
Finite element prediction and experimental verification for damage on citrus fruit from robot clamping
-
摘要:目的
研究作业机器人不同夹持条件下柑橘的内部应力变化,预测并分析柑橘的夹持损伤。
方法测定柑橘各组分的力学参数;根据柑橘结构建立其所对应的有限元模型,模拟机器人不同指面夹持柑橘的过程;利用自行设计的末端夹持平台对柑橘实物进行夹持验证试验。
结果柑橘的纵向果皮和果肉的弹性模量分别为11.408和0.277 MPa,极限应力分别为1.250和0.048 MPa。模拟试验中,得到柑橘果皮和果肉的等效应力分布图;柑橘果皮应力大于果肉;由于果肉的极限应力小于果皮,果肉的损伤先于果皮;相同夹持力下,弧指比平指对柑橘果皮和果肉作用应力小;平指夹持力为23 N时,果肉出现损伤,但弧指夹持力为45 N时,果肉才出现损伤;当夹持力分别为12、23、34、45、56、67 N时,平指比弧指夹持的柑橘果肉损伤率分别大0、10%、30%、40%、20%、20%。夹持试验结果验证了模拟预测试验。
结论可实现柑橘夹持损伤预测和评估,可为柑橘作业机器人的减损结构设计提供依据。
Abstract:ObjectiveTo study internal stress variations of citrus fruits clamped by working robot under different clamping conditions, and to predict and analyze clamping damages on citrus fruits.
MethodThe mechanical parameters of citrus peel and pulp were measured. Finite element model of each tissue structure of citrus fruit was established. The processes of clamping citrus fruits by robot with different finger planes were simulated. The verification experiments of citrus fruit clamping were performed using a custom terminal clamping platform.
ResultThe elastic modulus of citrus peel and pulp under vertical stress were 11.408 and 0.277 MPa respectively, and the limiting stress of citrus peel and pulp were 1.250 and 0.048 MPa respectively. The equivalent stress distribution diagrams of citrus peel and pulp were obtained in the simulation tests. The stress of peel was lower than that of pulp. Because the limiting stress of pulp was lower than that of peel, the pulp started to damage before the peel did. The stress on both peel and pulp caused by arc fingers was lower than that caused by plane fingers under the same clamping force. The peel started to damage at 23 N clamping force for the plane fingers, while the damage initiated at 45 N clamping force for the arc fingers. At the clamping force of 12, 23, 34, 45, 56 and 67 N, the pulp damage rates for the plane fingers were 0, 10%, 30%, 40%, 20% and 20% respectively higher than those for the arc fingers. The results of verification experiments using the clamping platform were consistent with the simulation outcomes.
ConclusionThe clamping damage on citrus fruit can be predicted and evaluated. This study provides a basis for designing damage reducing structures in citrus picking robot.
-
Keywords:
- citrus /
- robot clamping /
- mechanical parameter /
- damage prediction /
- finite element /
- surface form of finger
-
采用农业机器人对柑橘进行采摘、加工等作业时,常需要机器人末端执行器夹持果实,但果实被夹持时极易产生机械损伤。夹持所产生的损伤往往产生于果实内部,肉眼难以发现。通过受夹持柑橘内部应力和损伤特性分析,可减少夹持损伤并为作业机器人的减损结构优化设计提供依据。
国内外学者采用跌落、冲击和压缩等方法研究了果蔬在收获、贮运过程中的机械损伤,并分析了影响损伤的各种因素[1-6];也有学者采用有限元方法仿真模拟果蔬受载过程,研究果蔬内部应力分布和受载损伤的规律[7-13];对柑橘的力学特性和跌落损伤也有研究[14-16],上述研究均基于果蔬的减损控制和作业装备的优化设计,但目前鲜见有关柑橘力学特性和夹持损伤的研究报道。本研究通过测试柑橘各组分力学参数,构建其有限元模型,仿真模拟机器人夹持柑橘过程,分析受夹持柑橘内部应力分布规律,预测柑橘的夹持损伤,并采用自制末端夹持试验平台进行柑橘夹持验证试验,从而为减少柑橘的夹持损伤提供可控措施。
1. 材料与方法
1.1 材料
柑橘Citrus reticulate Blanco,品种为贡柑,橙与橘的自然杂交种,采摘于广东省广州市南沙果园,九成熟,其大小均匀、成熟度相近、完好无破损。肩部硬度为0.96 MPa(采用GY-1水果硬度计测量),直径为46.00~54.80 mm,质量为68.8~76.0 g。
柑橘组分力学参数测定所用设备为WD-20KE型精密微控电子万能试验机(广州市广材试验仪器有限公司),精度±5%,分辨率±1/120 000,试验过程可由微机自动完成力、位移和时间的数据采集。损伤预测验证采用自制末端夹持试验平台,试验用SF-400A电子台秤,精度0.1 g。
1.2 方法
1.2.1 柑橘组分力学参数测定
沿柑橘纵向取果皮拉伸试样10个,试样长40 mm,宽5 mm,平均厚度2.5 mm。采用单向拉伸试验[15],拉伸速率为10 mm·min-1。参考文献[11]求取果皮的拉伸极限应力σl(MPa)和拉伸弹性模量El(MPa):


式中,Flmax为试样失效时的拉力(N);b为试样宽度(mm);t为试样厚度(mm);Fl为拉伸力(N);ΔL为试样绝对伸长量(mm);L为试样原长(mm)。
为减少取样时对柑橘果肉细胞结构的破坏,试验时,把柑橘待测处果皮撕开,露出果肉,并固定于压力试验机的下压板上,采用半径4 mm刚性圆柱头以固定的加载速率10 mm·min-1垂直压缩果肉,试验重复10次。参考文献[17-18]计算果肉的弹性模量Ec(MPa)和压缩极限应力σc (MPa):


式中,Fc为压力(N);μ为物料的泊松比;D为变形量(mm);h为物料失效时压模的压入深度(mm);r为压模半径(mm)。
1.2.2 柑橘夹持损伤有限元预测
柑橘机器人本体由6自由度机械手和2指式末端执行器组成,通过6个伺服电机联合控制机器人工作。有限元预测分析首先需建立柑橘和夹持手指的接触几何模型,构建其有限元模型,然后模拟机器人手指夹持柑橘的过程。
建立柑橘有限元模型时,将果皮和果肉视为各向同性的线弹性材料,根据柑橘组分力学参数测定结果定义材料参数。柑橘实体模型采用四面体单元进行自由网格划分,果皮和果肉的单元类型分别选择Solid95和Solid92;材料单元边长分别为4和3 mm。
机器人手指夹持柑橘属于刚体与柔体间的接触,选择手指刚体的表面为目标面,采用TARGE170接触单元;选择柑橘表面为接触面,采用CONTA174接触单元。模拟夹持时,选取刚性目标面上的1个Keypoint作为控制节点,载荷加载在该控制节点上,通过控制节点Pilot控制整个刚性目标面的运动。
由前期的预备试验可知,当夹持力小于12 N时,柑橘没有出现机械损伤。为分析预测夹持过程中柑橘的内部损伤,采用平面和弧面2种指面形式,6个夹持力(12、23、34、45、56、67 N),分别进行夹持柑橘的有限元模拟试验,试验后通过柑橘的内部应力和应变图分析其损伤情况。
1.2.3 柑橘损伤预测验证试验
在自行设计的末端夹持试验平台上,采用部分夹持力(包含柑橘果皮和果肉出现损伤的夹持力)、平指和弧指,分别对柑橘实物进行夹持试验,每组试验数为5,试验重复5次。试验后通过测定柑橘的失质量率,以验证柑橘夹持损伤预测结果。夹持器通过电机驱动手指夹持柑橘,由压力传感器测量夹持力的大小。
经夹持后的柑橘放置于室温环境(20 ℃)中,一周后采用电子秤测定每组柑橘的质量,并计算其失质量率。
2. 结果与分析
2.1 柑橘组分力学参数
试验测得柑橘果皮、果肉的弹性模量分别为11.408、0.277 MPa;柑橘果皮、果肉的破坏应力分别为1.250、0.048 MPa,柑橘果皮、果肉建模所用泊松比参考文献[19]取值分别为0.42和0.47。
2.2 柑橘夹持损伤有限元预测
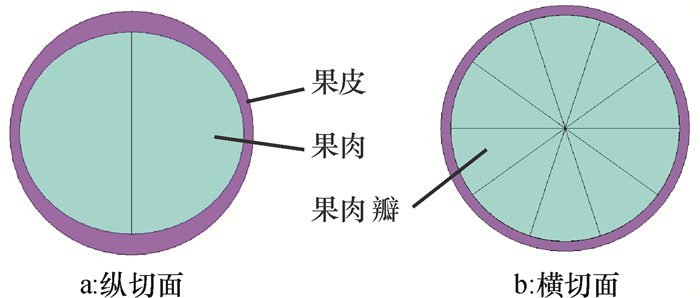
柑橘果实由外果皮、中果皮和多个外包瓢囊的汁胞等部分组成,如图 1所示。由于内、外果皮结合紧密,难以分开测定其力学参数,而瓢囊约8 ~ 12瓣,囊壁很薄。因此,将内、外果皮合成一体简化成果皮,而瓢囊与汁胞视为果肉,建模时,将柑橘看作是由果皮和果肉组成,模型内的果肉由10瓣组成。
通过前期试验测得柑橘球度系数为0.96,近似为球体。而由图 1a和实际测量可知,柑橘蒂部和顶部的果皮较厚,腰部较薄。因此,根据柑橘的实测平均几何尺寸,以外径为52.00 mm,果肉长轴为47.80 mm、短轴为42.72 mm,建立的柑橘几何模型如图 2所示。
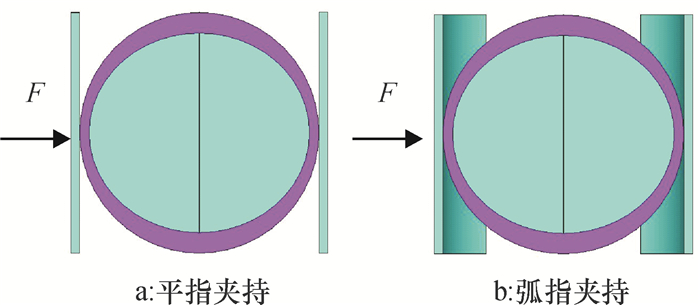
本研究采用平指和弧指(指面分别为平面和弧面)模拟机器人手指夹持柑橘,平指尺寸为55 mm(长)×52 mm(宽)×2 mm(厚),弧指尺寸为56.56 mm(长)×52 mm(宽)×2 mm(厚),模型中平指和弧指简化为薄的平面板和弧面板刚体,夹持力F指向柑橘腰部。由于柑橘结构的对称性,为减少计算机模拟分析所需时间,采用1/2柑橘对称模型进行研究,最终建立的机器人手指与柑橘的接触模型如图 3所示。
![]() 图 3 手指与柑橘夹持实体模型Figure 3. The solid models of citrus fruit clampied by different fingers
图 3 手指与柑橘夹持实体模型Figure 3. The solid models of citrus fruit clampied by different fingers2.3 有限元模拟夹持柑橘结果分析
2.3.1 不同指面夹持下柑橘内部应力
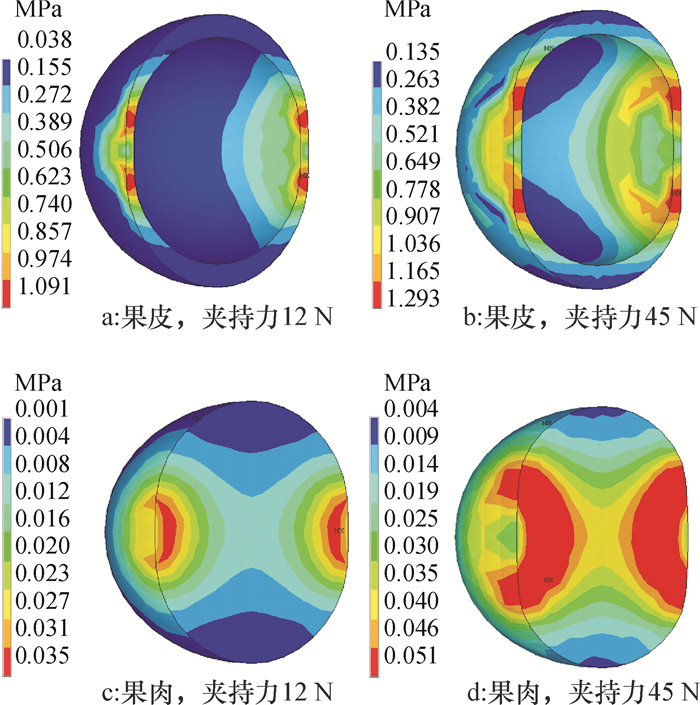
模拟试验后得到不同指面夹持下柑橘内部组织的等效应力分布图。比较可知,2种指面夹持下柑橘内部应力变化趋势相似,以平指夹持力12、45 N为例(图 4)说明:同一夹持力下,果皮的等效应力大于果肉;随着夹持力增加,柑橘果皮和果肉的最小和最大等效应力均增大,且最大等效应力面积也增大,同时往果肉中心方向扩展;手指与柑橘接触点两侧的应力最大,因此柑橘损伤首先出现在两侧。
![]() 图 4 平指夹持下柑橘的等效应力图Figure 4. Equivalent stress distribution diagrams of citrus fruit clamped by plane fingers
图 4 平指夹持下柑橘的等效应力图Figure 4. Equivalent stress distribution diagrams of citrus fruit clamped by plane fingers2.3.2 不同指面夹持下柑橘损伤预测
不同指面夹持下柑橘内部组织最大的等效应力见表 1,由表 1可知,采用平指,当夹持力为12 N时,柑橘果皮和果肉的最大等效应力分别为1.091和0.035 MPa,均小于各自的极限应力,不会对柑橘造成机械损伤;当夹持力为23 N时,果皮的最大等效应力仍小于其极限应力,而果肉的最大等效应力为0.049 MPa,其值虽然小于果皮的等效应力,但大于其极限应力0.048 MPa,果肉组织开始出现机械损伤;当夹持力增大到45 N时,果皮的最大等效应力为1.293 MPa,大于其极限应力1.250 MPa,说明此时除果肉产生机械损伤外,果皮也出现机械损伤。
表 1 柑橘夹持有限元模拟试验等效应力Table 1. The equivalent stress of finite element simulation of citrus fruit clamping
采用弧指,当夹持力分别为12、23和34 N时,柑橘果皮和果肉的最大等效应力均小于各自的极限应力;当夹持力为45 N时,果皮的最大等效应力为1.127 MPa,仍小于其极限应力,但果肉的最大等效应力为0.049 MPa,大于其极限应力,此时果肉组织首先出现机械损伤;试验范围内,果皮最大等效应力为1.229 MPa,接近其极限应力,果皮将出现机械损伤。
对比2种指面,相同夹持力下,弧指夹持柑橘,其果皮和果肉的最大等效应力和变形量均小于平指夹持的,柑橘出现损伤的概率低于平指夹持的。
2.3.3 不同指面夹持下柑橘果肉损伤率
由于柑橘果肉由多瓣构成,同一夹持力下,不同指面夹持的柑橘,其受损果肉瓣数不同。本文以等效应力值超过其极限应力的果肉瓣数统计为受损果肉瓣数,由此计算柑橘果肉损伤率,以进一步评估夹持力、指面形式对柑橘果肉损伤程度的影响。柑橘模型中果肉由10瓣组成,因此,果肉损伤率=(受损果瓣数/10)×100%,计算结果见表 2。由表 2可知,随着夹持力的增大,柑橘果肉损伤率增大,平指夹持的柑橘果肉损伤率高于弧指夹持,当夹持力分别为12、23、34、45、56、67 N时,平指夹持的柑橘果肉损伤率分别比弧指夹持的大0、10%、30%、40%、20%、20%,当夹持力达到67 N时,平指夹持的柑橘果肉损伤率为100%。
表 2 不同夹持力下柑橘果肉损伤率预测Table 2. Prediction in damage rate of citrus pulp under different clamping force
2.4 柑橘损伤预测验证试验
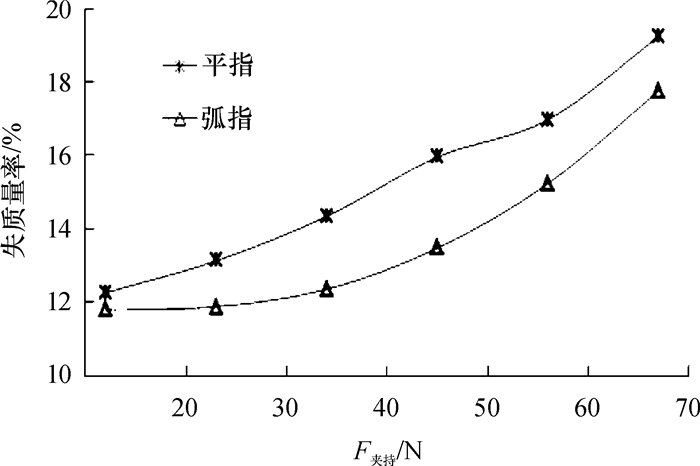
验证试验得到2种指型、不同夹持力下柑橘失质量率随时间变化(图 5)。由图 5可知,相同夹持力下,平指夹持下的柑橘失质量率普遍高于弧指夹持的,这与损伤预测分析相对应,因平指夹持的柑橘较早出现损伤,且果肉损伤率高于弧指夹持,因此柑橘容易失水导致失质量率增加。
![]() 图 5 不同夹持力下柑橘失质量率Figure 5. Weight loss rate of citrus fruit under different clamping force
图 5 不同夹持力下柑橘失质量率Figure 5. Weight loss rate of citrus fruit under different clamping force由图 5还可知,随着夹持力的增加,柑橘失质量率非线性增加,且不同指面夹持失质量率增加的趋势也不同,这也与损伤预测一致。当弧指夹持力小于45 N时,柑橘的失质量率差异较小,原因是此时柑橘末出现机械损伤;当夹持力大于45 N后,柑橘的失质量率上升较快,因其果肉已出现损伤,随着夹持力增大至67 N,果肉损伤率从20%增加至80%,导致柑橘水分流失加剧。平指夹持力大于45 N后,预测柑橘果肉损伤率增加至60%,且此时果皮也出现损伤,因此柑橘失质量率增加较多。
3. 结论
1) 通过压缩与拉伸试验,得到柑橘品种贡柑的纵向果皮和果肉的弹性模量分别为11.408和0.277 MPa,极限应力分别为1.250和0.048 MPa。
2) 柑橘夹持有限元模拟分析表明:夹持过程中柑橘果皮应力大于果肉,果肉先于果皮出现损伤,且损伤首先出现在夹持位置的两侧;弧指夹持下柑橘果皮和果肉的应力和变形量均小于平指夹持的,柑橘出现损伤的概率低于平指夹持;相同夹持力下,平指夹持的果肉损伤率高于弧指夹持。
3) 柑橘实物夹持试验结果表明,相同夹持力下,平指夹持的柑橘失质量率高于弧指夹持;无论是平指还是弧指夹持,柑橘失质量率增加的程度和趋势都与有限元对柑橘夹持损伤预测的结果相对应,这表明有限元法可以实现柑橘夹持损伤的模拟预测。
-
![]()
图 3 手指与柑橘夹持实体模型
Figure 3. The solid models of citrus fruit clampied by different fingers
![]()
图 4 平指夹持下柑橘的等效应力图
Figure 4. Equivalent stress distribution diagrams of citrus fruit clamped by plane fingers
![]()
图 5 不同夹持力下柑橘失质量率
Figure 5. Weight loss rate of citrus fruit under different clamping force
表 1 柑橘夹持有限元模拟试验等效应力
Table 1 The equivalent stress of finite element simulation of citrus fruit clamping
 下载: 导出CSV
下载: 导出CSV
表 2 不同夹持力下柑橘果肉损伤率预测
Table 2 Prediction in damage rate of citrus pulp under different clamping force
下载: 导出CSV
-
[1] LU L X. Dropping bruise fragilities and bruise boundaries of 'Gala' apples[J]. T CSAE, 2008, 24(4): 37-43. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=nygcxb200804007
[2] LIEN C C, AY C Y, TING C H. Non-destructive impact test for assessment of tomato maturity[J]. J Food Eng, 2009, 91(3): 402-407. doi: 10.1016/j.jfoodeng.2008.09.036
[3] ORTIZ C, BLASCO J, BALASCH S, et al. Shock absorbing surfaces for collecting fruit during the mechanical harvesting of citrus[J]. Biosyst Eng, 2011, 110(1): 2-9. doi: 10.1016/j.biosystemseng.2011.05.006
[4] 杨晓清, 王春光.河套蜜瓜机械特性与静载损伤关系的研究[J].农业工程学报, 2008, 24(3): 31-37. doi: 10.3321/j.issn:1002-6819.2008.03.007 [5] 吴杰, 郭康权, 葛云, 等.香梨果实跌落碰撞时的接触应力分布特性[J].农业工程学报, 2012, 28(1): 250-254. doi: 10.3969/j.issn.1002-6819.2012.01.044 [6] 陈燕, 向和平, 谈建豪, 等.挤压对荔枝机械损伤和力学参数的影响[J].湖南农业大学学报(自然科学版), 2013, 39(6): 688-692. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=SciencePaper201404040000001626 [7] KIM G W, DO G S, BAE Y, et al. Analysis of mechanical properties of whole apple using finite element method based on three dimensional real geometry[J]. Food Sci Technol Res, 2008, 14(4): 329-336. doi: 10.3136/fstr.14.329
[8] SADRNIA H, RAJABIPOUR A, JAFARI A, et al. Internal bruising prediction in watermelon compression using nonlinear models[J]. J Food Eng, 2008, 86(2): 272-280. doi: 10.1016/j.jfoodeng.2007.10.007
[9] 王芳, 王春光, 杨晓清.西瓜的力学特性及其有限元分析[J].农业工程学报, 2008, 24(11):118-122. doi: 10.3321/j.issn:1002-6819.2008.11.022 [10] 桑永英, 张东兴, 张梅梅.马铃薯碰撞损伤试验研究及有限元分析[J].中国农业大学学报, 2008, 13(1): 81-84. doi: 10.3321/j.issn:1007-4333.2008.01.017 [11] 陈燕, 蔡伟亮, 邹湘军, 等.荔枝的力学特性测试及其有限元分析[J].农业工程学报, 2011, 27(12): 358-363. doi: 10.3969/j.issn.1002-6819.2011.12.067 [12] 卿艳梅, 李长友, 黄汉东, 等.龙眼力学特性的有限元分析[J].农业机械学报, 2011, 42(6): 143-147. http://d.old.wanfangdata.com.cn/Periodical/nyjxxb201106029 [13] 吴杰, 李凡, 葛云, 等.香梨静压接触应力测量及损伤预测[J].农业工程学报, 2013, 29(6): 261-266. http://d.old.wanfangdata.com.cn/Periodical/nygcxb201306034 [14] SINGH K K, SREENIVASULA R B. Post-harvest physic-mechanical properties of orange peel and fruit[J]. J Food Eng, 2006, 73(2): 112-120. doi: 10.1016/j.jfoodeng.2005.01.010
[15] 姜松, 鲍黄贵, 蔡健荣, 等.不同成熟度柑橘力学特性研究[J].食品工业科技, 2007, 30(12): 117-119. http://d.old.wanfangdata.com.cn/Periodical/spgykj200912037 [16] 张水波, 鲍官军, 杨庆华, 等.基于机器人采摘的柑橘机械特性研究[J].浙江工业大学学报, 2012, 40(3): 340-344. doi: 10.3969/j.issn.1006-4303.2012.03.024 [17] 周祖锷.农业物料学[M].北京:农业出版社, 1994:49. [18] 瓦伦丁L. 波波夫, 李强, 等, 译. 接触力学与摩擦学的原理及其应用[M]. 北京: 清华大学出版社, 2011: 14. [19] GYASI S, FRIDLEY R B, CHEN P. Elastic and viscoelastic poisson's ratio determination for selected citrus fruits[J]. T ASAE, 1981, 24(3): 747-750. doi: 10.13031/2013.34332

计量
- 文章访问数: 1535
- HTML全文浏览量: 3
- PDF下载量: 1489
