。
1.2.3 种子传播与种群扩散
运用ArcGIS分别绘制样地黄牛木种群个体和冠幅空间分布专题图,分析黄牛木个体的聚集情况和种子的传播方向及种群的扩散方式,然后采用缓冲区分析与空间计算功能测定种群各径阶的累计冠层投影面积,分析其累计面积的增长规律,结合SPSS 17.0软件进行回归模拟分析,构建种群扩散速率相关模型,进而预测黄牛木种群扩散的动态规律。
1.2.4 扩散速率
借鉴胡梦宇[20]对藜蒴Castanopsis fissa种群扩散速率的研究方法,通过计算种群达到某一冠幅时投影增长量所需要的时间,测算黄牛木种群的扩散速率,即单位时间内种群冠层投影的增长面积。本文以胸径代替时间变化,种群扩散速率=种群冠层增长面积/胸径。
2. 结果与分析
2.1 种群扩散方式分析
绘制各样地黄牛木种群扩散分布图(图1)。从图1可以看出,黄牛木个体总是在母树某一方向上呈现明显的聚集分布并逐渐向外扩散,但这一方向不全为下坡方向,白云山的黄牛木种群甚至沿斜上坡方向扩散。可见,黄牛木扩散不只是靠种子重力传播。从图1还可以看出,3个样地的黄牛木种群均表现出不同程度的聚集分布,而且在常年风向的下风向的聚集现象更为明显,说明黄牛木种子的传播受风向影响的可能性较大。风传播是常见的非生物传播方式,许多植物的果实和种子以此方式传播[21],它们一般具有下述特征:1)细小质轻,能悬浮在空气中被风力吹送至远处,如兰科植物稗Echinochloa crusgalli、马唐Digitaria sanguinalis等;2)果实或种子的表面常生有絮毛、果翅,或其他有助于承受风力飞翔的特殊构造, 如马兜铃Aristolochia debilis、云杉属Picea、滇油杉Keteleeria evelyniana等。黄牛木种子为蒴果,一侧具翅,成熟期为夏季,成熟后小而轻[22]。初步推断,风力是黄牛木种群扩散传播的主要动力。
![]() 图 1 黄牛木种群扩散分布情况图中每一景观栅格为10 m×10 m,着色区域代表黄牛木个体的空间分布情况,红色区域为其种群斑块内的胸径最大个体,即种群母树Figure 1. Diffusion and distribution of Cratoxylum cochinchinense populationEach lattice is 10 m × 10 m in the graph,the coloring part indicates spatial distribution of Cratoxylum cochinchinense, and the red part indicates the parent tree with the largest DBH in population
图 1 黄牛木种群扩散分布情况图中每一景观栅格为10 m×10 m,着色区域代表黄牛木个体的空间分布情况,红色区域为其种群斑块内的胸径最大个体,即种群母树Figure 1. Diffusion and distribution of Cratoxylum cochinchinense populationEach lattice is 10 m × 10 m in the graph,the coloring part indicates spatial distribution of Cratoxylum cochinchinense, and the red part indicates the parent tree with the largest DBH in population文献显示,广州的风向常年为偏北或东北方向,夏季盛行风向为东风和东南风[23]。从图1a可以看出,在夏季盛行风向(偏东和东南方向)和常年风向(偏北或东北方向)上,茶山样地的植株呈现明显的聚集现象和扩散趋势,在偏东方向上尤为明显,因此认为种子的扩散受风力的影响极大。当然,由于该样地夏季盛行风向也是该坡的斜下坡方向,故不能排除种子由于受重力作用沿斜坡滚落散布在母树周边区域,使得幼苗在母株周围呈现密集的簇状分布。但从图1b可以看出,白云山种群个体扩散方向为该坡的斜上坡方向,正是该地种子成熟期盛行风向(东南风)的下风向。因此可基本断定,黄牛木种群靠风力扩散。
台山的风向常年为东北风、偏北风,夏季盛行南风[24]。从图1c可以看出,石花山样地的植株在南坡沿正南方向扩散,即种子成熟期盛行风向;在北坡沿东北偏北方向扩散,为该地常年风向。故进一步确定,黄牛木种群靠风力扩散。
2.2 种群扩散动态分析
上述分析表明,种群在不同的地域均呈现出聚集分布,这种分布特性直接关系到林木冠层空间分布的形成。为更直观地呈现黄牛木种群在不同发育阶段的扩散动态,结合样地中黄牛木个体的相对位置和各径阶平均冠幅,运用地理信息系统(GIS)数字化和输出专题图功能绘制黄牛木种群的冠层扩散分布图(图2)。
![]() 图 2 黄牛木种群冠层扩散情况图中每一景观栅格为10 m×10 m,着色区域代表黄牛木冠层投影的空间分布Figure 2. Canopy diffusion of Cratoxylum cochinchinense populationEach lattice is 10 m × 10 m in the graph,the coloring part indicates projection of Cratoxylum cochinchinense canopy
图 2 黄牛木种群冠层扩散情况图中每一景观栅格为10 m×10 m,着色区域代表黄牛木冠层投影的空间分布Figure 2. Canopy diffusion of Cratoxylum cochinchinense populationEach lattice is 10 m × 10 m in the graph,the coloring part indicates projection of Cratoxylum cochinchinense canopy结合茶山样地的黄牛木种群冠层扩散分布图(图2a),运用GIS软件中的合并计算功能,将各径阶分布依次合并,通过图层属性表的几何计算功能得到茶山黄牛木各个径阶冠层投影面积,茶山黄牛木种群第1~11径阶冠层投影增长面积依次为69.245、16.871、182.252、262.519、232.929、258.751、136.663、102.866、64.192、125.410和49.288 m2(表1)。由此可见,黄牛木种群冠层投影面积的增长量随径阶的增加而先增大后减小,说明种群扩散程度随径阶增大先上升后下降,在第4径阶时冠层面积增长量达到最大值262.519 m2,此时种群的扩散程度最高。
表 1 黄牛木种群生长因子统计与扩散速率测定结果Table 1. Growth factor statistics and diffusion rate of Cratoxylum cochinchinense population调查地点
Sample
plot径阶1)
Diameter
class平均胸径/cm
Average
DBH平均冠幅/m
Average crown width投影面积/m2
Canopy projection area累计投影面积/m2
Cumulative projection area投影增长面积/m2
Growth of projected area扩散速率2)/
(m2·cm-1)
Diffusion rate茶山
Chashan1 0.7 0.64 57.970 1 500.986 69.245 54.73 2 2.0 1.54 278.237 1 431.741 16.871 9.24 3 3.8 2.25 452.280 1 414.870 182.252 90.15 4 5.8 3.15 506.477 1 232.618 262.519 132.91 5 7.8 3.54 386.610 970.099 232.929 128.33 6 9.6 4.23 317.200 737.170 258.751 122.28 7 11.8 4.36 161.358 478.419 136.663 82.22 8 13.4 5.99 146.789 341.756 102.866 42.27 9 15.9 6.38 64.192 238.890 64.192 35.66 10 17.7 6.58 125.410 174.698 125.410 67.79 11 19.5 7.90 49.288 49.288 49.288 − 白云山
Baiyun mountain1 0.8 0.79 74.644 1 374.181 17.406 15.27 2 1.9 1.24 301.900 1 356.775 102.391 54.52 3 3.8 1.94 458.132 1 254.384 232.904 112.39 4 5.9 2.61 517.476 1 021.480 366.714 189.88 5 7.8 3.28 340.751 654.766 262.731 137.16 6 9.7 3.84 189.505 392.035 170.537 98.44 7 11.5 3.99 148.960 221.498 136.265 49.55 8 14.2 4.55 16.249 85.233 16.249 8.78 9 16.1 4.69 68.984 68.984 68.984 − 石花山
Shihua mountain1 0.8 0.38 0.775 709.426 0.115 0.09 2 2.1 1.11 16.533 709.311 6.207 3.14 3 4.1 2.10 91.076 703.104 42.808 24.96 4 5.8 2.66 177.765 660.296 100.895 45.04 5 8.0 3.49 244.906 559.401 152.533 91.28 6 9.7 4.23 220.326 406.868 186.753 79.85 7 12.1 5.25 194.700 220.115 194.700 76.35 8 14.6 1.45 1.673 25.415 1.672 1.11 9 16.1 5.50 23.743 23.743 23.743 − 1)第1径阶为树高 <1.3 m 和胸径 <1 cm 的黄牛木个体,其余径阶为树高 ≥1.3 m 且胸径≥1 cm 的黄牛木个体,胸径每隔 2 cm 增加一个径阶;2) “−” 代表需测得下一径阶数据方可进行计算
1) The first diameter class includes Cratoxylum cochinchinense trees with height <1.3 m and DBH <1 cm, the other diameter classes include trees with height≥1.3 m and DBH≥1 cm, and the diameter class increases one class with every 2 cm increase in DBH; 2) “−” indicates the value can not be calculated without data of the next diameter class结合白云山样地的黄牛木种群冠层扩散分布图(图2b),计算得到白云山黄牛木种群第1~9径阶冠层投影增长面积依次为17.406、102.39、232.904、366.714、262.731、170.537、136.265、16.249和68.984 m2(表1)。可见,黄牛木种群的冠层投影面积的增长量随径阶的增加而先增大后减小,和茶山的冠层投影面积增长趋势相似,在第4径阶种群扩散程度最高。
结合石花山样地的黄牛木种群冠层扩散分布图(图2c),计算得到黄牛木种群从第1~9径阶冠层投影增长面积依次为0.115、6.207、42.808、100.895、152.533、186.753、194.700、1.672和23.743 m2(表1)。可见,该地黄牛木种群的冠层扩散趋势与另外2个样地相似,冠层投影面积的增长量随径阶的增加而先增大后减小,但种群的扩散峰值有所延后,在第7径阶达到最大值,种群扩散程度最高。
2.3 种群扩散速率分析
黄牛木种群始终处于动态变化中,不同时期的种群斑块有不同的形状及大小。经统计相关数据,结合GIS空间分析与几何计算功能测定黄牛木种群冠层投影面积,进而计算种群的扩散速率,测定结果见表1。
运用SPSS 17.0软件进行曲线回归分析,得到黄牛木种群扩散速率与平均胸径的函数拟合曲线模型,茶山的相关参数估计值见表2(其他省略)。R2值越大、P值越小,两者的相关性越显著,据此选取黄牛木种群扩散速率的最适函数模型:茶山y1=−1.018 1x12+ 18.071x1+ 27.365(R2=0.443,P=0.129);白云山y2=−3.086 2x22+ 43.308x2 − 8.6201( R2=0.821,P=0.000);石花山y3=−1.5698x32 + 26.826x3 −39.059( R2=0.790,P=0.048)。其中x是各个径阶对应的平均胸径,cm;y是各个径阶对应的扩散速率,m2/cm。
表 2 茶山样地黄牛木种群扩散速率与平均胸径的函数模型方程及参数估计Table 2. Function model and parameter estimation of diffusion rate - average DBH for Cratoxylum cochinchinense population in Chashan sample plot方程
Equation参数估计值 Estimated parameter R2 F ν1 ν2 P a b1 b2 线性 Linear 0.003 0.023 1 8 0.882 80.041 −0.393 对数 Logarithm 0.037 0.310 1 8 0.593 61.672 8.042 倒数 Reciprocal 0.082 0.717 1 8 0.422 85.001 −30.106 二次 Quadratic 0.443 2.786 2 7 0.129 27.365 18.071 −1.018 复合 Composite 0.018 0.145 1 8 0.713 52.329 1.019 幂 Power 0.082 0.719 1 8 0.421 40.396 0.229 S 0.065 0.560 1 8 0.476 4.266 −0.514 增长 Growth 0.018 0.145 1 8 0.713 3.958 0.019 指数 Index 0.018 0.145 1 8 0.713 52.329 0.019 Logistic 0.018 0.145 1 8 0.713 0.019 0.982 模型构建结果表明,各样地种群扩散速率的函数模型均为一元二次方程。可以计算出,当种群的扩散速率无限接近0时,黄牛木种群冠幅的扩散受到阻碍,此时必须对黄牛木进行间伐,以促进种群的生长与更新。将y=0分别代入各样地的函数模型,得到x1=19.2 cm,x2=13.8 cm,x3=15.5 cm;即黄牛木的茶山种群处于第10径阶,白云山种群处于第8径阶,石花山种群处于第9径阶时,应对黄牛木种群进行间伐。因此,在目前的生长状态下,3个样地均应采取适当的间伐措施,以促进黄牛木天然林的更新生长和保护管理。
从模型构建结果还可以看出,各样地黄牛木种群扩散趋势相似,可以整理获得黄牛木种群扩散的一般规律。计算黄牛木种群胸径和扩散速率的平均值,并运用SPSS软件进行曲线拟合回归分析,得到黄牛木种群的扩散速率模型:y =−0.969 2x2 + 17.336x + 20.531(R2=0.943,P=0.020)。
2.4 单株黄牛木扩散速率分析
整理外业调查数据,得到单株黄牛木各径阶的胸径、冠幅、冠幅面积以及冠幅增长面积,结果见表3。通过对冠幅与胸径进行回归分析,得到二次函数的R2最大,为0.998,P为0.000,说明冠幅(y)与胸径(x)呈显著正相关,故单株黄牛木的冠幅与胸径的最佳拟合模型是一元二次方程,函数公式为y4=−0.009 6x42 + 0.481 9x4 + 0.303 9(R2=0.999,P=0.000)。当黄牛木的胸径为3.03 cm时,黄牛木的模拟冠幅为1.18 m,与实际调查数据1.67 m相差不大,拟合情况较好。
表 3 黄牛木单株生长因子调查与模拟结果Table 3. Survey and simulation results of growth factors of individual Cratoxylum cochinchinense tree径阶1)
Diameter
class数量/株
Number胸径/cm
DBH实际调查结果 Survey result 模拟分析结果 Simulation analysis result 冠幅/m
crown width冠幅面积/m2
crown area增长面积/m2
growth area冠幅/m
crown width冠幅面积/m2
crown area增长面积/m2
growth area扩散速率/
(m2·cm−1) Diffusion rate1 422 0.8 0.60 0.286 0.286 0.68 0.366 0.366 0.4 2 607 2.0 1.29 1.316 1.030 1.24 1.209 0.843 0.7 3 389 3.9 2.10 3.451 2.135 2.02 3.209 2.000 1.1 4 250 5.8 2.81 6.189 2.738 2.79 6.114 2.905 1.5 5 123 7.9 3.43 9.259 3.070 3.51 9.681 3.567 1.7 6 59 9.7 4.10 13.190 3.930 4.08 13.060 3.379 1.9 7 34 11.7 4.53 15.658 2.468 4.63 16.828 3.769 1.9 8 8 14.1 5.27 21.809 6.151 5.19 21.126 4.298 1.8 9 7 16.0 5.52 23.926 2.118 5.56 24.238 3.112 1.6 1) 第 1 径阶为树高 <1.3 m 和胸径 <1 cm 的黄牛木个体,树高≥1.3 m 且胸径≥1 cm 的黄牛木个体,其胸径每隔 2 cm 增加一个径阶
1) The first diameter class includes Cratoxylum cochinchinense trees with height <1.3 m and DBH <1 cm, the other diameter classes include trees with height≥1.3 m and DBH≥1 cm, and the diameter class increases one class with every 2 cm increase in DBH由冠幅−胸径函数得到单株黄牛木冠幅模拟数据,计算出冠幅扩散速率的数值,模拟结果见表3。运用扩散速率与胸径作回归分析,其最佳拟合模型亦是一元二次方程,函数公式为:y0=−0.013 5x02 + 0.310 6x0 + 0.111 3(R2=0.999,P=0.000)。
从表3中数据可以看出,单株黄牛木的扩散速率随径阶增大先增大后减小,说明黄牛木在生长发育的初期,营养、空间等资源较充足,种群扩散比较顺利;随着黄牛木年龄的增长,种群扩散速率趋势由上升趋势变为下降,直至无限接近于0,其冠幅扩散速率同样无限接近于0,此时黄牛木冠幅达到最大值,若不采取适当措施,黄牛木的冠幅增长将受到限制。
3. 讨论与结论
本文结合种群生态学和景观生态学的研究方法,通过GIS技术,对黄牛木种群的扩散规律进行系列分析,从种子的传播方式到种群的扩散动态,然后建立种群扩散速率的函数模型,进而模拟预测种群未来的发展趋势。本研究为植物种群扩散和空间分布研究提供了一种新的思路和方法。
植物种群扩散通常与种子扩散关系密切,而种子扩散往往需要借助动物、风、水、机械及人等媒介。在热带雨林中,满身是刺的黑尾狐棕靠刺鼠偷食其种子得以扩散传播[25];胡梦宇[20]则通过对不同样地藜蒴种群的景观物种流研究,确定了藜蒴种子靠重力传播。黄牛木种子质轻具翅,其种子扩散方向多为当地夏季(黄牛木种子成熟期)盛行风向或常年风向的下风向,这与风传播的特征吻合[21, 26],因此认为黄牛木种群靠风力扩散。但是,不排除其他方式传播的可能性,例如,在种子和根茎的扩散过程中遇到障碍使大量繁殖体集中在一处而其他地方很少或完全没有,动物及人为活动对聚集分布也有加强作用。今后可以采用标记黄牛木种子的方法来精确、直接地研究种子的传播,进一步确定种子在不同途径下的传播机制,丰富黄牛木的生物学理论,为种群空间分布动态和扩散规律提供更深厚的研究基础。
黄牛木的冠层扩散情况表明,冠层扩散程度随径阶增大先上升后下降,原因是群落环境水分、光照、养分等自然条件从充足到紧缺、种内和种间的竞争加大,最终导致扩散程度的变化,这种变化为物种生长扩散过程中的正常现象,该结果支持了前人的研究[20, 27-28]。但石花山的扩散程度峰值有所延后,原因可能是在生长发育初期,种群基数小,产生的种子也较少,以至于扩散程度较低,随着种群数量的增加,种群基数也逐年增加,扩散程度亦逐渐上升;也有可能是种群在扩散过程中受到环境、人为等方面的影响,具体原因有待进一步研究。研究黄牛木种群的冠层变化规律,有利于深入了解植物上层空间随时间变化的特点,为种群空间分布研究提供理论基础。
运用冠层面积增长动态推算种群扩散规律,结果表明,种群扩散速率随种群年龄增加先增大后减小,各样地种群扩散速率与胸径的函数模型为一元二次方程,这与胡梦宇[20]对珠三角地区不同样地藜蒴景观林的研究结果基本一致,说明不同种群的扩散趋势有相似之处。但各个模拟函数的系数均不相同,说明不同的生长环境其生长情况可能不同,模拟方程呈现的是特定区域种群生长扩散的规律,不能盲目套用,一概而论。赵强民[28]通过对广州白云山木荷种群的扩散速率进行研究,得到木荷种群的冠层累计投影面积与径阶的函数模型为一元一次方程,说明种群扩散速率的模型构建有不同的拟合选择,应充分根据不同研究对象进行合理选择。未来,还要进一步考虑边缘效应、林窗效应、自然稀疏的随机性以及种子传播过程中风速产生的影响,提升黄牛木种群扩散速率模拟预测的精确度,为种群的合理间伐及抚育更新提供理论依据。
通过对单株黄牛木的胸径增长和冠幅扩散规律进行研究,利用扩散速率与胸径的函数模型,可进一步确定人工经营下黄牛木种植密度以及间伐时间,由扩散速率-胸径函数模型计算得到,当y达到最大值1.90 m2/cm时,x =11.5 cm,即黄牛木的胸径为11.5 cm时,种群处于第6径阶,其冠幅扩散速率达到最大值1.90 m2/cm,此时应对种群进行适当间伐,以保证其最大效益;将y=0代入函数,计算得到x=23.4,即黄牛木胸径达到23.4 cm时,种群处于第12径阶,此时必须对黄牛木进行间伐,否则将会阻碍种群的生长。当然,树种的生长状况处于不断变化中,以上推论为理想状态下黄牛木的间伐时间,但可以肯定的是,适时进行合理的人工疏伐,有利于实现人工经营下黄牛木种群的最大景观价值和森林生产效益。
黄牛木作为华南地区的优良乡土树种,目前仍处于天然群落生长状态,对黄牛木扩散机制的研究还不够深入,对其种子传播后种子的命运、种子适应各种传播途径的机理以及扩散速率与风速之间的关系等方面的研究都有待加强。因此,只有了解了黄牛木的扩散机制,才能采取有针对性的措施,实现黄牛木天然林的合理抚育和有效管理。从人工经营管理的角度而言,分析黄牛木的生长规律和扩散动态,可加快黄牛木在景观绿化及植被修复中的推广应用,从而促进困难立地的生态平衡与发展。
-
![]()
图 1 缺钾对香蕉苗氮、磷、钾含量的影响
各图中同一植株部位的柱子上方凡是具有一个相同小写字母者,表示处理间差异不显著(P>0.05,Duncan’s 法)
Figure 1. Effects of K deficiency on N, P and K contents in banana seedlings
![]()
图 2 缺钾对香蕉苗氮、磷、钾吸收量的影响
各图中同一植株部位的柱子上方凡是具有一个相同小写字母者,表示处理间差异不显著(P>0.05,Duncan’s 法)
Figure 2. Effects of K deficiency on N, P and K uptakes in banana seedlings
表 1 改良的Hoagland营养液配方1)
Table 1 Chemical composition of improved Hoagland nutrient solution c/(μmol·L–1)
化合物 正常 缺钾 KNO3 6 000 Ca(NO3)2·4H2O 4 000 4 000 NH4H2PO4 2 000 2 000 MgSO4·7H2O 1 000 1 000 NH4NO3 3 000 KCl 50 NaCl 50 (NH4)6Mo7O24·4H2O 0.071 0.071 EDTA-Na·Fe 50 50 MnSO4·H2O 2 2 ZnSO4·7H2O 2 2 CuSO4·5H2O 0.5 0.5 H3BO3 25 25 1)表中为1个剂量浓度的营养液配方  下载: 导出CSV
下载: 导出CSV
表 2 香蕉苗缺钾典型症状及时间表
Table 2 Typical characteristics and appearance time of K-deficient symptoms in banana seedlings
t缺钾/d 缺钾症状 40 老叶开始发黄;中部叶片橙黄化,从主脉开始向叶缘蔓延,叶缘现一行密集的椭圆形小枯斑 50 抽叶速度变慢,株高生长缓慢 60 叶片近叶脉处的叶肉发黑;上部叶片疲软 80 叶片死亡速度加快,枯叶数急剧增加 200 植株只有3片绿叶,生长几乎停滞 260 植株接近死亡
下载: 导出CSV
表 3 缺钾对香蕉苗每株干生物量的影响1)
Table 3 Effects of K deficiency on banana seedling dry biomass
m/g 处理 地上部 根 全株 正常 22.52±0.21a 2.92±0.12a 25.44±0.25a 缺钾 15.50±0.86c 1.79±0.22b 17.22±1.02c 缺钾恢复 17.98±0.75b 1.89±0.15b 19.88±0.89b 1)同列数据后凡是具有一个相同小写字母者表示处理间差异不显著(P>0.05,Duncan’s 法)
下载: 导出CSV
表 4 缺钾对香蕉苗根系形态参数的影响1)
Table 4 Effects of K deficiency on morphological parameters of banana seedling roots
处理 每株根长/cm 每株根表面积/cm2 平均根直径/mm 每株根体积/cm3 正常 15 675.2±1 391.9a 2 507.9±202.1a 0.510±0.011a 32.88±2.43a 缺钾 11 034.3±492.8b 1 724.1±62.5b 0.498±0.004a 21.70±0.62c 缺钾恢复 13 285.6±1 190.9ab 2 110.8±123.4ab 0.509±0.021a 27.45±1.13b 1)同列数据后凡是具有一个相同小写字母者表示处理间差异不显著(P>0.05,Duncan’s 法)
下载: 导出CSV
表 5 缺钾对香蕉苗不同直径根系形态参数的影响1)
Table 5 Effects of K deficiency on morphological parameters of banana seedling roots with different diameter
处理 每株根长/cm 每株根表面积/cm2 每株根体积/cm3 细根 中根 粗根 细根 中根 粗根 细根 中根 粗根 正常 15 212.1±1 361.9a 371.5±25.5a 72.3±3.3a 1 721.8±155.1a 346.5±22.7a 128.6±5.2a 28.6±3.0a 27.2±1.7a 18.9±0.6a 缺钾 10 709.8±482.8b 305.8±11.2b 8.2±0.4c 1 256.6±42.8b 266.5±11.5b 13.6±0.6c 19.8±0.5c 19.2±1.0b 1.8±0.1c 缺钾恢复 12 875.0±1 210.2a 366.7±25.1a 28.6±4.8b 1 480.6±110.4b 343.4±23.3a 48.8±9.4b 23.2±1.5b 26.8±1.7a 6.8±1.5b 1)同列数据后凡是具有一个相同小写字母者表示处理间差异不显著(P>0.05,Duncan’s 法)
下载: 导出CSV
表 6 缺钾对香蕉苗氮、磷、钾平衡的影响
Table 6 Effects of K deficiency on N, P and K balance in banana seedlings
植株部位 处理 N/K N/P P/K N∶P∶K 地上部 正常 0.62±0.01 5.32±0.10 0.12±0.00 1.00∶0.19∶1.61 缺钾 2.66±0.24 4.42±0.13 0.60±0.04 1.00∶0.23∶0.38 缺钾恢复 0.85±0.01 6.22±0.12 0.14±0.00 1.00∶0.16∶1.18 根系 正常 1.10±0.05 5.67±0.18 0.19±0.01 1.00∶0.18∶0.92 缺钾 3.08±0.17 6.10±0.10 0.50±0.03 1.00∶0.16∶0.33 缺钾恢复 1.13±0.03 5.85±0.26 0.19±0.01 1.00∶0.17∶0.89 全株 正常 0.64±0.01 5.34±0.08 0.12±0.00 1.00∶0.19∶1.55 缺钾 2.68±0.23 4.51±0.11 0.59±0.04 1.00∶0.22∶0.37 缺钾恢复 0.86±0.01 6.19±0.13 0.14±0.01 1.00∶0.16∶1.16
下载: 导出CSV
-
[1] 胡润芳. 香蕉施用钾肥对生长和产量的影响[J]. 福建果树, 1997(4): 14-15. [2] 杨苞梅, 李进权, 姚丽贤, 等. 钾钙镁营养对香蕉生长和叶片生理特性的影响[J]. 中国土壤与肥料, 2010(1): 29-32. [3] 赖达德. 钾肥对香蕉生长和结果的影响[J]. 福建果树, 2000(3): 33. [4] 林电, 颜速亮, 常春荣, 等. 反季节组培香蕉氮钾肥料配比、施肥时期及其效应研究[J]. 热带作物学报, 2002, 23(2): 36-40. [5] 杨苞梅, 李进权, 姚丽贤, 等. 钾钙镁营养对香蕉产量、品质及贮藏性的影响[J]. 中国生态农业学报, 2010, 18(2): 290-294. [6] 樊小林, 梁有良, 王孝强, 等.香蕉营养与施肥[M]. 北京: 中国农业出版社, 2007: 70-76. [7] 谭宏伟. 香蕉施肥管理[M]. 北京: 中国农业出版社, 2010: 103-106. [8] 郭玉婷, 伏广农, 张新明, 等. 高州市香蕉施肥状况调查与分析[J]. 安徽农学通报, 2013, 19(20): 46-48. [9] 张新明, 王建武, 樊小林, 等. 民众镇香蕉园土壤肥力及施肥状况调查与分析[J]. 福建果树, 2004(4): 13-15. [10] 李瑞民, 杨苞梅, 梁华赐, 等. 氮磷钾缺素对香蕉生长、抽蕾及产量的影响[J]. 广东农业科学, 2011, 38(4): 72-73. [11] OKUMU M O, VAN ASTEN P J A, KAHANGI E, et al. Production gradients in smallholder banana (cv. Giant Cavendish) farms in Central Kenya[J]. Sci Hortic, 2011, 127(4): 475-481.
[12] TAULYA G. East African highland bananas (Musa spp. AAA-EA) ‘worry’ more about potassium deficiency than drought stress[J]. Field Crops Res, 2013, 151: 45-55.
[13] 李丰年, 曾惜冰, 黄秉智. 香蕉栽培技术[M]. 广州: 广东科技出版社, 1999. [14] FREITAS A S, POZZA E A, POZZA A A A, et al. Impact of nutritional deficiency on yellow Sigatoka of banana[J]. Aust Plant Pathol, 2015, 44(5): 583-590.
[15] SULLIVAN W M, JIANG Z, HULL R J. Root morphology and its relationship with nitrate uptake in Kentucky bluegrass[J]. Crop Sci, 2000, 40(3): 765-772.
[16] 徐梦莎, 朱高浦, 付贵全, 等. 氮磷钾缺乏对苗期仁用杏生长和养分吸收的影响[J]. 西北农林科技大学学报(自然科学版), 2017, 45(5): 81-90. [17] 张志勇, 王清连, 李召虎, 等. 缺钾对棉花幼苗根系生长的影响及其生理机制[J]. 作物学报, 2009, 35(4): 718-723. [18] 曹秀, 夏仁学, 杨环宇, 等. 沙培条件下磷、钾、钙亏缺对枳(Poncirus trifoliata)幼苗根系形态及营养吸收的影响[J]. 植物营养与肥料学报, 2014, 20(4): 981-988. [19] 潘艳花, 马忠明, 吕晓东, 等. 不同供钾水平对西瓜幼苗生长和根系形态的影响[J]. 中国生态农业学报, 2012, 20(5): 536-541. [20] 曾秀成, 王文明, 罗敏娜, 等. 缺素培养对大豆营养生长和根系形态的影响[J]. 植物营养与肥料学报, 2010, 16(4): 1032-1036. [21] 习金根, 韦艳明, 张小玲, 等. 缺乏不同营养元素对剑麻幼苗生长和养分含量的影响[J]. 热带作物学报, 2013, 34(3): 403-407. [22] 宁运旺, 马洪波, 许仙菊, 等. 氮磷钾缺乏对甘薯前期生长和养分吸收的影响[J]. 中国农业科学, 2013, 46(3): 486-495. [23] 王秀荣, 曾秀成, 王文明, 等. 缺素培养对大豆养分含量的影响[J]. 华南农业大学学报, 2011, 32(4): 31-34. -
期刊类型引用(1)
1. 许凯,马英,陈三雄,骆金初,黄伟城,郭微,王龙远. 广东德庆县香山森林公园黄牛木种群特征研究. 安徽农业科学. 2022(13): 125-129 .  百度学术
百度学术
其他类型引用(0)


计量
- 文章访问数: 1519
- HTML全文浏览量: 26
- PDF下载量: 1813
- 被引次数: 1
