
Design of a grass separation equipment with driven rotating tooth disk
-
摘要:目的
设计1种从动型旋转齿盘分草装置,弥补现有分草装置结构复杂、清除率不高、对秸秆覆盖适应性差等不足。
方法优化设计1种由从动齿盘和分草栅条组成的分草装置;以秸秆覆盖高度及机具行走速度为影响因子对分草器进行EDEM仿真试验;进行田间通过性及种床带上秸秆清除效果试验。
结果仿真结果表明该分草装置可以在秸秆覆盖高度100~200 mm时稳定工作,清除率为49.0%~57.5%。随着机具前进速度和秸秆覆盖高度的增加,清除率增大。田间试验结果表明,当作业速度为3 km·h–1时,机具发生2次轻微堵塞,秸秆清除率为50.9%;当作业速度达到5 km·h–1时,机具无堵塞,通过性良好,秸秆清除率为57.5%。
结论该装置清草效果良好,可以满足小麦田间免耕作业的实际需要。
Abstract:ObjectiveTo design a grass separation equipment with driven rotating tooth disk, and resolve the disadvantages of existing grass separation equipment, such as the complex structure, low clearance rate and weak adaptability for straw coverage.
MethodA grass separation equipment with driven rotating tooth disk and grass allocation grids was designed. EDEM simulated experiments were performed with the impact factors of straw covering height and equipment forward speed. Field trafficability test was performed and the straw clearance effects on the seed bed belts were obtained.
ResultThe simulated result showed that this equipment worked steadily when the straw covering heights were from 100 to 200 mm, with the clearance rates ranging from 49.0% to 57.5%. Clearance rates increased with the increasing of equipment forward speed and straw covering height. The field experiment showed two slight congestions with 50.9% straw clearance rate at the operating speed of 3 km·h–1. At the operating speed of 5 km·h–1, the trafficability was good with no congestion and the straw clearance rate enhanced to 57.5%.
ConclusionThis equipment has good weed control effects, and it can meet the practical needs of no-till operation in wheat field.
-
Keywords:
- grass separation equipment /
- EDEM simulation /
- straw /
- clearance rate /
- no-till operation /
- trafficability
-
免耕播种机要求播种后地表有大量秸秆覆盖,但研究发现,种床带上秸秆覆盖量太大,对玉米的出苗有较大影响,同时也容易引起病虫害[1]。江苏地区采用小麦玉米轮作制,地表覆盖大量残茬秸秆,极易造成播种机开沟器的堵塞,分草装置的应用已成必然。齿式拨草装置具有转动灵活、除草效果好的特点,满足麦茬地玉米免耕播种机防堵分草的要求[2-3]。陈君达等[4]设计了一种轮齿防堵装置,其结构特点在于先将秸秆切断,然后利用拨草轮将秸秆两侧分开,2部分独立操作,配合分草。李洪文等[5]设计了一种对转爪拨压式防堵装置,其结构特点是通过齿轮啮合传递动力带动拨指工作,适用于秸秆含水量大的农田。卢彩云等[6]设计了一种斜置驱动拨草防堵装置,其结构特点是动力驱动2个拨草轮同向转动。范旭辉等[7]设计了仿形爪式机构,但仍处于研究中,不能适应秸秆覆盖量的变化,目前仍无法实体化。
本文在上述研究的基础上,利用爪式拨草轮的特点,优化设计一种结构简单的从动型旋转齿盘分草装置,以期解决驱动型分草器结构复杂、功耗大、不稳定等问题,完善从动式防堵装置对秸秆覆盖量适应性差的问题。结合江苏省麦茬地情况,分草装置配合尖角开沟器使用。
1. 分草装置的设计
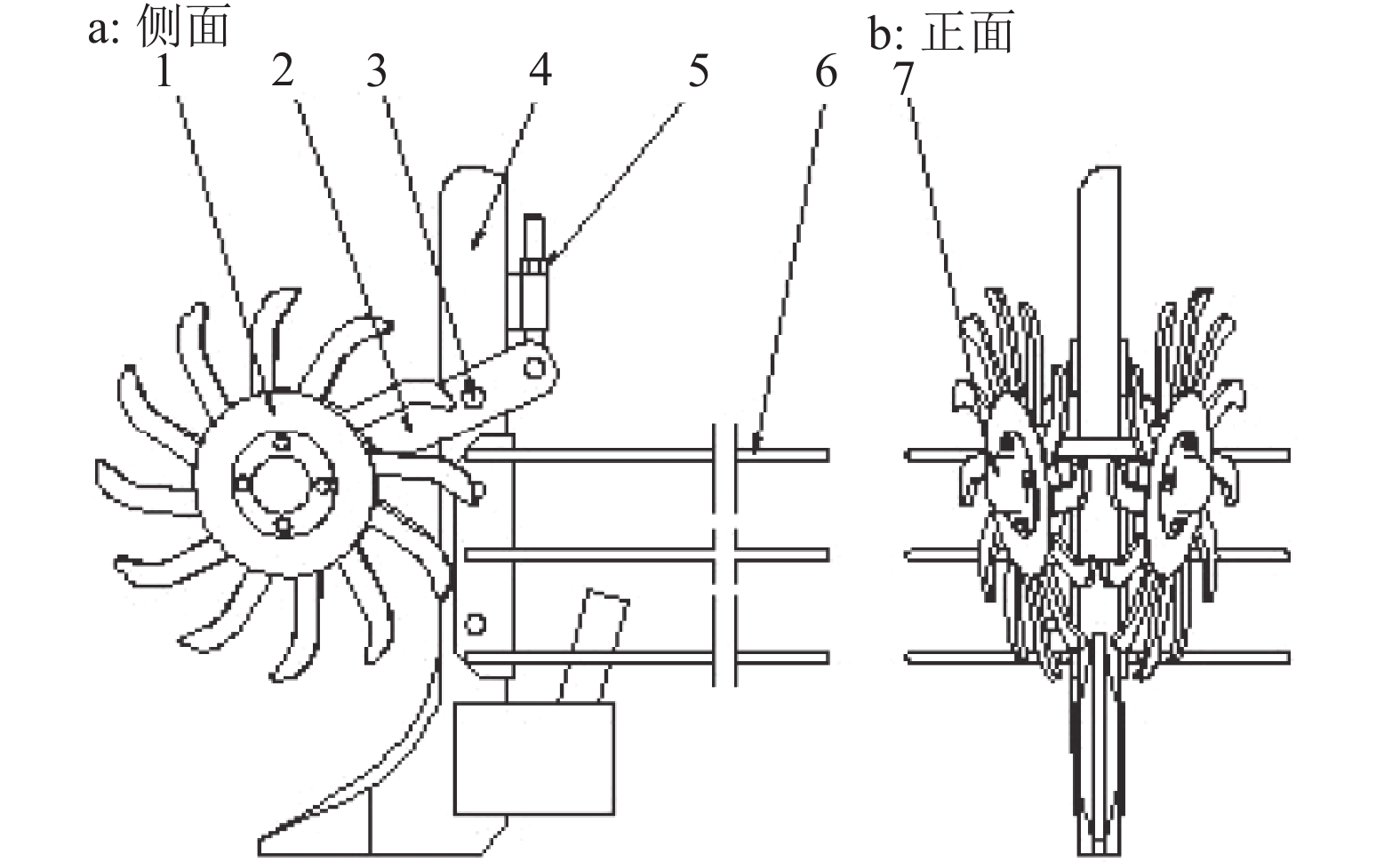
分草装置的作用是防止播种机具因秸秆而堵塞,保障播种作业的正常进行。本设计借鉴多齿拨禾星轮的原理,同时结合左右对称式双圆盘开沟器的排列方式对秸秆同时具有挑拨和侧分的功能。该机构(图1)包括2个12齿的齿盘,成八字型安装在连接摆臂上,齿盘前端相交于一点,且互不干涉。该机构安装于施肥开沟器的前端,通过销轴与施肥开沟器立柱铰接,末端安装有限位螺栓,可以调整齿盘与地面的高度。工作时,齿盘在重力的作用下,始终与地面接触。齿盘上的齿先进行压草,秸秆随齿端插入秸秆层与土壤接触,在拖拉机的牵引下齿盘转动,旋转的齿盘将前方秸秆推移、拨动、挑起,再向侧后方排放,在分草栅条的作用下被引向种床两侧,形成一条秸秆较少的种肥带。同时该装置可以清除较大的土块,提高播种机的通过性。
![]() 图 1 旋转齿盘分草装置1:齿盘;2:连接摆臂;3:销轴;4:开沟器立柱;5:限位调节螺栓;6:分草栅条;7:轴承及轴承盖Figure 1. Grass separating equipment with rotary tooth
图 1 旋转齿盘分草装置1:齿盘;2:连接摆臂;3:销轴;4:开沟器立柱;5:限位调节螺栓;6:分草栅条;7:轴承及轴承盖Figure 1. Grass separating equipment with rotary tooth1.1 分草齿盘的设计
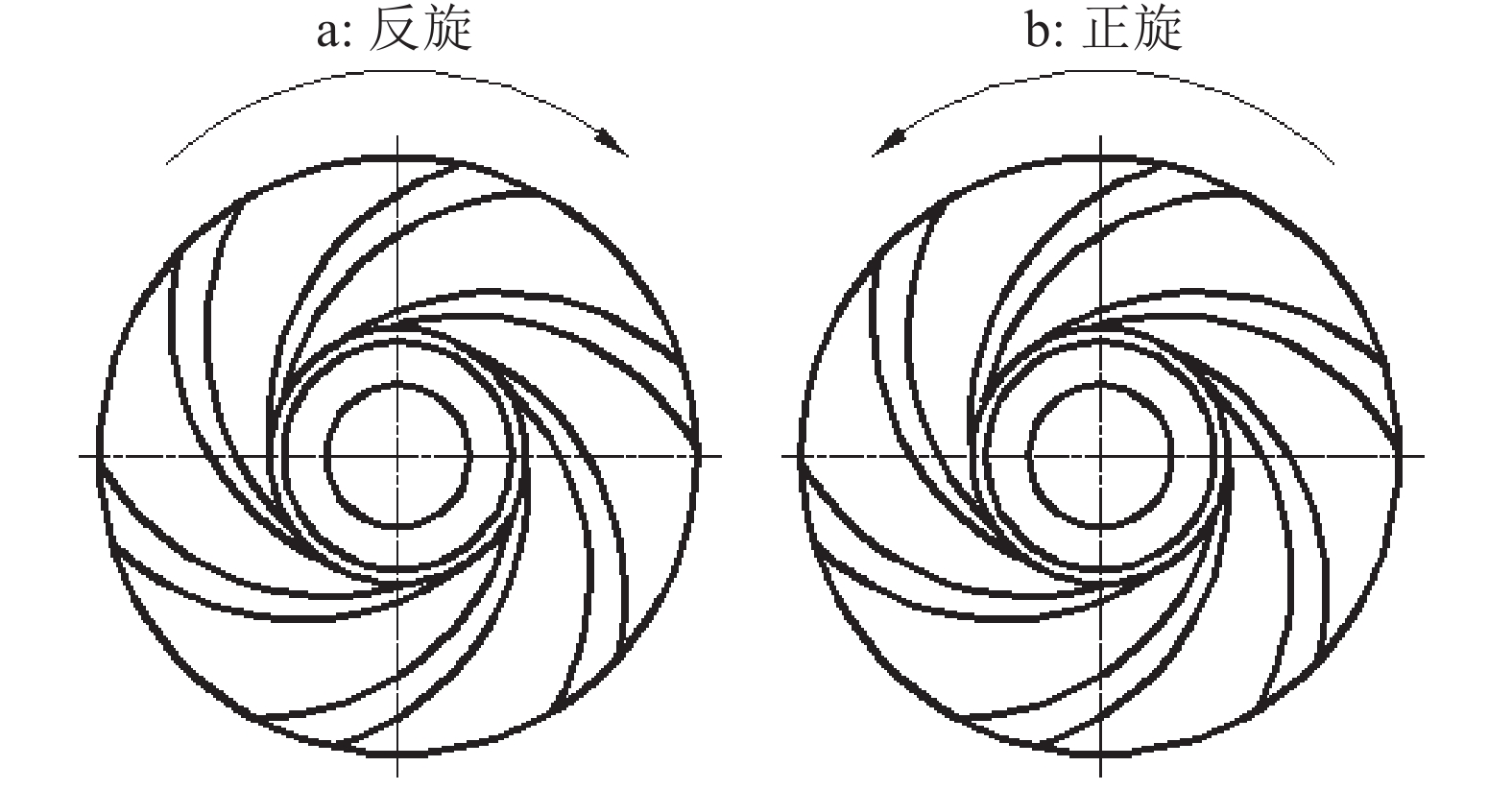
1.1.1 齿盘布置 旋转齿盘需要具有将秸秆层拨向两侧,并与秸秆分离的功能。齿盘结合星形轮曲线和开式叶轮水力曲线分为反旋和正旋(图2)。反旋的齿盘拨草效果较好,但不利于脱草,很容易造成堵塞。正旋的齿盘,只要角度适当,不但拨草效果不受影响,而且还便于秸秆与齿的分离,可防止秸秆的缠绕[8-11]。分草齿盘分草的作用过程是压草、拨草、挑起、排放,只有齿盘采用正旋方式,才能使分草的效果更好。故本研究齿盘采用正旋方式。
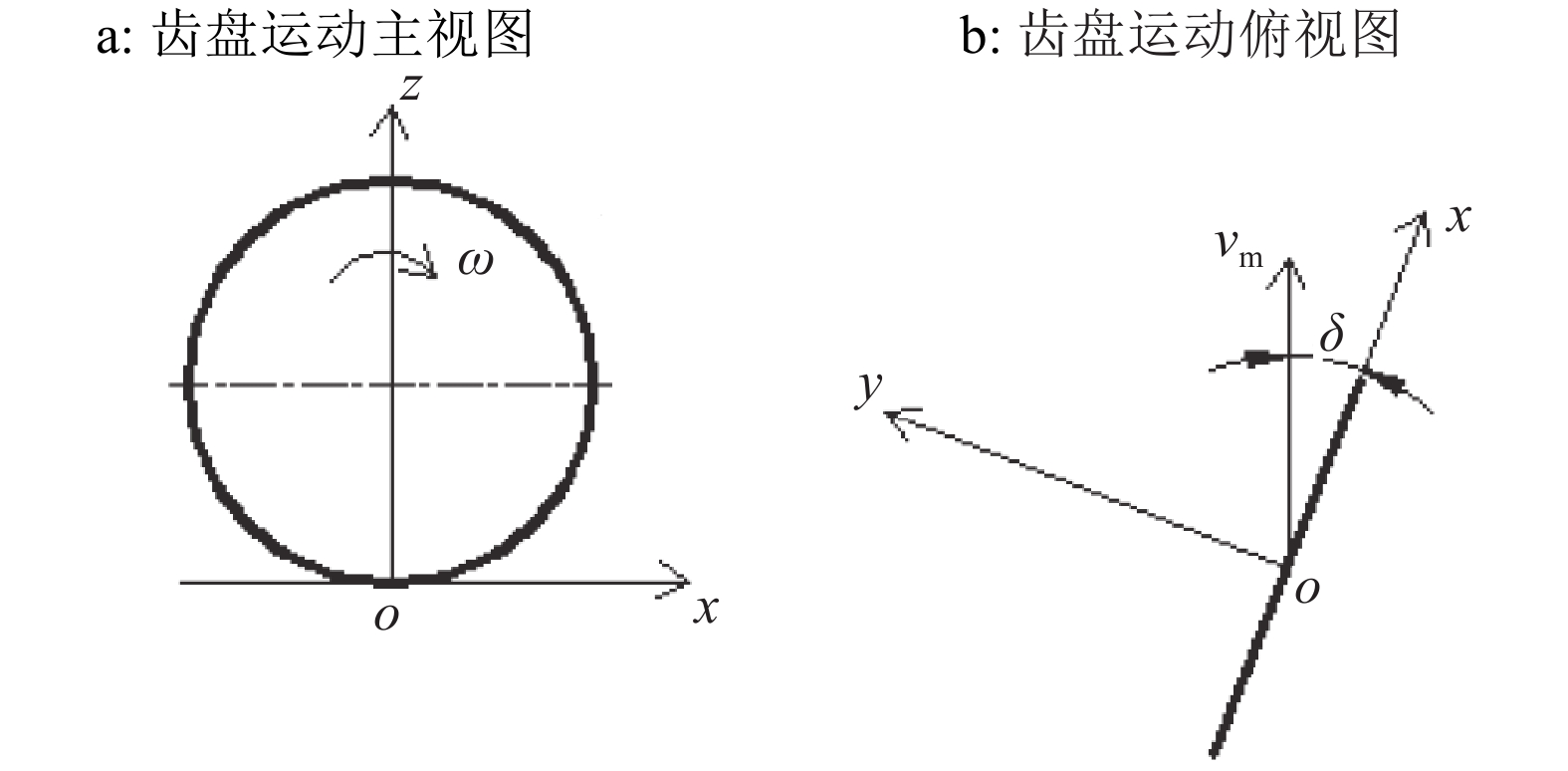
1.1.2 齿盘运动分析 由于运动偏角的存在,随着机具的前进,齿盘上轮爪的运动是一种复合运动[12-13],以轮爪与土壤的接触点为坐标原点,x轴和y轴分别平行和垂直齿盘平面,z轴垂直于x、y轴,平行齿盘平面向上建立坐标系(图3)。
其轮爪上任一顶点的运动方程为:
$$\left\{ {\begin{array}{*{20}{l}} {x = {v_{\rm m}}t\cos \delta + R{{\sin }\omega }t}\\ {y = {v_{\rm m}}t\sin \delta}\\ {z = R\left( {1 - {{\cos }\omega }t} \right)} \end{array}},\right.$$ 式中,vm表示机具前进速度,m·s–1;δ表示齿盘运动偏角,(°);ω表示齿盘旋转角速度,rad·s–1;R表示齿盘半径,m。
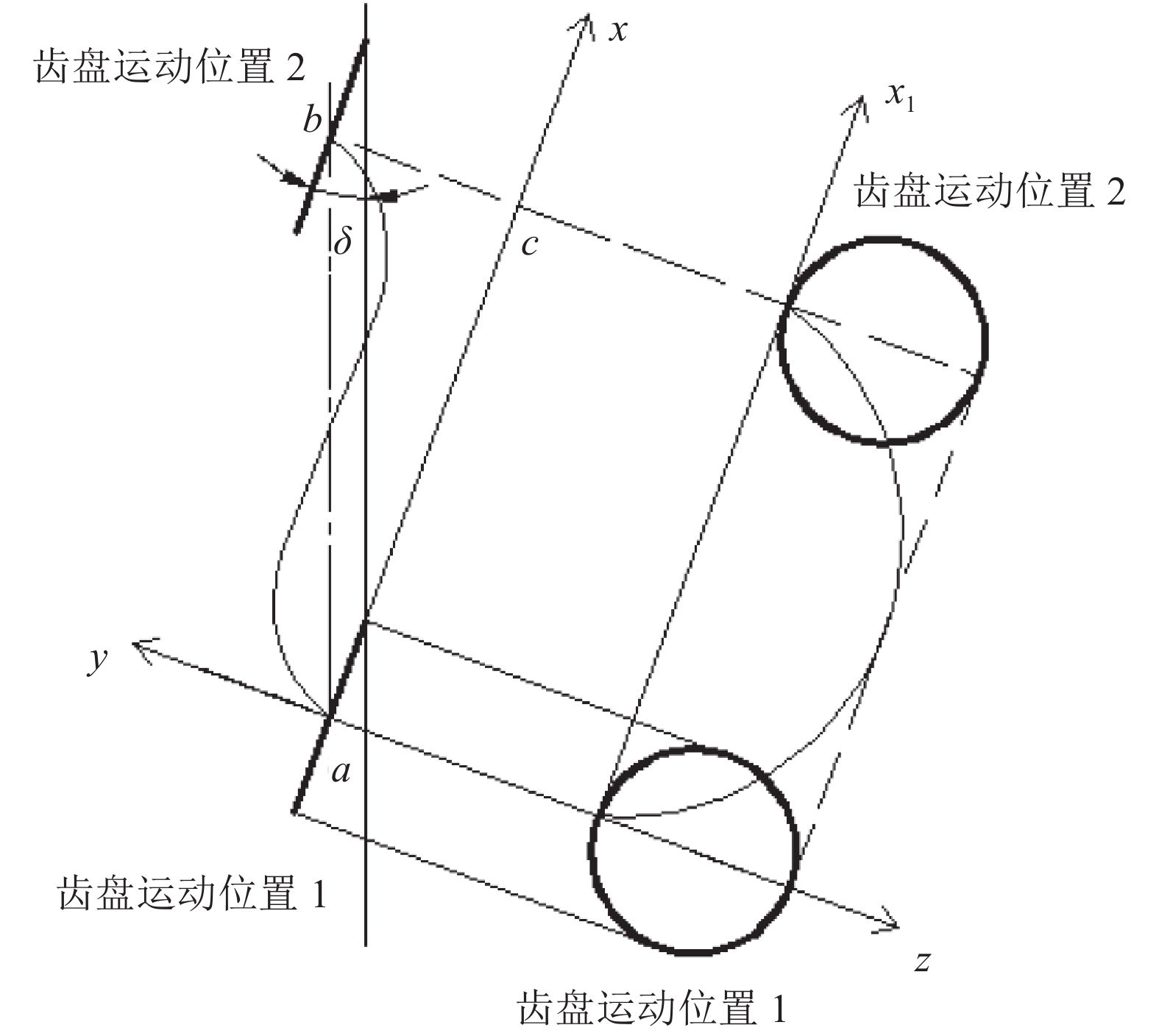
根据运动方程作出轮爪上任一顶点运动轨迹,将zoy面展开(图4)。由图4可知,轮爪顶点的运动轨迹是一条空间螺旋线,其运动可以看作是从a点到c点的纯滚动以及c点到b点的平移。从xoy面的运动轨迹可以看出,轮爪顶点横移明显,对秸秆有侧向推移的作用。增加运动偏角δ,对秸秆的侧推作用也随之变强,但对残茬的拨动能力减弱。运动偏角δ越小,对秸秆的拨动作用也相应变强。δ过大,会影响轮爪的转动;δ过小,轮爪对秸秆侧推能力较弱,分草效果不好。
![]() 图 4 轮爪任一顶点的运动轨迹位置1、2分别表示x-y平面、x-z平面Figure 4. The trajectory of any vertex of a wheel
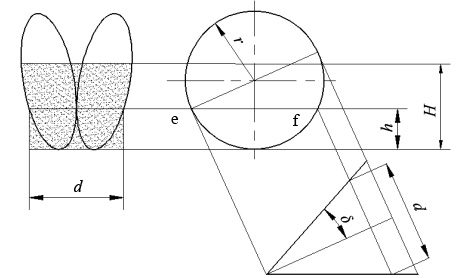
图 4 轮爪任一顶点的运动轨迹位置1、2分别表示x-y平面、x-z平面Figure 4. The trajectory of any vertex of a wheel1.1.3 齿盘相关指标 齿盘(图5)的啮合点高度(h)是影响拨草质量的重要参数之一。一般情况下,为了清秸干净,应使h等于或略小于秸秆覆盖厚度。h值太小,秸秆会从啮合点上方进入齿盘中间,造成堵塞;λ值太大会使轮爪在秸秆层中啮合不严,造成中间秸秆遗漏。小麦机收秸秆粉碎后均匀洒在地表,同时因秸秆可能覆盖在残茬上,秸秆覆盖高度约为100~200 mm。由于小麦经收割机收获后秸秆流动性较差且本设计要适用于秸秆高度在100~200 mm范围内不同覆盖量田间的清秸,故取h=100 mm,通过对运动偏角(δ)、齿盘直径(D)的调整,来实现设计要求。
![]() 图 5 齿盘结构示意图H为某瞬时秸秆覆盖高度(mm);h为齿盘的啮合点高度(mm);d为工作幅宽(mm);δ为运动偏角(°);r为齿盘在机具前进方向上的虚拟效应圆半径(mm);f为幅宽定义点;e为两齿盘啮合点Figure 5. Structure sketch of tooth disc
图 5 齿盘结构示意图H为某瞬时秸秆覆盖高度(mm);h为齿盘的啮合点高度(mm);d为工作幅宽(mm);δ为运动偏角(°);r为齿盘在机具前进方向上的虚拟效应圆半径(mm);f为幅宽定义点;e为两齿盘啮合点Figure 5. Structure sketch of tooth disc由图5可知工作幅宽(d)的计算公式为:
$$d = 2{l_{\rm {ef}}}\sin \delta = 4\sin \delta \sqrt {{r^2} - {{(r - h)}^2}} = 4\sin \delta \sqrt {2rh - {h^2}} ,$$ 式中,lef表示e点与f点的实际距离。
由于要保证种肥沟之间有超过4 cm的土壤隔离层,取工作幅宽d=200 mm。由图5可知啮合点在e处,应保证H–R<R–h,即200–R<R–100才能满足机具实际应用要求。将以上已知条件代入工作幅宽公式得出δ<20.7°。由于δ 过小,轮爪对秸秆侧推能力较弱,分草效果不好[14],将δ取整得20°以便于投产,则r=156 mm,综上,取齿盘直径D=332 mm。
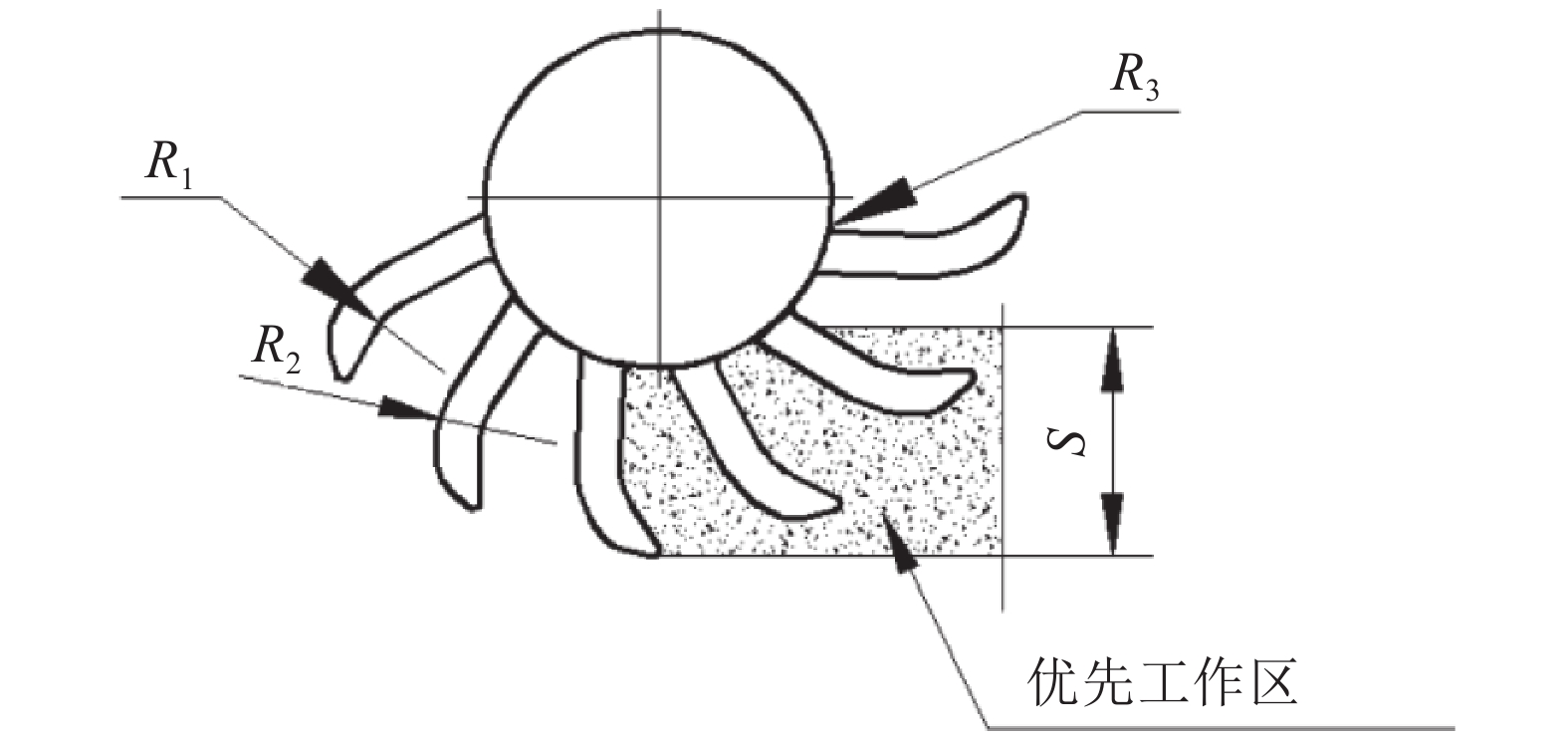
为了保证齿盘能连续不断地进行拨草,齿盘上要设定有一定数量的轮爪。轮爪的数量过多,轮爪难以插入秸秆层进行拨草,数量过少,需要很大的转速才能连续拨草。为使轮爪能连续地拨草,本研究的齿盘(通常8~14齿)参照多齿拨禾星轮的形状(图6),当秸秆覆盖高度较低时,应保证优先工作区覆盖高度(S)内至少有2个以上的齿盘正在工作。但随齿数的增多会增加齿盘的制造难度和制造误差[15-16]。利用CAD的1︰1绘型并综合以上限制条件取齿数12且均匀分布。为防止挂草和堵草,齿形各处用圆弧过渡。相邻齿根部用圆弧过渡,圆弧(R)应尽量大,以防止夹草。
1.2 分草栅条的设计
采用旋转齿盘式分草机构清扫施肥开沟器前方的秸秆,使秸秆不会缠绕或堆积在施肥开沟器上。但我国玉米播种机大都是3~4行,质量较轻,导致后端播种开沟器不足以切断较厚的秸秆或杂草等。因此,为了避免前方齿盘分离的秸秆因碰到残茬回落到种床带上,需要在齿盘后方安装一挡草装置。由图7可知,分草栅条通过螺栓安装在开沟器立柱上,紧随前方分草装置。该装置的使用既避免了秸秆被齿盘甩到种床上,同时将秸秆分流至相邻的种行间,保证种床带上秸秆量较小,从而方便双圆盘开沟器作业并减小病虫害的发生。
![]() 图 7 分草栅条位置示意图1:施肥开沟器立柱;2:分草装置;3:分草栅条Figure 7. Diagram of localization of grass allocation grids
图 7 分草栅条位置示意图1:施肥开沟器立柱;2:分草装置;3:分草栅条Figure 7. Diagram of localization of grass allocation grids2. 清秸效果模拟
利用EDEM2.6软件进行分草器清秸仿真。秸秆的仿真一直极有难度[17],方会敏等[18]将5个直径16 mm的颗粒堆成长条模型进行秸秆仿真。本研究为减少仿真误差,将6个直径8 mm、球心间隔6 mm的颗粒堆积成38 mm长的模型。仿真时所需的物理和力学性质参数根据参考文献[17-18]设定,具体仿真参数为:秸秆密度241 kg·m–3、秸秆泊松比0.4、秸秆剪切模量1×106 Pa、铁密度7 865 kg·m–3、铁泊松比0.3、铁剪切模量7.9×1010 Pa、秸秆–铁恢复系数0.30、秸秆–铁静摩擦因数0.30、秸秆–铁滚动摩擦因数0.01、秸秆–秸秆恢复系数0.30、秸秆–秸秆静摩擦因数0.30、秸秆–秸秆动摩擦因数0.25。
利用Solidworks建立分草齿盘与开沟部分模型,导入EDEM中。在EDEM中,由于秸秆被拨离工作区域以后无法再回落仿真区域,故不需要导入分草栅条结构。改变秸秆覆盖高度会使有效工作齿数改变,以机具前进速度(按玉米播种机一般作业速度取3、4、5 km·h–1)和秸秆覆盖高度(取高度100、120、140、160、180、200 mm)为影响因子得出18种秸秆清除率来考评分草器是否达到设计要求。
模拟试验所得秸秆清除率见表1,由表1可知,该装置随机具前进速度和秸秆覆盖高度的增加,清除率增大,从秸秆覆盖高度为100 mm的49.0%增加到200 mm的57.5%。
表 1 模拟试验不同覆盖高度的秸秆清除率Table 1. Straw clearance rates of simulation tests% 作业速度/
(km·h–1)覆盖高度/mm 100 120 140 160 180 200 3 49.0 49.2 50.0 50.3 50.8 51.0 4 53.0 53.1 53.2 53.5 53.7 53.9 5 55.2 55.5 56.0 56.6 56.9 57.5 3. 分草装置田间试验
3.1 试验条件
2016年6月在南京市江浦农场进行田间试验。该地雨水充沛,小麦长势较好,受其良好的气候环境影响,该地麦秸秆量每年均处于全省较高水平,因此选择在该地试验分草装置考察该装置的实用性及稳定性。作业地块是小麦秸秆经联合收割机作业粉碎后全部还田的麦茬地。
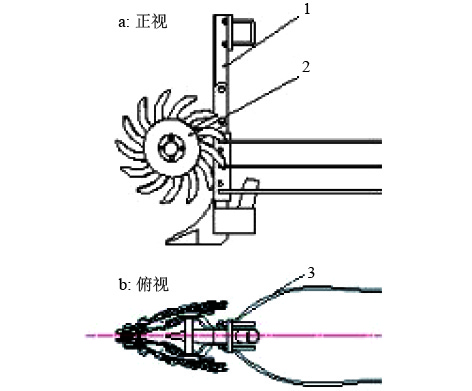
分草装置实体见图8,试验地地表覆盖大量秸秆,试验前对地貌进行了测定:小麦行距150 mm,地表留茬高度100~150 mm,秸秆覆盖量0.76 kg·m–2,土壤坚实度1 991 kPa,秸秆含水率(w)19%、秸秆长度100~150 mm,秸秆分布较均匀,秸秆覆盖高度148~170 mm。
3.2 试验内容
3.2.1 机具的通过性 以机具的作业速度为影响因素,设置前进速度为3、4、5 km·h–1 3个水平,每个水平往返3次,对机具的防堵性能进行测试。试验设计参照《免耕播种机质量评价技术规范》[19],设定测区长度为60 m,在往返1个行程内,按机具发生堵塞的程度进行分类:秸秆堵塞至不停机清理无法正常作业为重度堵塞;秸秆、根茬壅堵,不需停机清理为中度堵塞;秸秆、根茬壅堵,但堵塞物能自行滑落开沟器为轻微堵塞;机具不发生堵塞或发生1次轻微堵塞为合格。
3.2.2 秸秆清除率 试验前,在测定区随机选取10个点,测量单位面积内的秸秆量,取平均值作为秸秆覆盖量(
$X_1$ );试验后,在测定区有效清扫范围内随机选取10个点,测量单位面积内的秸秆量,取平均值作为秸秆残余量($X_2$ )。则秸秆清除率(η)为:$$\eta = \frac{{{X_1} - {X_2}}}{{{X_1}}} \times 100\text{%}\,\,{\text{。}} $$ 3.3 田间试验结果与分析
田间试验结果表明,机具在作业速度3 km·h–1时,发生了2次轻微堵塞,秸秆清除率为50.9%,但不需要停机清草,随着机具的前进,秸秆与齿盘逐渐分离,当作业速度增加到4 km·h–1以上时,无堵塞现象发生,通过性能良好,4、5 km·h–1的秸秆清除率分别为53.2%、57.5%,清草效果良好,作业后满足《免耕播种机质量评价技术规范》[19],结果表明该除草设备能满足实际工作要求。
仿真试验营造了一个相对苛刻的田间环境,因此在理论上仿真结果要低于田间试验,但本研究田间试验结果与仿真试验结果相近,因为田间机具作业时会存在秸秆间拉扯、麦茬干扰、地表不平、秸秆过长等问题。本研究结果表明从动型分草防堵装置随着作业速度的增加,秸秆清除率增加,后端分草栅条装置能够有效防止前方排放的秸秆回落到种床带上,为后续播种开沟作业创造了良好的条件。相对于市面已有的3种免耕播种机防堵装置[20],本装置秸秆清除率较高。
4. 结论
针对江苏省麦茬地作业特点,设计了一种从动型拨草齿盘式分草装置,并对该装置进行了清秸仿真试验与田间试验。清秸仿真试验表明该装置随机具前进速度和秸秆覆盖高度的增加,清除率增大,在作业速度为3~5 km·h–1,秸秆覆盖高度为100~200 mm时,清除率为49.0%~57.5%。田间试验表明当机具作业速度为3 km·h–1时,发生2次轻微堵塞,但仍可顺利通过,机具作业速度为5 km·h–1时,秸秆清除率可由3 km·h–1时的50.9%提高到57.5%。该装置通过性良好、清秸效果好、适应田间实际复杂情况且不需更改装置结构、工作稳定且功耗小,具有广阔的应用前景。
-
![]()
图 1 旋转齿盘分草装置
1:齿盘;2:连接摆臂;3:销轴;4:开沟器立柱;5:限位调节螺栓;6:分草栅条;7:轴承及轴承盖
Figure 1. Grass separating equipment with rotary tooth
![]()
图 4 轮爪任一顶点的运动轨迹
位置1、2分别表示x-y平面、x-z平面
Figure 4. The trajectory of any vertex of a wheel
![]()
图 5 齿盘结构示意图
H为某瞬时秸秆覆盖高度(mm);h为齿盘的啮合点高度(mm);d为工作幅宽(mm);δ为运动偏角(°);r为齿盘在机具前进方向上的虚拟效应圆半径(mm);f为幅宽定义点;e为两齿盘啮合点
Figure 5. Structure sketch of tooth disc
![]()
图 7 分草栅条位置示意图
1:施肥开沟器立柱;2:分草装置;3:分草栅条
Figure 7. Diagram of localization of grass allocation grids
表 1 模拟试验不同覆盖高度的秸秆清除率
Table 1 Straw clearance rates of simulation tests
% 作业速度/
(km·h–1)覆盖高度/mm 100 120 140 160 180 200 3 49.0 49.2 50.0 50.3 50.8 51.0 4 53.0 53.1 53.2 53.5 53.7 53.9 5 55.2 55.5 56.0 56.6 56.9 57.5  下载: 导出CSV
下载: 导出CSV
-
[1] 张永恩, 褚庆全, 王宏广. 中国玉米消费需求及生产发展趋势分析[J]. 安徽农业科学, 2009, 37(21): 10159-10161. [2] 李洪文, 陈君达, 邓健, 等. 旱地玉米机械化保护性耕作技术及机具研究[J]. 中国农业大学学报, 2000, 5(4): 68-72. [3] 李滨. 免耕播种机防堵技术存在的问题及发展趋势[J]. 当代农机, 2011(5): 68-69. [4] 陈君达, 高焕文, 王兴文. 免耕播种机轮齿式防堵装置: 98218325.9[P].1999–11–10. [5] 李洪文, 李慧, 王庆杰. 对转爪拨压式免耕防堵装置: 201110159818.4[P]. 2012–12–12. [6] 卢彩云, 赵春江, 孟志军. 斜置驱动拨草轮免耕防堵装置: 201410856945.3[P]. 2015–05–13. [7] 范旭辉, 贾洪雷, 张伟汉, 等. 免耕播种机仿形爪式防堵清茬机构参数分析[J]. 农业机械学报, 2011, 42(10): 56-60. [8] 孙彦波, 孙宏宇. 收割机割台拨禾星形轮的设计[J]. 机械工程师, 2008(6): 77-78. [9] 蓝文锋. 拨禾星轮的曲线[J]. 农业机械学报, 1982(1): 85-91. [10] 关醒凡.现代泵理论与设计[M].北京:中国宇航出版社, 2011: 314-324. [11] 李刚, 刘华明. 叶轮、叶片类复杂零件造型方法的研究[J]. 机械设计与制造, 2000(3): 59-60. [12] 陈钢, 晏世雷. 不同坐标系对质点运动的描述[J]. 物理与工程, 2011(3): 9-11. [13] 聂一鸣. 不同坐标系对质点运动的描述[J]. 科技经济导刊, 2016(26): 159. [14] 张国轩. 机械制造工艺分析[J]. 民营科技, 2013(8): 30. [15] 雷长贵, 李冠效. 机械加工精度的影响因素及提高措施[J]. 科技传播, 2014(5): 60. [16] 吴宗泽. 机械设计实用手册[M]. 北京: 化学工业出版社, 2003: 6. [17] LENAERTS B, AERTSEN T, TIJSKENS E, et al. Simulation of grain-straw separation by discrete element modeling with bendable straw particles[J]. Comput Electron Agr, 2014, 101: 24-33.
[18] 方会敏, 姬长英, AHMED A T, 等. 秸秆-土壤-旋耕刀系统中秸秆位移仿真分析[J]. 农业机械学报, 2016(1): 60-67. [19] 农业部农业机械试验鉴定总站, 甘肃省农业机械鉴定站. 中华人民共和国农业部. 免耕播种机质量评价技术规范: NY/T 1768—2009[S]. 2009: 1-12. [20] 杨彦光. 三种小麦免耕播种机性能试验及其效果对比研究[D]. 北京: 中国农业科学院, 2010.






计量
- 文章访问数: 1279
- HTML全文浏览量: 1
- PDF下载量: 1519
