
Construction of a gE/gI-deleted mutant strain of epidemic porcine pseudorabies virus and its biological characteristics
-
摘要:目的
为研制针对猪伪狂犬病病毒流行毒株的疫苗提供候选毒株。
方法构建针对猪伪狂犬病病毒流行毒株的gE/gI缺失重组转移质粒pMD-LA-RA及携带EGFP标记基因的重组转移质粒pMD-LA-EGFP-RA,将pMD-LA-EGFP-RA与伪狂犬病病毒流行毒株PRV AH进行同源重组,利用EGFP为筛选标记,获得携带EGFP基因的gE/gI基因缺失突变株PRV AH gE –/gI –/EGFP+,以此毒株与pMD-LA-RA进行第2次同源重组,筛选去除EGFP基因的gE/gI基因缺失突变株PRV AH gE –/gI –,并通过生长曲线、易感细胞连续传代和动物免疫评价其增殖能力、遗传稳定性及免疫原性。
结果通过2次同源重组,结合荧光观察、空斑纯化和PCR检测,成功获得了PRV AH gE –/gI –,经PCR鉴定、荧光观察及测序鉴定,证实该毒株gE和gI基因被成功缺失,且不携带EGFP标记基因。生物学特性研究结果表明,该毒株增殖能力与亲本毒株相当,遗传稳定性及免疫原性良好。
结论采用同源重组技术成功构建了免疫原性良好的猪伪狂犬病病毒流行毒株gE/gI基因缺失突变株,为研制针对流行毒株的基因缺失疫苗奠定了一定的基础。
Abstract:ObjectiveTo obtain a candidate vaccine strain against epidemic porcine pseudorabies virus.
MethodA gE/gI-deleted transferring plasmid pMD-LA-RA and a recombinant plasmid carrying EGFP gene were constructed according to the sequence of epidemic porcine pseudorabies virus. The homologous recombination was operated between pMD-LA-EGFP-RA and PRV AH, and then the recombinant mutant virus PRV AH gE –/gI –/EGFP+ was selected using EGFP as screening marker. In order to obtain the gE/gI-deleted mutant strain PRV AH gE –/gI – without the EGFP gene, the second homologous recombination was carried out between pMD-LA-RA and PRV AH gE –/gI –/EGFP+. The proliferation ability, genetic stability and immunogenicity of PRV AH gE –/gI – were evaluated by its growth curve, continuous passage in susceptible cells and animal immunization.
ResultPRV AH gE –/gI – was obtained through two homologous recombinations with fluorescence observation, plaque purification and PCR. The deletion of gE, gI genes and EGFP gene marker in PRV AH gE –/gI – was identified by PCR, fluorescence observation and sequencing. PRV AH gE –/gI – had similar proliferation ability to the parental strain and had good genetic stability and immunogenicity.
ConclusionA gE/gI-deleted mutant strain of epidemic porcine pseudorabies virus with good immunogenicity was constructed successfully, which provides a basis for the development of gene deleted vaccine targeting epidemic strains.
-
凡纳滨对虾Paneaus vannamei为世界三大养殖虾类之一。自1988年引入中国以来,迄今已有40多年的养殖历史,2020年全国养殖产量达到186.3万吨[1]。然而,随着集约化养殖的加强,由于养殖密度过高、过度投喂以及饲料营养不均衡引发的肠道损伤问题频发,由此导致对虾消化吸收功能下降、免疫能力和生长受损,养殖成功率和养殖效益下降。因此,开发绿色、高效的对虾肠道保健产品,成为保障凡纳滨对虾健康养殖的当务之急。
丁酸梭菌Clostridium butyricum (CB)属于芽孢杆菌科梭菌属,是一种专性厌氧的革兰阳性芽孢杆菌。自1933年被发现、1935年被分离及1992年引入我国以来,其促进肠道健康的效果已在多种水产养殖动物如罗非鱼[2]、石斑鱼[3]、鳗鲡[4]、鮸鱼[5]、鲤鱼[6]等中得到证实,其已知作用途径主要包括增强肠道黏膜屏障功能、调节宿主肠道菌群、调节肠道pH,促进肠道对营养物质消化吸收等。丁酸梭菌于2009年被我国正式批准作为新饲料添加剂使用,在当前国家养殖减抗、饲料替抗(生素)的大背景下,其开发和应用将受到持续关注。
近年来的研究表明,饲用丁酸梭菌能改善克氏原螯虾、罗氏沼虾、日本囊对虾的抗氧化功能,提高肠道消化酶活性,或改善肠道形态、调节肠道菌群结构[7-9],说明丁酸梭菌在虾类饲料中也具有潜在的应用价值。但将丁酸梭菌作为肠道健康产品应用于凡纳滨对虾的研究还比较缺乏。有研究表明,在饲料中分别添加丁酸梭菌活菌、芽孢、破碎菌体或活菌与发酵上清混合物,均能提高凡纳对虾生长性能,且丁酸梭菌活菌和破碎菌体皆可提高凡纳对虾对副溶血弧菌的抵抗力,综合考虑生长性能、抗病力及产品保质期,丁酸梭菌芽孢更具有应用价值[10-11]。为此,本研究拟选用一种丁酸梭菌芽孢制剂,研究其对凡纳滨对虾生长性能、血清生化指标、肠道菌群及肠道内短链脂肪酸含量的影响,为明确丁酸梭菌芽孢制剂在凡纳滨对虾中的应用效果及探讨其可能的作用途径提供更多理论依据。
1. 材料与方法
1.1 试验饲料配制
以进口鱼粉、小麦粉、去皮豆粕为主要蛋白源,精制鱼油为脂肪源,配制凡纳滨对虾基础饲料,基础饲料配方及其营养水平测定值如表1所示。丁酸梭菌菌株(DZNCB-1-105)分离自土壤,其芽孢制剂由广东大泽农生物科技有限公司提供,粉状,活菌数为5×108 CFU/g。分别将质量分数为0(对照)、0.050%、0.075%和0.100%的丁酸梭菌添加到基础饲料中,即饲料中的活菌数分别为0、2.50×105、3.75×105和5.00×105 CFU/g,各处理简称为对照组(CK)、 0.050% CB组、0.075% CB组和0.100% CB组。和其他饲料原料充分混合均匀后,用双螺杆挤压机制成直径为1.5 mm的4种试验饲料,于−20 ℃条件下保存。
表 1 基础饲料配方及营养组成(风干基础)Table 1. Feed formulation and nutrient composition of the basal diet (air-dry basis)项目 Item 成分 Ingredient w/% 原料 Ingredient 鱼粉 Fish meal 36.0 小麦粉 Wheat flour 25.5 去皮豆粕 Soybean meal 15.0 花生粕 Peanut meal 8.0 菜籽粕 Rapeseed meal 10.0 鱼油 Fish oil 1.0 大豆卵磷脂 Soybean lecithin 1.0 磷酸二氢钙 Ca(H2PO4)2 1.5 预混料1) Premix 2.0 合计 Total 100.0 实测营养成分 Measured nutrient 粗蛋白 Crude protein 41.5 粗脂肪 Crude lipid 6.5 粗灰分 Crude ash 9.5 水分 Mositure 10.0 1)每千克预混料含有:VA 4 000 000 IU,VD3 1 500 000 IU,VE 100 000 IU,硫胺素 10 g,核黄素30 g,B6 30 g,泛酸 60 g,烟酸 80 g,B12 0.04 g,生物素 0.2 g,叶酸 6 g,肌醇120 g,VC 150 g,MgSO4·7H2O 102.45 g,KI 0.08 g,ZnSO4·7H2O 4.05 g,MnSO4·5H2O 2.19 g,CoCl2·6H2O 0.5 g,FeSO4·7H2O 16.55 g,Na2SeO4 0. 004 g,Ca(H2PO4)2 251.67 g 1)The premix per kilogram provides the following: VA 4 000 000 IU, VD3 1 500 000 IU, VE 100 000 IU, thiamine 10 g, riboflavin 30 g, B6 30 g, pantothenic acid 60 g, nicotinic acid 80 g, B12 0.04 g, biotin 0.2 g, folic acid 6 g, inositol 120 g, VC 150 g, MgSO4·7H2O 102.45 g, KI 0.08 g,ZnSO4·7H2O 4.05 g, MnSO4·5H2O 2.19 g,CoCl2·6H2O 0.5 g, FeSO4·7H2O 16.55 g,Na2SeO4 0. 004 g, Ca(H2PO4)2 251.67 g 1.2 试验动物与饲养管理
凡纳滨对虾购于广东省湛江市某对虾养殖场,购回后放入广东恒兴集团“863”基地室内海水流水养殖系统中暂养2周,暂养期间每日投喂基础饲料4次。暂养期结束后,挑选480尾大小均匀、健康的幼虾,初始体质量为(1.42±0.02) g,将幼虾随机分成4组,每组设4个重复,每个重复30尾虾,置于流水养殖系统的350 L PVC水族箱中。分别投喂4种试验饲料,每日投喂4次(07:00、11:30、16:00和20:30),日投饲率4%~6%,每3 d根据虾的摄食情况调整投饲率。试验用水为经砂滤、臭氧消毒的天然海水,水温(28±1) ℃,盐度(22±2)‰。通过不定期进水、排污及充氧,保持溶氧为(8.0±0.2) mg/L,氨氮(0.04±0.003) mg/L。养殖试验持续30 d。
1.3 样品采集与分析
1.3.1 生长性能
养殖试验结束时,每个水族箱的虾称质量、计数,统计饲料投喂量,计算各处理组对虾的存活率、质量增加率和饲料系数。
$$ 存活率=收获尾数/放养尾数 \times 100{\text{%}}, $$ $$ 质量增加率=(末质量-初质量)/初质量 \times 100{\text{%}} ,$$ $$ 饲料系数=投喂量/(末质量-初质量) 。$$ 1.3.2 全虾及基础饲料的营养组成
每箱随机取6尾虾,作为1个重复,保存于−20 ℃条件下用于检测全虾体成分。分别采用105 ℃恒温干燥法、凯氏定氮法、索氏抽提法及550 ℃灼烧法测定全虾及基础饲料的水分、粗蛋白、粗脂肪和粗灰分含量。
1.3.3 血清生化指标
每箱随机取9尾虾,使用1 mL注射器,从围心腔抽取血淋巴,4 ℃静置2 h后,使用冷冻离心机(Eppendorf,5425R)10 000 r/min离心10 min,取上层血清。每3尾虾的血清混合成1个样品,作为1个重复,保存于−80 ℃条件下待测,每个处理12个重复。血清生化指标用全自动生化分析仪(HITACHI,7170型)根据专用试剂盒说明书进行检测。
1.3.4 肠道菌群组成
每箱随机取6尾虾,在冰盘上取出肠道内容物,混合后置于冻存管,作为1个重复,迅速保存于−80 ℃条件下待测,每个处理4个重复。用DNA Kit(Omega Biotek)试剂盒提取肠道细菌基因组DNA。取合格样本DNA扩增16S rDNA序列的V4可变区。扩增引物为515F(5'-GTGC-CAGCMGCCGCGG-3')和806R(5'-GGACTACH-VGGGTWTCTAAT-3')。将PCR产物进行定量检测,之后按照每个样品的测序量要求,进行相应比例的混合,构成测序样品文库。由Illumina MiSeq测序仪采用MiSeq Reagent Kitv2(500-cycle)完成测序。采用Qiime平台和RDP classifier 2.2软件对97%相似水平的OTU代表序列进行分类学分析,置信度阈值为0.8,并在各个分类水平统计每个样品的群落组成。利用Mothur软件统计样品的α多样性,结果用Sob、Chao、Ace、Shannon以及Simpson多样性指数来表示。用QIIME v1.80软件统计分析样品的β多样性指数,采用主坐标分析(Principal coordinates analysis,PCoA)来进行数据统计分析,用PCoA散点图来表示不同处理之间的微生物群落物种组成的差异。
1.3.5 肠道内短链脂肪酸含量
每箱随机取6尾虾,在冰盘上取出虾肠道内容物,混合后置于1.5 mL无菌离心管中,作为1个重复,置于−80 ℃冰箱中保存,每个处理4个重复。取0.1 g样品于2.0 mL的Ep管中,加入400 μL体积分数为50%的乙腈溶液、200 μL的200 mmol/L 3-NPH及200 μL的120 mmol/L EDC溶液(含体积分数为6%的吡啶)涡旋1 min混匀,40 ℃反应1 h,期间每隔5 min震荡1次;反应完成后12 000 r/min、4 ℃离心15 min,取上清液上机进行LC-MS/MS (美国WATERS的液相色谱Waters Acquity UPLC 、美国SCIEX的质谱AB SCIEX 5500 QQQ-MS )。色谱分离条件:柱温40 ℃,流速0.30 mL/min;流动相组成:A为水(含φ为0.1%的甲酸),B为乙腈(含φ为0.1%的甲酸)。运行8 min,进样量6 μL。质谱分析条件:电子轰击电离(ESI离子源),离子源温度450 ℃,离子源气体35 arb。选择离子检测(SIM)进行定量分析。按照上述色谱及质谱条件进样,利用MultiQuant软件进行积分,利用标准曲线进行相对浓度计算。
1.4 数据统计
用统计软件SPSS 23.0进行数据的分析统计,试验数据用平均数±标准误表示。结果先进行单因子方差分析(One-way ANOVA),再通过 Duncan’s 多重比较分析数据之间的差异显著性。
2. 结果与分析
2.1 丁酸梭菌对凡纳滨对虾生长性能的影响
如表2所示,各组对虾的存活率为96.66%~100.00%。其中,0.100% CB组存活率达到100.00%。与对照组相比,0.050%和0.075% CB组对虾的末质量显著提高(P<0.05);其中0.050% CB组质量增加率也显著提高。饲料系数在丁酸梭菌各添加组有下降趋势,但和对照组相比没有统计学差异(P>0.05)。
表 2 饲料中添加丁酸梭菌(CB)对凡纳滨对虾生长性能的影响1)Table 2. Effects of dietary Clostridium butyricum (CB) on growth performance of Paneaus vannameiw(CB)/% 初质量/g Initial body weight 末质量/g Final body weight 存活率/% Survival rate 质量增加率/% Weight gain rate 饲料系数 Feed conversion rate 0(CK) 1.42±0.00a 7.03±0.14a 96.66±3.85a 394.53±9.51a 1.50±0.04a 0.050 1.41±0.00a 7.37±0.08b 97.50±1.67a 421.00±5.31b 1.44±0.04a 0.075 1.44±0.03a 7.37±0.03b 99.17±1.67a 410.43±8.65ab 1.45±0.08a 0.100 1.42±0.01a 7.31±0.07ab 100.00±0.00a 412.99±5.84ab 1.43±0.03a 1) 同列数据后的不同小写字母表示差异显著(P<0.05,Duncan’s 法) 1) Different lowercase letters of the same column indicate significant differences (P<0.05, Duncan’s method) 2.2 丁酸梭菌对凡纳滨对虾营养组成的影响
如表3所示,添加丁酸梭菌各组凡纳滨对虾的水分、粗蛋白、粗脂肪、粗灰分含量和对照组相比,均没有显著差异。
表 3 饲料中添加丁酸梭菌(CB)对凡纳滨对虾营养组成的影响1)Table 3. Effects of dietaryClostridium butyricum (CB) on nutrient compositions ofPaneaus vannameiw/% w(CB)/% 水分 Mositure 粗蛋白 Crude protein 粗脂肪 Crude lipid 粗灰分 Crude ash 0(CK) 272.93±14.82a 70.57±0.80a 7.33±0.20a 12.87±0.99a 0.050 261.06±7.97a 70.63±0.12a 7.28±0.11a 12.73±0.23a 0.075 280.64±2.11a 70.24±0.37a 7.12±0.36a 13.10±0.60a 0.100 285.98±12.38a 70.56±0.18a 6.36±0.90a 13.60±0.67a 1)同列数据后的不同小写字母表示差异显著(P<0.05,Duncan’s 法) 1) Different lowercase letters of the same column indicate significant differences (P<0.05,Duncan’s method) 2.3 丁酸梭菌对凡纳滨对虾血清生化指标的影响
如表4所示,和对照组相比,0.075% CB组对虾血清的葡萄糖浓度显著下降;该组胆固醇、甘油三酯浓度也低于其他各组,但没有统计学差异。随着丁酸梭菌添加量的升高,血清尿素氮浓度呈下降趋势,0.100% CB组显著低于对照组;而添加丁酸梭菌提高了血清磷浓度,在0.050% CB组达到显著性水平。总蛋白浓度在各组之间无显著差异。
表 4 饲料中添加丁酸梭菌(CB)对凡纳滨对虾血清生化指标的影响1)Table 4. Effects of dietaryClostridium butyricum (CB) on biochemical parameters in serum of Paneaus vannameiw(CB)/% c/(mmol·L−1) 总蛋白/ (U·L−1) Total protein 葡萄糖 Glucose 胆固醇 Cholesterol 甘油三酯 Triglyceride 尿素氮 Urea nitrogen 磷 Phosphorus 0(CK) 1.69±0.15b 2.21±0.23a 2.05±0.15a 1.23±0.17b 1.44±0.32a 64.38±3.13a 0.050 1.62±0.29ab 2.28±0.18a 2.09±0.16a 1.04±0.06ab 1.93±0.31b 66.43±2.94a 0.075 1.39±0.13a 1.83±0.18a 1.97±0.21a 1.11±0.14ab 1.79±0.24ab 66.53±3.83a 0.100 1.55±0.10ab 2.05±0.23a 2.03±0.19a 0.95±0.06a 1.80±0.26ab 65.47±3.72a 1) 同列数据后的不同小写字母表示差异显著(P<0.05,Duncan’s 法) 1) Different lowercase letters of the same column indicate significant differences (P<0.05, Duncan’s method) 2.4 丁酸梭菌对凡纳滨对虾肠道内容物菌群组成的影响
表5展示了各组丰度排名前10位的菌门。和对照组相比,丁酸梭菌各添加组浮霉菌门Planctomycetes的丰度显著升高,且在0.075% CB组达到最高;0.050% CB组髌骨菌门Patescibacteria的丰度也显著高于对照组。其余各菌门的丰度在组间差异不显著。
表 5 饲料中添加丁酸梭菌(CB) 对凡纳滨对虾肠道菌群组成(门水平)的影响1)Table 5. Effects of dietary Clostridium butyricum (CB) on intestinal flora composition (phylum level) of Paneaus vannameiw(CB)/% 变形菌门 Proteobacteria 拟杆菌门 Bacteroidetes 疣微菌门 Verrucomicrobia 浮霉菌门 Planctomycetes 厚壁菌门 Firmicutes 0(CK) 43.9±3.9a 41.9±3.2a 6.8±1.3a 2.8±0.4a 1.5±0.8a 0.050 36.0±2.0a 46.3±1.7a 6.7±0.7a 5.7±0.6b 0.5±0.2a 0.075 36.5±1.5a 43.0±3.5a 8.1±1.8a 7.1±0.5b 0.4±0.1a 0.100 43.2±4.3a 42.4±3.5a 5.7±1.2a 4.6±0.8b 0.7±0.3a w(CB)/% 放线菌 Actinobacteria 衣原体 Chlamydiae 髌骨菌门 Patescibacteria 埃普西隆杆菌门 Epsilonbacteraeota 柔膜菌门 Tenericutes 0(CK) 1.1±0.1a 0.8±0.4a 0.4±0.0a 0.2±0.1a 0.2±0.0a 0.050 1.9±0.7a 1.3±0.2a 1.1±0.3b 0.0±0.0a 0.0±0.0a 0.075 2.1±1.1a 1.7±0.7a 0.5±0.1a 0.0±0.0a 0.1±0.0a 0.100 0.8±0.1a 1.1±0.6a 0.3±0.0a 0.0±0.0a 0.2±0.1a 1)同列数据后的不同小写字母表示差异显著(P<0.05,Duncan’s 法) 1) Different lowercase letters of the same column indicate significant differences (P<0.05, Duncan’s method) 在属水平,和对照相比,0.050%和0.075% CB组弧菌属Vibrio丰度显著下降,假交替单孢菌属Pseudoalteromonas丰度有下降趋势;但后者在0.100% CB组丰度反而呈升高趋势。其余各属在组间差异不显著(表6)。
表 6 饲料中添加丁酸梭菌(CB)对凡纳滨对虾肠道菌群组成(属水平)的影响1)Table 6. Effects of dietary Clostridium butyricum (CB) on intestinal flora composition (genus level) of Paneaus vannameiw(CB)/% 弧菌属 Vibrio Motilimonas Haloferula 黏着杆菌属 Tenacibaculum Tamlana 0(CK) 7.8±3.6b 6.4±1.9a 6.3±1.3a 5.9±0.8a 5.3±0.7a 0.050 1.2±0.3a 6.8±3.0a 6.2±0.8a 4.3±0.3a 7.6± 0.7a 0.075 1.1±0.2a 8.5±1.6a 7.7±1.8a 5.1±0.4a 6.1± 0.8a 0.100 3.8±2.5ab 6.4±1.0a 5.0±1.2a 4.1±0.7a 6.7±0.5a w(CB)% 假交替单胞菌属 Pseudoalteromonas 鲁杰氏菌属 Ruegeria Hoppeia 康氏菌属 Kangiella Halocynthiibacter 0(CK) 2.6±0.5ab 2.6±0.3a 2.5±0.8a 2.2±1.5a 1.6±0.5a 0.050 1.5±0.3a 3.4±0.5a 4.1±0.4a 2.0±0.5a 1.1±0.3a 0.075 2.3±0.4a 4.1±0.9a 3.9±0.8a 0.8±0.2a 1.6±0.3a 0.100 3.7±0.6b 2.5±0.5a 4.7±2.1a 1.3±0.6a 1.4±0.1a 1)同列数据后的不同小写字母表示差异显著(P<0.05,Duncan’s 法) 1) Different lowercase letters of the same column indicate significant differences (P<0.05, Duncan’s method) 以97%序列相似度为阈值生成OTU后,计算不同样品的5个α多样性指数,并生成箱形图,以分析样品的α多样性(图1)。结果表明,Sob、Chao、Ace和Shannon多样性指数的变化趋势一致:与对照组相比,0.050%和0.075% CB组对虾肠道菌群的4项指数较高;当进一步将添加量提高到质量分数为0.100%时,4项指数数值均下降,但各组之间差异不显著。Simpson多样性指数亦在组间差异不显著。β多样性分析表明,同一处理样品基本聚为一簇,其中0.050%和0.075% CB组的样本距离较接近,而对照组和0.100% CB组与其他2组样本的距离较远,表明0.050%和0.075% CB组物种组成较相似(图2)。
![]() 图 1 基于5种多样性指数分析的α多样性箱形图图中空心圆点为各样本点Figure 1. The α diversity box plot based on analysis of five diversity indexesThe hollow dots in the figures represent individual samples
图 1 基于5种多样性指数分析的α多样性箱形图图中空心圆点为各样本点Figure 1. The α diversity box plot based on analysis of five diversity indexesThe hollow dots in the figures represent individual samples![]() 图 2 基于主坐标分析(PCoA)的各丁酸梭菌(CB)添加组肠道菌群组成散点图Figure 2. Scatter plot of intestinal flora composition in groups added with different amounts of Clostridium butyricum (CB) based on principal coordinates analysis (PCoA)
图 2 基于主坐标分析(PCoA)的各丁酸梭菌(CB)添加组肠道菌群组成散点图Figure 2. Scatter plot of intestinal flora composition in groups added with different amounts of Clostridium butyricum (CB) based on principal coordinates analysis (PCoA)2.5 丁酸梭菌对凡纳滨对虾肠道内短链脂肪酸含量的影响
随着丁酸梭菌添加量的升高,对虾肠道内5种短链脂肪酸的含量均有不同程度的升高(表7)。和对照组相比,乙酸含量在0.050%、0.075%和0.100% CB组分别升高了31.4%、98.1%和182.5%;丙酸含量升高了6.7%、40.0%和266.7%;丁酸含量升高了−10.0%、65.0%和60.0%;戊酸含量升高了9.7%、95.6%和140.7%,己酸含量升高了5.7%、68.6%和117.1%。可见,饲料中添加丁酸梭菌总体上大幅增加了肠内5种短链脂肪酸的含量。
表 7 饲料中添加丁酸梭菌(CB)对凡纳滨对虾肠道内短链脂肪酸含量的影响1)Table 7. Effects of dietaryClostridium butyricum (CB) on intestinal short-chain fatty acid contents ofPaneaus vannameiw/(mg·g−1) w(CB)/% 乙酸 Acetic acid 丙酸 Propanoic acid 丁酸 Butyric acid 戊酸 Pentanoic acid 己酸 Hexanoic acid 0(CK) 5.32±0.31a 0.30±0.04a 0.20±0.00a 1.13±0.02a 0.35±0.00a 0.050 6.99±0.24b 0.32±0.03a 0.18±0.01a 1.24±0.07a 0.37±0.01a 0.075 10.54±0.61c 0.42±0.03a 0.33±0.00b 2.21±0.14b 0.59±0.01b 0.100 15.03±0.31d 1.10±0.08b 0.32±0.01b 2.72±0.11c 0.76±0.01b 1)同列数据后的不同小写字母表示差异显著(P<0.05,Duncan’s 法) 1) Different lowercase letters of the same column indicate significant differences (P<0.05, Duncan’s method) 3. 讨论与结论
3.1 丁酸梭菌对生长性能的影响
丁酸梭菌对动物的促生长作用与其生物学特性密切相关。该菌属厌氧芽孢杆菌,在不良环境胁迫下能以孢子形式存在,具有更强的耐酸、耐胆盐特性,能够很好地在肠道定植而发挥作用。研究表明,丁酸梭菌能提高罗氏沼虾[12]、日本沼虾[13]和斑节对虾[14]等的生长性能,本研究中适宜数量的丁酸梭菌显著提高了凡纳滨对虾的生长性能,进一步表明虾等无脊椎动物也可有效利用饲料中的丁酸梭菌,提高生长性能。
3.2 丁酸梭菌对血清生化指标的影响
血清生化指标可以反映出机体能量代谢和营养物质吸收能力。血清总蛋白和白蛋白含量的升高是机体蛋白质代谢旺盛的体现[15],尿素氮是衡量动物体内蛋白质代谢和氨基酸平衡的一个重要指标,血清尿素氮的降低通常伴随着蛋白质沉积的升高[16]。在猪和肉鸡中的研究表明,饲料中添加丁酸梭菌可以显著降低两者血清尿素氮的浓度,并显著提高血清总蛋白、白蛋白和球蛋白的浓度[16-17]。本研究也发现,添加丁酸梭菌可显著降低对虾血清尿素氮浓度,0.050%和0.075% CB组的总蛋白浓度也有升高趋势;同时,还观察到血清磷浓度的显著升高。这可能是因为进入肠道的丁酸梭菌在肠道产生的短链脂肪酸促使肠道酸度增加,从而有利于某些有益菌的生长繁殖,增强消化酶活性,提高营养物质尤其是蛋白质及矿物元素的消化率[16]。血糖水平在一定程度上可以反映机体的合成代谢情况。本研究发现,饲料中添加质量分数为0.075%的丁酸梭菌显著降低了对虾血糖水平。类似的研究包括,丁酸梭菌可以降低糖尿病小鼠的空腹血糖,改善糖尿病及相关代谢紊乱[18];饲料中添加丁酸梭菌可降低猪对感染大肠埃希菌的应激反应,降低血糖水平[19]。但也有研究表明,饲料中添加丁酸梭菌能提高犊牛的生长性能和血清葡萄糖含量[20]。有关丁酸梭菌调节动物糖代谢的机制及效果,还需要更多的研究数据支持。
3.3 丁酸梭菌对肠道菌群的影响
本研究中,凡纳滨对虾肠道第一、二优势菌分别是变形菌门和拟杆菌门,它们也是大多数研究中对虾肠道的常见优势菌[21-22],丰度并未受饲料中丁酸梭菌的影响,表明它们可能是对虾体内稳定的“核心菌群”[23]。和对照组相比,添加3种不同质量分数的丁酸梭菌均显著提高了对虾肠道内浮霉菌门的丰度。浮霉菌门的成员在水生系统中普遍存在,包含多种厌氧氨氧化细菌,是高氨污染系统脱氮的生态友好型微生物[24]。郁维娜等[25]的研究显示,健康凡纳滨对虾肠道中浮霉菌门丰度显著高于患病虾,表明浮霉菌门的增殖有利于提高对虾的健康水平。同时,饲料中添加质量分数为0.050%和0.075%的丁酸梭菌降低了对虾肠道弧菌属的丰度,对假交替单胞菌属的丰度有降低的趋势。弧菌属中的很多种类是水产养殖业中常见的病原菌,其在肠道以及水体中丰度的升高常被视为虾类弧菌病暴发的前奏[26]。Sumon等[27]的研究表明,丁酸梭菌可以抑制罗氏沼虾肠道内弧菌的生长;宋增福等[28]使用肠细胞模型评估丁酸梭菌对鳗弧菌黏附能力的拮抗作用,发现丁酸梭菌有效抑制鳗弧菌对鱼类肠道的黏附。假交替单胞菌是一种能产生多种胞外活性物质的海洋细菌属,为对虾养殖环境中的常见致病菌,有研究证实其具有较强的致腐能力,并且其在患病凡纳滨对虾中的丰度显著高于健康组[25]。可见,添加丁酸梭菌能降低肠道中这些潜在病原菌的丰度,这可能与丁酸梭菌能够通过改善肠道屏障功能来降低病原菌的入侵和定植,以及其含有的脂磷壁酸等益生成分抑制了病原菌对肠道细胞的黏附作用,或其代谢产物如短链脂肪酸的抑菌作用有关[29-30]。在本研究中,发现梭菌属并非肠道内容物的优势菌,推测丁酸梭菌对于肠道上皮细胞具有高黏附作用,大部分可定植在肠壁,而非直接进入肠腔,但需要通过细菌定植试验进一步证实。
3.4 丁酸梭菌对肠道短链脂肪酸的影响
丁酸梭菌可在动物肠道发酵,产生以丁酸为主的短链脂肪酸,它们对动物肠道的能量供应、肠黏膜屏障的维持、肠道高敏感和肠道动力的调节、免疫调节等具有重要作用[31]。本研究发现,随着丁酸梭菌添加量的升高,凡纳滨对虾肠道内容物中5种短链脂肪酸(乙酸、丙酸、丁酸、戊酸和己酸)的含量均显著升高,这与Duan等[9]在日本囊对虾Marsupenaeus japonicus中的研究结果一致。不同种类的短链脂肪酸或其衍生物对提高水产动物生长性能、调节营养代谢的作用也在多项研究中得到证实。如饲料中添加丙酸钠可增加凡纳滨对虾对能量和磷的表观消化率,调节其肠道微生物并促进凡纳滨对虾生长[32];在饲料中添加丁酸盐能够显著提高商业虾采食量,改善其生长[33];添加3%(w)的β−羟基丁酸可改善凡纳滨对虾的生长性能、肠道消化酶活性及免疫水平,并增加其肠道短链脂肪酸含量[34];在蓝蟹Salvelinus alpinus饲料中分别添加乙酸钠、柠檬酸钠、丁酸钠或2%(w)丙酸钠等有机酸,均能够促进蓝蟹生长,促进其蜕皮,并提高其存活率[35]。目前关于戊酸对水产动物肠道健康的影响研究甚少,但研究发现,饲料中添加戊酸酯可降低小鼠肠道艰难梭菌的存活数[36],且肠道菌群代谢产生的戊酸可防止小鼠的辐射损伤[37]。然而,本研究中也观察到,添加丁酸梭菌至0.100%(w)时,尽管肠道短链脂肪酸含量进一步升高,但对虾的生长性能指标并未继续升高,且肠道菌群组成和0.050%及0.075% CB组相比,发生了较大变化,表明短链脂肪酸的过量产生可能会使肠道pH过低,导致肠道发生酸胁迫,从而破坏肠道菌群平衡[38],影响对虾的生长性能。
综上,本研究证实,饲料中添加丁酸梭菌芽孢制剂可抑制肠道潜在病原菌的丰度、提高肠道短链脂肪酸的含量,改善肠道健康水平,从而改善营养物质的利用,提高凡纳滨对虾的生长性能。本试验条件下,丁酸梭菌芽孢制剂在饲料中的适宜添加剂量为2.50×105或3.75×105 CFU/g。
-
![]()
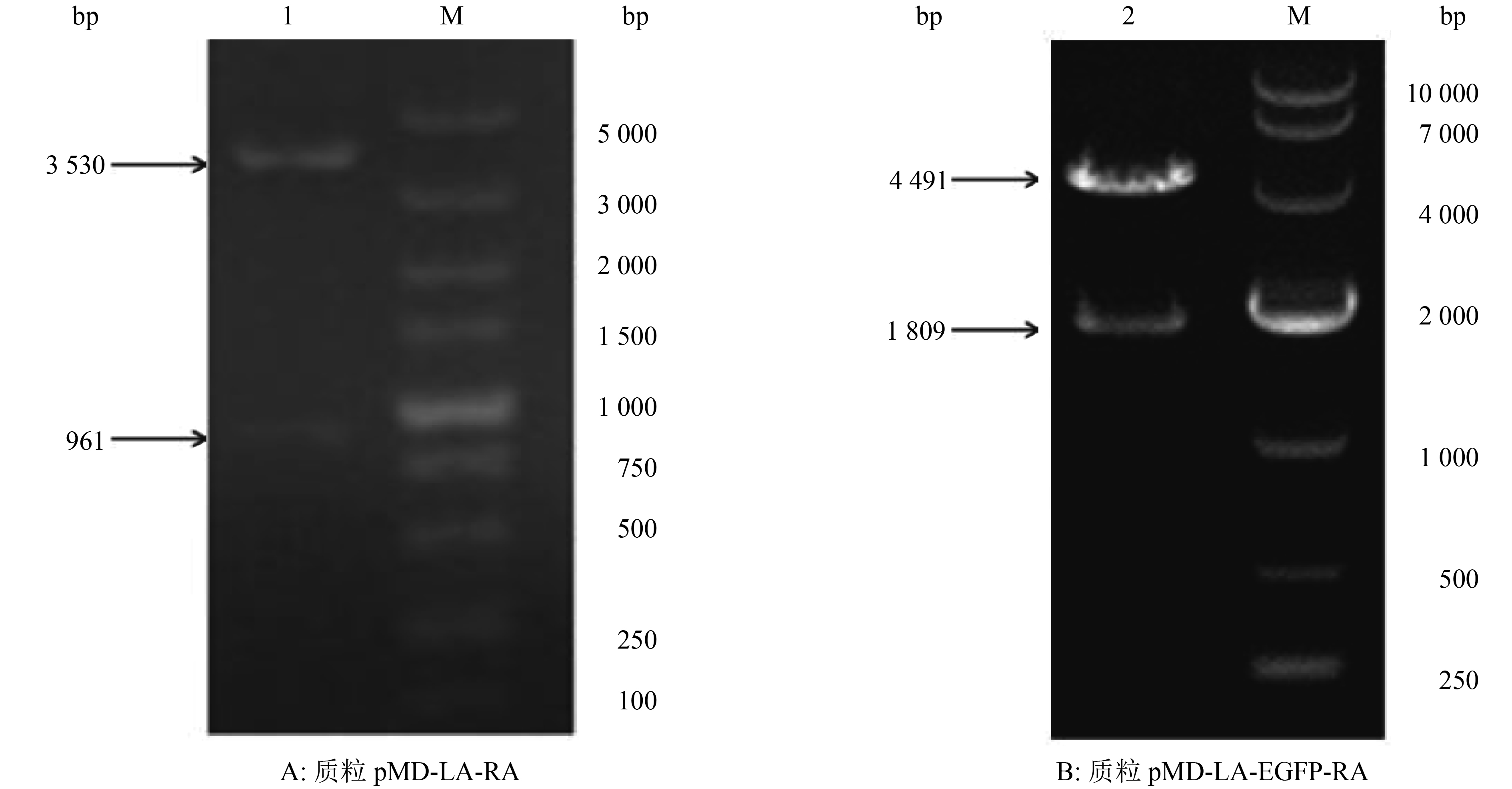
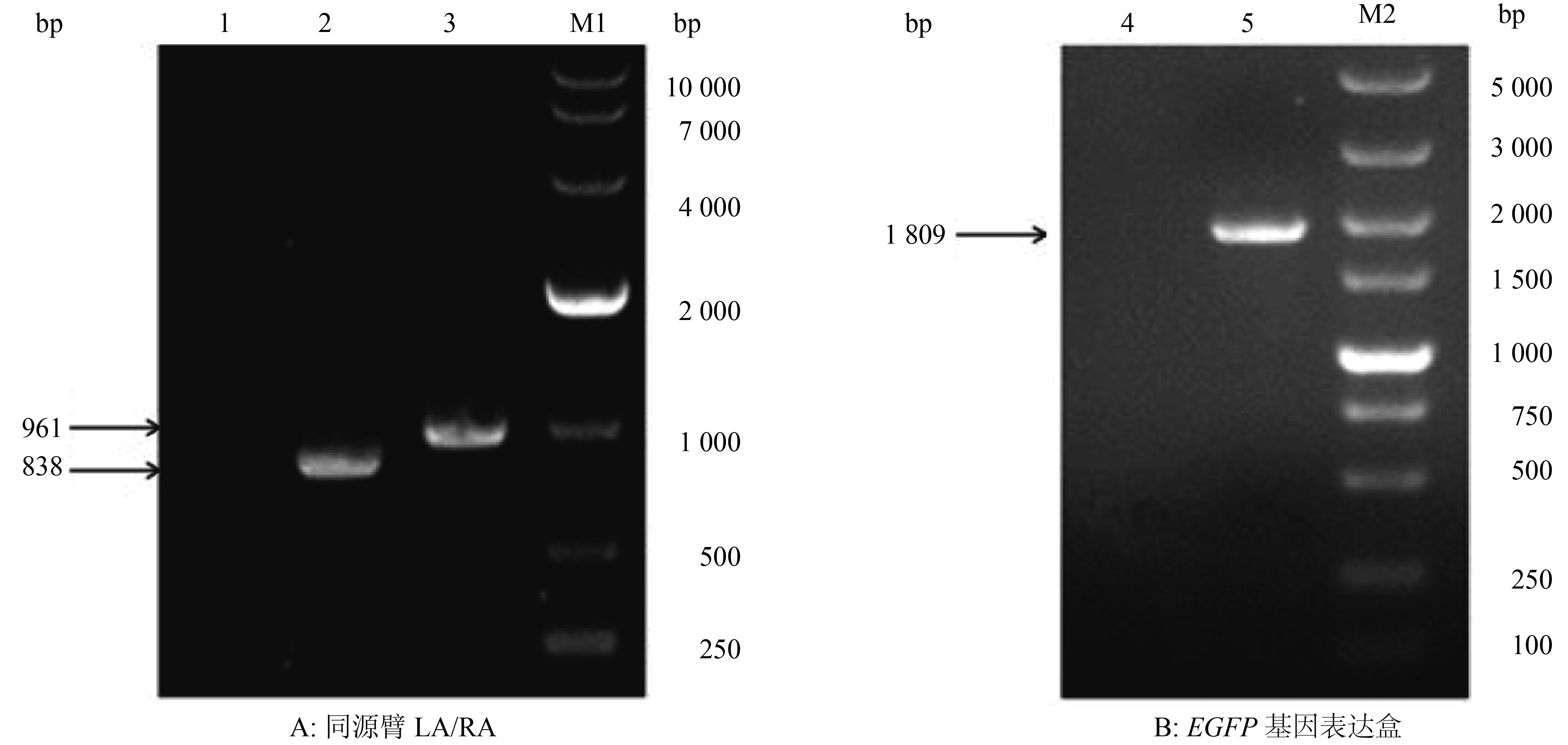
图 1 同源臂LA/RA与EGFP基因表达盒的PCR扩增结果
1:阴性对照;2:LA;3:RA;M1:DL10000 DNA Marker;4:阴性对照;5:EGFP基因表达盒;M2:DL5000 DNA Marker
Figure 1. PCR amplication of homologous arm LA/RA and EGFP gene expression cassette
![]()
图 2 重组质粒的酶切鉴定结果
M:DL5000 DNA Marker;1:PstⅠ+ Hind Ⅲ;2:EcoR Ⅴ
Figure 2. Identification of recombinant plasmid by restriction enzyme digestion
![]()
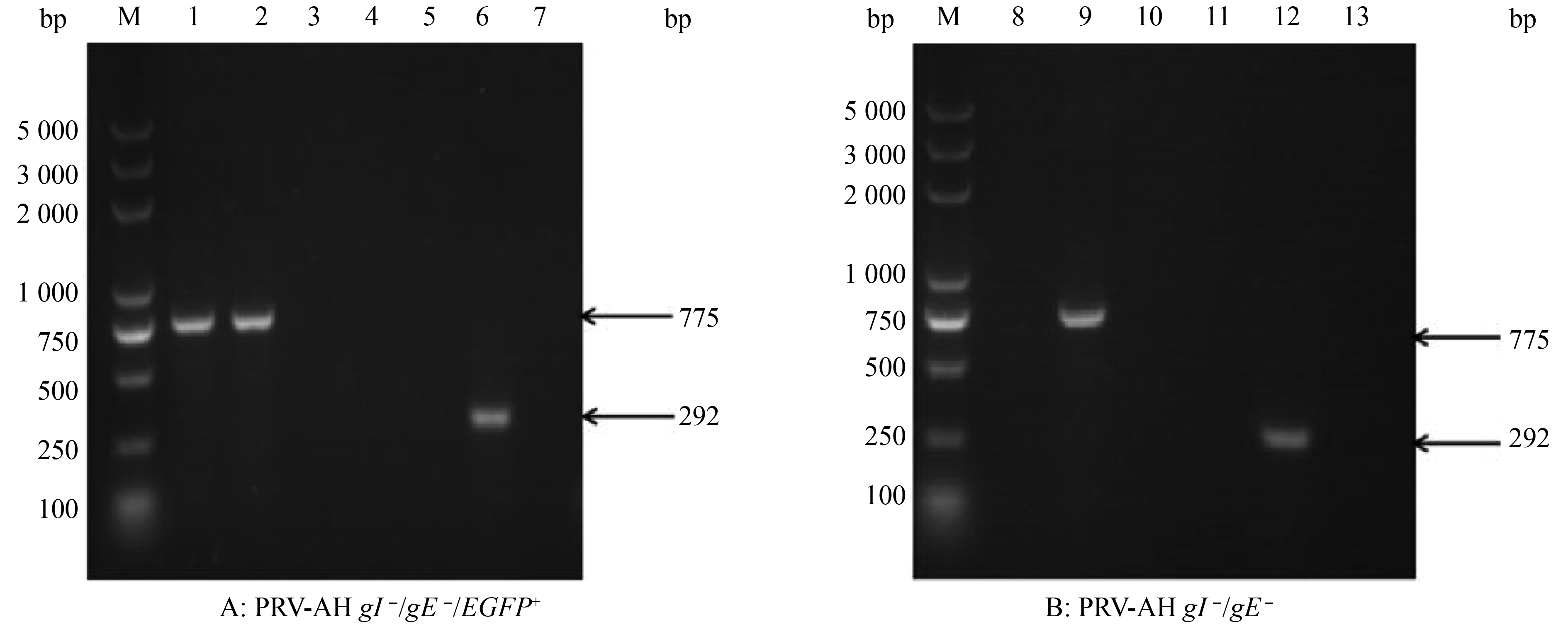
图 3 重组病毒PRV-AH gI –/gE –/EGFP+与PRV-AH gI –/gE –的PCR鉴定结果
M:DL5000 DNA Marker;1~4:EGFP扩增结果(775 bp);5~7:gE片段扩增结果(292 bp);1、5:PRV-AH gI –/gE –/EGFP+; 2:pEGFP-N1;3、6:PRV AH;4、7:阴性对照;8~10:EGFP片段扩增结果(775 bp);11~13:gE片段扩增结果(292 bp);8、11:PRV AH gI –/gE –;9:pEGFP-N1;10、13:阴性对照;12:PRV AH
Figure 3. Identification of recombinant virus PRV-AH gI –/gE –/EGFP+ and PRV-AH gI –/gE – by PCR
![]()
图 4 重组病毒感染BHK-21细胞的病变情况
A1、B1、C1:普通光观察PRV AH、PRV AH gE –/gI –/EGFP+及PRV AH gE –/gI –感染BHK-21细胞的病变;A2、B2、C2:荧光观察PRV AH、PRV AH gE –/gI –/EGFP+及PRV AH gE –/gI –感染BHK-21细胞的病变;A3、B3、C3:普通光与荧光观察结果的叠加图(merge);D1、D2、D3:未感染病毒的BHK-21细胞
Figure 4. Cytopathic effect of BHK-21 cells infected by recombinant viruses
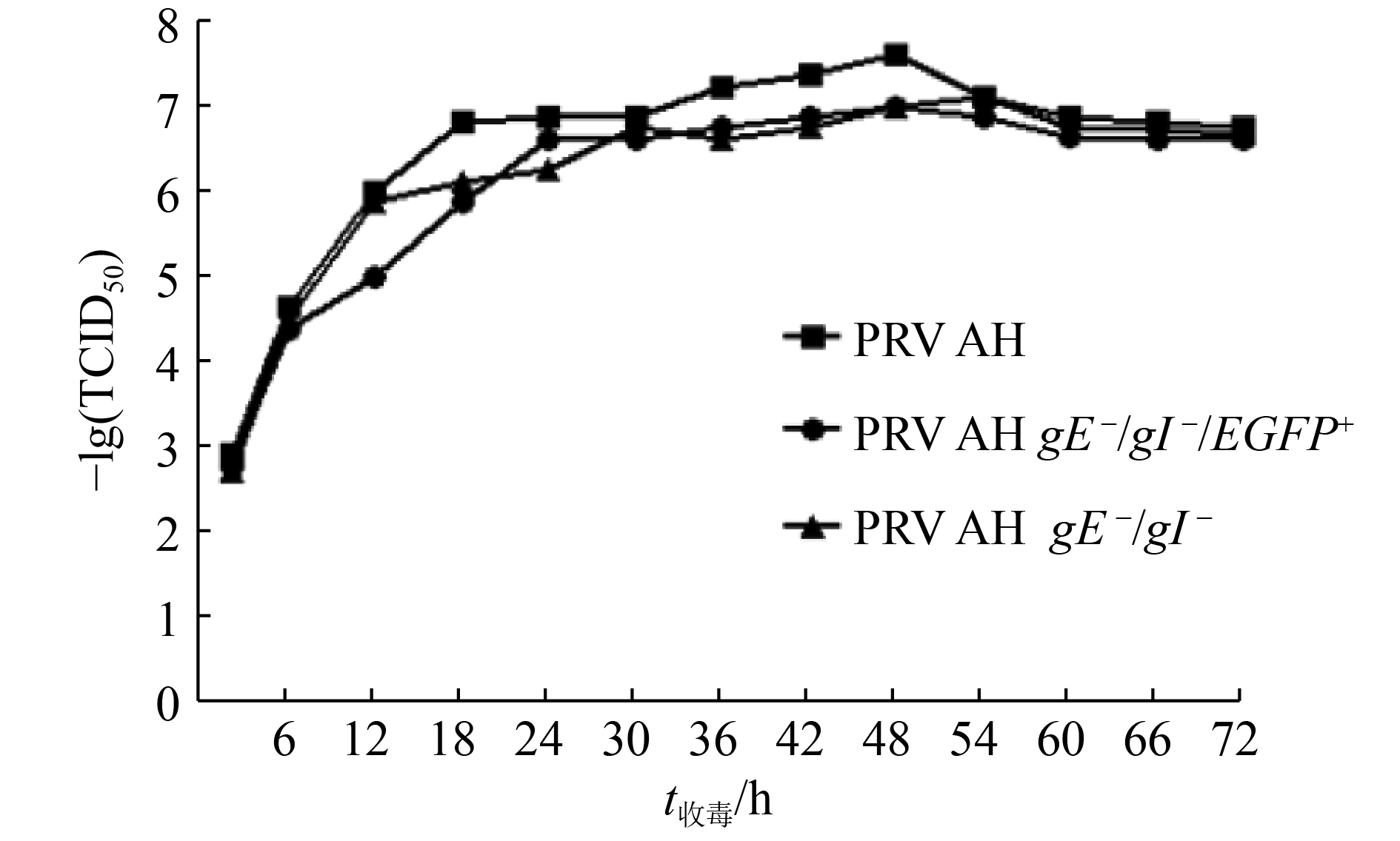
![]()
图 5 重组病毒和亲本病毒的生长曲线
Figure 5. The growth curve of recombinant viruses and parental strain
表 1 本试验所涉及的引物
Table 1 Primers used in this study
引物名称 引物序列(5′→3′) 扩增位置(碱基位点) 扩增长度/bp LA-F CCGGAATTCACCAGCACCGCACGTACAAGTT 120 511~121 349 838 LA-R CAGCAGCGTCCCGTCTATCGT RA-F AAACTGCAGGATATCCGGAAGTGACGAATGG 124 033~124 994 961 RA-R CTCGGTGGTGATGTAGAAAAGCTTGGG EGFP-F1 AACGATATCGTTTAAACGTTCTTTCCTGCGTTATCC 4 679~4 733;0~1 755 1 809 EGFP-R1 AACGATATCAACCCTATCTCGGTCTATTCT EGFP-F2 GTGGATAGCGGTTTGACTC 396~1 170 775 EGFP-R2 CACCTTGATGCCGTTCTT gE-F GTGATGACCCACAACGG 123 426~123 717 292 gE-R GCACGCAGAGCCAGAT LA-F2 AGTACCGGCGTCGATGATGA 121 144~121 348;
124 033~124 137310 RA-R2 GGTCAAACGTGTCCATGTCG  下载: 导出CSV
下载: 导出CSV
表 2 不同剂量PRV AH gE –/gI –灭活疫苗免疫小鼠后的中和抗体水平
Table 2 Neutralizing antibody titers of immunized mice with different dosages of PRV AH gE –/gI – inactivated vaccine
组别(免疫剂量)1) 首次免疫后不同时间的抗性水平 0 2周 4周 5周 6周 7周 8周 A(105.0TCID50) <2 <2 <2 <2 2.45±4.60 7.35±2.70 4.36±0.14 B(105.0TCID50) <2 <2 <2 <2 2.13±0.30 6.88±0.27 4.18±0.30 C(106.0TCID50) <2 <2 4.39±2.43 7.83±3.17 8.96±4.35 19.13±3.08 9.46±2.72 D(106.0TCID50) <2 <2 3.98±1.54 7.37±5.86 7.70±4.73 15.73±6.06 9.28±6.89 E、F、G(未免疫对照) <2 <2 <2 <2 <2 <2 <2 1) A、C、E为PRV AH株;B、D、F为PRV经典株;G为空白对照
下载: 导出CSV
-
[1] 童光志, 陈焕春. 伪狂犬病流行现状及我国应采取的防制措施[J]. 中国兽医学报, 1999, 19(1): 1-2. [2] 彭金美, 安同庆, 赵鸿远, 等. 猪伪狂犬病病毒新流行株的分离鉴定及抗原差异性分析[J]. 中国预防兽医学报, 2013, 35(1): 1-4. [3] 童武, 张青占, 郑浩, 等. 免疫后发病仔猪中伪狂犬病毒的分离和鉴定[J]. 中国动物传染病学报, 2013, 21(3): 1-7. [4] 叶培根. 猪伪狂犬病: 剖析猪伪狂犬病的新动向[J]. 北方牧业, 2013, 12: 23. [5] AN T Q, PENG J M, TIAN Z J, et al. Pseudorabies virus variant in Bartha-K61-vaccinated pigs, China, 2012[J]. Emerg Infect Dis, 2013, 19(11): 1749-1755.
[6] WU R, BAI C, SUN J, et al. Emergence of virulent pseudorabies virus infection in Northern China[J]. J Vet Sci, 2013, 14(3): 363-365.
[7] YU X L, ZHOU Z, HU D M, et al. Pathogenic pseudorabies virus, China, 2012[J]. Emerg Infect Dis, 2014, 20(1): 102-104.
[8] LUO Y Z, LI N, CONG X, et al. Pathogenicity and genomic characterization of a pseudorabies virus variant isolated from Bartha-K61-vaccinated swine population in China[J]. Vet Microbiol, 2014, 174(1/2): 107-115.
[9] 向柯宇, 潘慧, 吉艺宽, 等. 猪伪狂犬病病毒新流行株主要糖蛋白的分子特征分析[J]. 华南农业大学学报, 2016, 37(3): 23-28. [10] WANG C H, YUAN J, QIN H Y, et al. A novel gE-deleted pseudorabies virus (PRV) provides rapid and complete protection from lethal challenge with the PRV variant emerging in Bartha-K61-vaccinated swine population in China[J]. Vaccine, 2014, 32(27): 3379-3385.
[11] GU Z Q, DONG J, WANG J C, et al. A novel inactivated gE/gI deleted pseudorabies virus (PRV) vaccine completely protects pigs from an emerged variant PRV challenge[J]. Virus Res, 2015, 195: 57-63.
[12] TONG W, LI G X, LIANG C, et al. A live, attenuated pseudorabies virus strain JS-2012 deleted for gE/gI protects against both classical and emerging strains[J]. Antiviral Res, 2016, 130: 110-117.
[13] ZHANG C L, GUO L H, JIA X R, et al. Construction of a triple gene-deleted Chinese pseudorabies virus variant and its efficacy study as a vaccine candidate on suckling piglets[J]. Vaccine, 2015, 33(21): 2432-2437.
[14] HU R M, ZHOU Q, SONG W B, et al. Novel pseudorabies virus variant with defects in TK, gE and gI protects growing pigs against lethal challenge[J]. Vaccine, 2015, 33(43): 5733-5740.
[15] CONG X, LEI J L, XIA S L, et al. Pathogenicity and immunogenicity of a gE/gI/TK gene-deleted pseudorabies virus variant in susceptible animals[J]. Vet Microbiol, 2016, 182: 170-177.
[16] LIANG X, SUN L Q, YU T, et al. A CRISPR/Cas9 and Cre/Lox system-based express vaccine development strategy against re-emerging Pseudorabies virus[J]. Sci Rep, 2016, 6: 19176.






计量
- 文章访问数: 1847
- HTML全文浏览量: 15
- PDF下载量: 1922
