Physiological response of Phoebe bournei seedlings with different phosphorus supply levels to natural low temperature
-
摘要:目的
探明自然低温对不同磷素供给的闽楠Phoebe bournei苗木叶片生理特性的影响。
方法设置8个磷素水平的单因素盆栽试验,施肥结束后,将苗木置于自然低温环境处理,测定其叶片的光合生理特性及其他生理指标,采用主成分分析法评价苗木抗寒性。
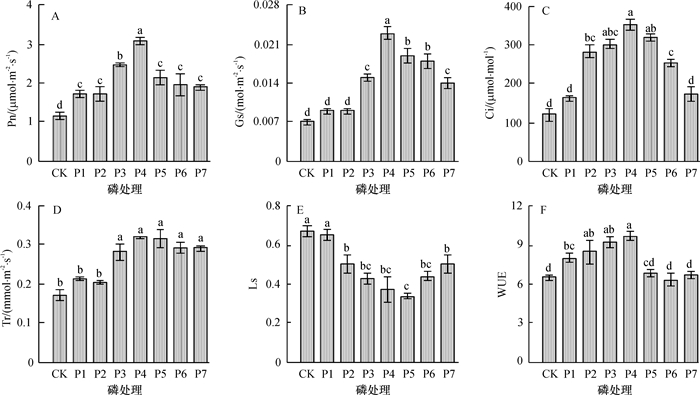
结果在自然低温环境时,随磷素水平的升高,正负相关指标均表现出了明显的升降趋势。30 mg·株-1磷素供应量的闽楠苗木叶片净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)、光合水分利用效率(WUE)、脯氨酸含量和超氧化物歧化酶(SOD)活性均最高,其值分别为3.09 μmol·m-2·s-1、0.023 mol·m-2·s-1、350.18 μmol·mol-1、0.32 mmol·m-2·s-1、9.69、41.58 μmol·g-1和332.03 U·g-1,丙二醛含量和气孔限制值(Ls)为第2低,其值分别为3.77 μg·g-1和0.34。主成分综合分析结果显示,30 mg·株-1磷素供应量苗木得分最高。
结论30 mg·株-1磷素供给最有利于闽楠苗木抗低温胁迫能力的提高。
Abstract:ObjectiveTo determine the effects of different phosphorus supply levels on physiological activity of Phoebe bournei seedling leaf under natural low temperature.
MethodSingle factor pot experiment was set up with eight phosphorus levels, and following fertilization the seedlings were left under natural low temperature. Photosynthetic and other physiological indexes of the leaves were measured and analyzed. Cold resistance of the seedlings was assessed by principal component analysis.
ResultUnder natural low temperature environment, all the positive and negative indexes indicated clear and gradual trend with the increase of phosphorus levels. net photosynthetic rate(Pn), stomatal conductance(Gs), intercellular CO2 concentration(Ci), transpiration rate(Tr), water use efficiency(WUE), proline content and SOD activity of P. bournei seedling leaf peaked at a phosphorus level of 30 mg phosphorus per seedling, and their values were 3.09 μmol·m-2·s-1, 0.023 mol·m-2·s-1, 350.18 μmol·mol-1, 0.32 mmol·m-2·s-1, 9.69, 41.58 μmol·g-1 and 332.03 U·g-1, respectively. At the same phosphorus level, MDA content (3.77 μg·g-1) and stomatal limitation value (Ls, 0.34) were the second lowest. The highest cold resistance score was obtained at a phosphorus level of 30 mg per seedling based on principal component analysis.
ConclusionThe optimum phosphorus level is 30 mg per seedling for improving cold resistance of P. bournei seedlings.
-
芳香族化合物是一类具有芳香性质的有机小分子,包含1个或多个芳香环结构。这些分子在食品、化妆品、医药等领域具有重要的作用。芳香族化合物可以从植物、微生物和化学合成等多种来源获得,但由于其种类繁多、结构复杂、生物活性差异大以及受环境影响和资源限制等因素,传统的生产和利用方式难以满足人们对芳香族化合物的需求。因此,合成生物学作为一种新兴的跨学科领域,为芳香族化合物的研究和应用提供了新的思路和方法,也是生物育种中的一条有效途径。合成生物学在芳香族化合物方面的应用主要包括2个方面:一是利用微生物或植物作为代谢工程平台,对芳香族化合物的生物合成途径进行优化或重构,提高芳香族化合物的产量和质量;二是在植物中对芳香族化合物的生物合成进行优化和改造,通过合成生物学技术,可以对植物中芳香族化合物进行定向调控和改良,从而提高作物的抗逆性、抗病性,改善营养价值和食味品质等,实现其在植物育种中的利用。本文介绍了芳香族化合物及合成生物学,重点综述芳香族化合物在生物育种中的研究进展。
1. 芳香族化合物及生物合成
1.1 芳香族化合物的来源及用途
芳香族化合物是一类化学性质稳定且具有芳香性质的有机分子,通常含有一个或多个芳香环结构。这些分子的共轭π电子体系使其具有稳定性和芳香性质[1]。芳香族化合物的种类非常多,包括苯、萘、吡啶、吡咯和噻吩及其衍生物等(图1)。其中,苯是最简单的芳香化合物,苯环上每个碳原子都与一个氢原子相连形成一个六元环结构。苯环的π电子构成一个连续的电子体系,使其具有很强的稳定性[2]。在某些情况下,含有苯环或其衍生物结构的化合物也能被归类为芳香族化合物。除苯环外,还有许多其他类型的芳香环,它们的共轭π电子体系和化学性质与苯环有所不同,但它们都具有芳香性质。萘是由2个苯环融合而成的多环芳香化合物,比苯更活泼,更容易发生各种反应[3]。吡啶则是一种含有1个六元杂环的芳香化合物,由于含有1个氮原子,使其具有许多特殊的物理和化学性质[4]。芳香族化合物是有机化学研究中的重要组成部分,其多样的结构和广泛的应用使其在化学、材料和生命科学等领域具有重要的地位。
芳香族化合物可以从多种来源获得,包括植物、微生物和化学合成等。植物中的许多挥发性油和香料都富含芳香族化合物,如薄荷Mentha haplocalyx Briq.中的薄荷醇和茉莉花Jasminum sambac (L.) Ait中的苯乙醇等[5-6]。微生物也是一种重要的芳香族化合物的生产来源,例如,青霉菌Penicillium和链霉菌Streptomyces sp.等微生物可以合成多种芳香族化合物[7]。此外,化学合成也是获得芳香族化合物的另一种重要途径,包括从原料中提取或通过化学反应合成等多种方式[3]。
芳香族化合物在食品、化妆品、医药等许多领域都有广泛的应用。在食品工业中,芳香族化合物被广泛用作食品添加剂,如香料和香精等,以赋予食品独特的香味和口感[8-9];一些芳香族化合物还具有抗菌、抗氧化和消炎等生物活性,在食品中可以使用这些芳香族化合物作为天然防腐剂来保证食品的新鲜度和品质,同时还能增加食品的营养和保健功能[10-11]。在化妆品领域,许多芳香族化合物被广泛用于制造香水、洗发水、护肤品等产品[12];芳香族化合物还用于制作芳香疗法精油,广泛应用于舒缓压力、缓解焦虑等方面[13]。在医药领域,芳香族化合物被广泛用于治疗各种疾病[14]。
1.2 合成生物学及应用
合成生物学是一种利用基因工程、代谢工程和人工合成细胞等技术,设计和构建具有新功能的生物系统学科[15]。它致力于设计和构建新的生物系统和生物器件,以实现有用的功能。合成生物学通过将生物学、工程学、计算机科学和物理学等多学科的知识和方法融合,开发出更加可控、可重复和可预测的生物系统,为解决生物技术、能源、环境等领域存在的问题提供新的解决方案和新的工具。合成生物学的研究内容包括合成生物系统的设计原理、生物元件的开发策略、合成基因组的构建方法、代谢途径的优化技术等[16]。基因工程是合成生物学中最为基础和核心的技术之一,它可以通过人工合成或改造生物体的基因组来控制生物系统的行为[17];代谢工程则是通过基因调控和代谢途径的改造来构建新的代谢网络,从而实现对代谢产物的高效生产[18]。在合成生物学的应用中,可以利用基因工程和代谢工程技术构建具有特定功能的微生物或植物,例如合成新的代谢产物、制备高效的酶催化体系等。合成生物学在医药、环境保护、能源和食品等领域都具有广阔的应用前景[19-20]。
合成生物学为芳香族化合物的生产提供了新思路和新途径[21]。传统的化学合成芳香族化合物的方法不仅需要消耗大量的能源和化学试剂,还会产生大量的污染物,对环境造成极大的破坏。采用合成生物学的方法,利用微生物或植物生产芳香族化合物,不仅可以降低生产成本,还可以减少对环境的负担,对于解决工业生产中的环境和经济问题具有重要意义[22]。此外,利用合成生物学的方法,可以对芳香族化合物的生物合成途径进行精准的调控和优化,从而提高生产效率和产物品质,拓展产物种类。例如,通过基因工程和代谢工程技术,可以改造菌株或植物的代谢途径,增加芳香族化合物的产量和品质;还可以构建新的代谢途径,实现对新型或稀有芳香族化合物的生产。
1.3 芳香族化合物的主要生物合成途径——莽草酸途径
莽草酸途径又称分支酸途径,是生物合成芳香族氨基酸和其他芳香族化合物的重要途径[23]。莽草酸途径存在于细菌、真菌和植物等多种生物中,但在动物中却不存在。所以,对于动物而言,该途径中的一些代谢产物是必需的营养素[24-25]。莽草酸合成的2种初始底物是磷酸烯醇式丙酮酸(PEP)和赤藓糖−4−磷酸(E4P),PEP来自糖酵解与糖异生途径,E4P则来自卡尔文循环与磷酸戊糖途径。1个E4P分子和2个PEP分子首先在DAHP合成酶的催化下合成为3−脱氧−D−阿拉伯庚糖酮酸−7−磷酸(DAHP),并释放1个磷酸;DAHP再在DHQS的催化下环化为3−脱氢奎尼酸(DHQ);DHQ在DHD的催化下脱水形成3−脱氢莽草酸(DHS);之后,在SDH的催化下,DHS消耗掉1个NADPH还原为莽草酸(SHK)[26]。 接下来,SHK在莽草酸激酶的作用下消耗ATP磷酸化为莽草酸−3−磷酸(S3P);S3P再在EPSP合成酶的催化下与PEP结合形成5−烯醇式丙酮酸莽草酸−3−磷酸(EPSP)。而EPSP则在分支酸合成酶的催化下去掉1个磷酸基团生成分支酸(CHR)(图2)。从分支酸开始就可以合成芳香族氨基酸AAAs(L−络氨酸、L−苯丙氨酸、L−色氨酸),也可以通过其他支路合成多种有机化合物,如生育酚、维生素B9、木质素、异黄酮、香豆素、香草醛、苯甲酸衍生物等[27-28]。
![图 2 植物中莽草酸和芳香族氨基酸代谢网络示意简图]() 图 2 植物中莽草酸和芳香族氨基酸代谢网络示意简图Figure 2. The schematic diagram of shikimate and aromatic amino acid metabolic networks in plants
图 2 植物中莽草酸和芳香族氨基酸代谢网络示意简图Figure 2. The schematic diagram of shikimate and aromatic amino acid metabolic networks in plants在大多数细菌中,莽草酸途径的主要功能是为蛋白质合成提供芳香族氨基酸。这一途径在细菌中得到了广泛的探索,部分原因是该途径在食品和医药工业中具有重要应用价值,例如,可以利用该途径生产食品添加剂香豆素、流感药物“达菲”和止痛药“吗啡”等[29]。
在植物中,阐明莽草酸途径的代谢和调控机制也至关重要。据估计,来源于莽草酸途径的木质素是植物细胞壁中含量最高的生物聚合物之一,约占生物圈有机碳的30%[30]。这一途径在植物发育以及与环境的相互作用中也发挥着广泛的调节作用,因此,植物必须微调莽草酸途径的活性,以控制丰富多样的芳香化合物的产生,并协调复杂的发育和生理过程[31]。
DAHPS是催化莽草酸途径第一步反应中控制植物碳代谢流向芳香族天然产物生物合成的关键酶。DAHPS可分为I型和II型2种形式,I型和II型DAHPS序列相似性较低但具有相似的三维结构,大多数微生物具有I型酶,而II型DAHPS则存在于光合生物及少数的真菌和细菌中。陆生植物中通常存在多种DAHPS亚型,这与植物可产生多种芳香族化合物的进化方向相一致[28,32]。在细菌中,DAHPS受到芳香族氨基酸产物的反馈抑制[33-34]。不同生物的DAHPS反馈抑制机制有所不同,在拟南芥Arabidopsis thaliana (L.) Heynh中,DAHPS受到下游产物酪氨酸、色氨酸、芳基丙氨酸、咖啡酸和分支酸等的反馈抑制[35]。因此,通过突变DAHPS与抑制剂的结合位点可以解除这种反馈抑制,增加DAHPS的活性。例如,在大肠埃希氏菌Escherichia coli中,将AroG第175位的亮氨酸(Leu)突变为谷氨酸(Gln),可以有效地解除Phe的抑制[36];在拟南芥中,通过对tyra2突变体进行抑制和突变体筛选,找到了DAHPS的多个突变位点,这些突变能有效地解除DAHPS的反馈抑制,显著增加植物中芳香族氨基酸的含量[37]。
2. 芳香族化合物在生物育种中的应用
植物需要芳香族化合物来参与其生长、发育和应激反应,这些化合物是由莽草酸途径产生的。莽草酸途径是一种主要的代谢途径,它可以合成芳香族氨基酸(色氨酸、苯丙氨酸和酪氨酸)和各种次级代谢物(木质素、黄酮类和芳香化合物)[38]。近年来,随着人们对植物中芳香族化合物代谢途径、反馈回路、功能基因等方面的深入了解,以及生物信息挖掘分析、目标酶的定向进化、基因编辑等生物技术的利用,使合成生物学成为了一个有前景的领域,可以用来改造植物中的芳香族化合物[39-41]。与微生物相比,植物作为内源或异源代谢途径的宿主具有一些优势,例如,植物可以自给自足地进行代谢过程,不需要额外的培养基或设施,而且更容易扩大规模,从而降低成本[42];此外,植物的花朵和果实往往是人们食用的部分,因此改善它们的芳香族化合物含量可以提高食品的风味和营养品质,这也是生物育种中的一个重要目标[43-46]。
2.1 富香味物质生物育种
植物体内存在着许多芳香族化合物,它们是由苯丙氨酸转化而来的。这些化合物散发出各种令人愉悦的芳香,例如丁香酚(丁香的香气)、覆盆子酮(覆盆子的香气)和香草素(香草的香气)等,这些化合物不仅对植物本身,还对动物和人类都有重要的生理功能。科学家对植物中这些化合物的生物合成机制进行了广泛深入的研究,揭示了参与生成和释放这些化合物的基因和转录因子的作用 ,为利用生物技术提高这些芳香族化合物的含量提供了重要的基因资源[47-50]。辣椒素是香草素的前体,通过在辣椒Capsicum frutescens愈伤组织中沉默掉催化香草素转化的酶或者使香草醛合酶基因(VpVAN)异位表达,就可以显著增加香草素及其葡萄糖苷的含量[51-52];VpVAN也可以在烟草Nicotiana tabacum和大麦Hordeum vulgare中表达,使植物产生香草醇葡萄糖苷[53]。另一种方法是利用假单胞菌中的4−羟基肉桂酰辅酶A水合酶/裂解酶(HCHL),将木质素前体阿魏酰辅酶A转化为香草素,这种方法已经在甜菜Beta vulgaris毛状根培养物中成功应用[54-55]。HCHL也可以在烟草中异位表达,从而提高香草素衍生物的含量,但是会导致植物出现多种表型异常[56-57]。矮牵牛花Pharbifis nil是一种富含香草素的植物,如果在矮牵牛花中过表达苯丙烷途径的转录因子PAP1或者使反馈抑制不敏感的AroG和PheA酶异位表达,均可以进一步增加香草素及其衍生物的水平[58-60]。为了提高植物中芳香族化合物的含量,单个基因的操控往往不够,需要多个基因的协同作用,以优化复杂的生物合成途径。例如,草莓 Fragaria × ananassa果实中单独表达矮牵牛异丁香酚合酶(IGS)并不能有效地产生异丁香酚,但当同时抑制类黄酮生物合成途径的查尔酮合酶(CHS)就可以显著增加异丁香酚的含量 [44, 59, 61]。类似地,在烟草中同时表达BAS和RZS1可以使覆盆子酮及糖苷积累;如果再抑制CHS,就可以使更多的底物香豆酰辅酶A(p-CA)流向BAS作用的合成途径,从而大大提高覆盆子酮的含量[62]。除了抑制支流外,还可以通过表达转录因子来增强合成途径的基因表达。例如,在烟草中过表达BAS和RZS1并抑制CHS的基础上,再过表达拟南芥转录因子PAP1,就可以使烟草叶片中的覆盆子酮含量增加3倍,达到与覆盆子果实中相近的含量水平[62]。转录因子PAP1可以正向调控苯丙烷途径;此外,还有其他许多转录因子可以调控苯丙烷途径和挥发性苯类化合物的合成,其中,比较有名的是MYB-ODO1-EOB三联体,主要在矮牵牛中发挥作用,在其他植物中也有同源物 。ODO1可以激活苯丙氨酸解氨酶(PAL)以及其他苯丙烷途径相关基因的表达,从而增加苯丙氨酸的可利用性;EOB I和II可以激活ODO1和一些挥发性苯类化合物合成相关基因的表达,从而增加花香的产生[50, 63-64]。另外,GRAS家族成员Phenylpropanoid emission-regulating scarecrow-like (PES)可以在花瓣中同时激活ODORANT1、苯丙烷及槲皮酸途径酶的基因表达,使矮牵牛的花产生大量挥发性香味物质[65]。尽管多基因组合具有潜力,但同时过表达他们不一定能得到我们想要的目标,还可能受到产物的抑制、代谢网路及转录因子的调控[66-67]。因此,研究不同转基因之间的相互作用及其对代谢流的影响对阐释不同基因及其组合的效果十分必要。
2.2 抗除草剂生物育种
草甘膦是一种广泛应用的除草剂,它的作用机制是抑制莽草酸途径中的关键酶5−烯醇丙酮酰亚胺−3−磷酸合成酶(EPSPS)。为了让农作物在使用草甘膦除草时不受影响,一种方法是将具有草甘膦抗性的EPSPS基因转入农作物中,使其能够正常进行莽草酸途径的代谢[68]。例如,将带有叶绿体定位信号的CP4-EPSPS基因(一种草甘膦抗性变体)过表达在大豆Glycine max、玉米Zea mays、水稻Oryza sativa和棉花Gossypium herbaceum等作物中[69-71],可以培育出抗草甘膦的转基因作物,有效地抑制杂草的生长,同时不损害作物的生长和产量,这些作物已经实现了商业化种植[72]。另一种方法是利用基因编辑技术,将植物自身的EPSPS基因编辑为草甘膦抗性的突变体,这样既能保证材料的稳定性,又能避免转基因成分的问题。例如,Endo 等[73]利用定向突变技术,将水稻 EPSPS 第102位的苏氨酸(T)替换成异亮氨酸(I)(T102I),创造出抗草甘膦的水稻。Li 等[74]采用引导编辑技术,将水稻 EPSPS 169位的苏氨酸(T)、第170位的丙氨酸(A)和第173位的脯氨酸(P)分别替换为异亮氨酸(I)、缬氨酸(V)和丝氨酸(S)(T169I、A170V和P173S) ,得到了高度抗草甘膦的水稻。这些研究表明,基因编辑技术在提高农作物对草甘膦耐受性方面的高效性和巨大的潜力。
2.3 富含类黄酮类生物育种
黄酮类化合物是一类广泛存在于苔藓植物到被子植物中的酚类化合物,主要以二氢黄酮、黄酮、黄酮醇、异黄酮等形式存在,它们在植物抵抗生物和非生物胁迫中起重要作用,同时也是一类具有很强生物活性的化合物,是天然的抗氧化剂,能降低心肌耗氧量、防治血管硬化等,具有抗衰老、增强机体免疫力、抗癌防癌的功效,在医药、食品等领域有广阔的应用前景[75-76]。利用转基因技术,可以通过过表达或抑制类黄酮次生代谢的关键酶基因,改变类黄酮次生代谢途径,从而提高或减少类黄酮物质的含量。例如,查尔酮合成酶(CHS)是黄酮生物合成途径中的关键酶和第一个速率限制性酶[77-78]。在番茄Solanum lycopersicum中,通过RNA 干扰(RNAi)介导抑制CHS可以降低总黄酮水平[79]。查尔酮还原酶(CHR)是一种醛酮还原酶超家族成员,作用于 CHS 反应的中间体,催化C−6′ 去羟基化,生成异甘草素(4,2′,4′−三羟基查尔酮)[80]。在矮牵牛中过表达来自日本莲Lotus japonicus的 CHR1 基因,导致异甘草素的形成和花青素含量的降低[81]。 查尔酮异构酶(CHI)是黄酮生物合成途径中的第2个关键限速酶 [82]。在龙血树Dracaena draco和烟草中,DcCHI1或DcCHI4的过表达导致类黄酮积累增加 [83]。黄酮是由黄烷酮类在黄酮合成酶(FNS)的作用下生成的。在拟南芥中过表达黄丝瓜藓Pohlia nutans 的FNSI增强了拟南芥中的黄酮生物合成途径,使植株抵抗干旱胁迫和UV−B辐射的能力加强[84]。
然而,植物类黄酮生物合成途径非常复杂,涉及到多种类型的酶[85-86],有时单个关键酶并不能完全控制类黄酮的合成。通过转录因子作用,可能激活类黄酮次生代谢途径中多个功能基因的表达,从而达到更明显的效果。包含MYB、bHLH和WD40转录因子家族的MBW(MYB-bHLH-WDR)复合体是调控类黄酮合成的主要因子[87]。在石斛兰Dendrobium nobile中过表达DhMYB2或DhbHLH1能够使白色花瓣合成花青素而改变颜色[88]。在番茄果肉中表达玉米MYB和bHLH转录因子C1和Lc基因,可以显著提高黄酮醇相关酶基因的表达,黄酮醇含量约为对照的20倍[89]。而另一个名为SlAN11的WD40转录因子能够显著影响番茄中类黄酮的合成和种子休眠。过表达SlAN11的番茄植株中花青素和原花青素的含量明显增加,而黄酮醇的含量则降低;反之,抑制SlAN11的番茄植株,类黄酮的合成则受到抑制[90]。SlAN11的激活机制是通过与bHLH转录因子直接互作,以及通过bHLH转录因子与MYB转录因子间接互作而形成MBW复合体[90]。MBW复合体可以激活类黄酮合成途径中的后期基因,如SlDFR等[90]。除MBW复合体外,其他的转录因子也能够调控类黄酮代谢途径,如抑制NF-YB基因的表达可以降低番茄果实成熟过程中CHS1的表达水平,使得果实发育成粉红色果肉和无色果皮[91],为培育不同果色的番茄提供了理论基础。
2.4 低木质素生物育种
木材是由次生增厚细胞壁构成的木质纤维素生物质,是一种重要的资源。次生细胞壁主要含有纤维素、半纤维素和木质素,均是由不同单元组成的大分子化合物;纤维素和半纤维素是由单糖连接而成的多糖;木质素是由对香豆醇、松柏醇和芥子醇这3种芳香族单体聚合而成的一种复杂的酚类物质[92]。纤维素是制浆造纸工业的重要原料,纤维素和半纤维素都可以被转化为单糖,用于生产各种发酵产品,例如生物乙醇和乳酸[93]。一方面,木质素对以上加工有不利影响,因此,可以通过基因工程使树木积累更少的木质素,从而提高生产纸张和可发酵糖的效率;另一方面,木质素也是一种有价值的芳香源,它可以被分解为芳香族化合物,但由于分解难度大,其利用还处于初级阶段[94]。关于木质素的生物育种目前主要是为了降低其含量,从而改善木材加工性能[95](图3)。
![图 3 芳香族化合物在生物育种中的应用]() 图 3 芳香族化合物在生物育种中的应用a:富香味物质生物育种;b:抗除草剂生物育种;c:富含类黄酮类生物育种;d:低木质素生物育种Figure 3. Application of aromatic compounds in biological breedinga: Breeding for flavour-rich substances; b: Breeding for herbicide resistance; c: Flavonoid-rich bio-breeding ; d: Low lignin bio-breeding
图 3 芳香族化合物在生物育种中的应用a:富香味物质生物育种;b:抗除草剂生物育种;c:富含类黄酮类生物育种;d:低木质素生物育种Figure 3. Application of aromatic compounds in biological breedinga: Breeding for flavour-rich substances; b: Breeding for herbicide resistance; c: Flavonoid-rich bio-breeding ; d: Low lignin bio-breeding木质素的生物合成途径包括从苯丙氨酸到单酚醇多个步骤,理论上任何一个步骤的活性降低都可能导致木质素含量的降低,不同步骤的活性下调对木质素含量的影响程度不同,而且还受到环境因素的制约。一般来说,从苯丙氨酸解氨酶(PAL)到肉桂酰辅酶A还原酶(CCR)步骤的下调可以更有效地降低木质素含量[95-96]。一些下调了这些基因(如4CL、CAD1)的杨树Populus L.在温室中木质素含量明显降低,但在田间却没有明显变化;这些杨树在温室中生长正常,但在田间却出现了生长抑制[97-100]。为了避免木质素含量降低对植株生长的不利影响,需要控制目标基因的调控强度和空间表达位置。例如,在松树Pinus species中利用RNAi技术下调CSE基因可以使木质素含量降低25%,且不影响生长[101];但利用CRISPR技术敲除CSE基因则会使木质素含量降低35%,同时导致生长受阻[102]。在杨树中利用CRISPR技术对CCR2基因进行弱突变可以使木质素含量降低10%且不出现敲除后的生长异常[103]。此外,在杨树中可以通过特异性抑制纤维细胞中木质素生物合成以实现纤维低木质素化,例如,通过特异性下调4CL1或特异性表达抑制子LTF1AA可以使转基因杨树具有低木质素化的纤维和正常的导管及植株生长;相反,如果在导管中抑制木质素生物合成,则会导致转基因杨树出现导管塌陷和严重的生长抑制[104-105]。这些研究表明,要获得低木质素化的木材,保证导管中有足够的木质素是避免抑制植株生长的关键。
木质素生物合成途径复杂且受环境影响大,因此利用AI模拟调控策略,有助于实现低木质素生物育种的目标。SULIS等[106]采用机器学习模型,寻找最优的多基因编辑策略,对21个木质素生物合成基因的69 123种可能组合进行了评估,得出了7种可以同时改变6个基因的策略,利用这些策略生成了174种编辑过的杨树突变体,结果表明,部分突变体木材中的碳水化合物与木质素的比例高达野生型的228%,显著提高了纤维制浆效率;这些突变体不仅解决了纤维生产的主要瓶颈问题,还带来运营效率以及生物经济和环境效益方面的巨大优势。
3. 展望
合成生物学是一门不断发展的学科,它可以广泛应用于芳香族化合物的研究。随着新工具和技术的不断涌现,利用合成生物学手段探索和操纵这一领域的可能性将持续增加,可以预见,合成生物学在芳香族化合物研究中将具有巨大的潜力。
3.1 开发具有改良特性的新型植物产品
通过改变植物的代谢途径,可以提高有价值化合物的产量,或者创造出具有理想特性的新化合物,用于医药、化妆品或其他行业。此外,合成生物学可以帮助研究芳香族化合物在生物体内的作用机制,利用合成生物学技术构建改变代谢途径的合成生物体以产生特定的芳香族化合物,或者研究不同的芳香族化合物如何与生命系统中的其他分子相互作用。在使用合成生物学研究芳香族化合物时也面临着一些挑战和限制,如创建和操纵复杂的代谢途径可能是一个困难和耗时的过程;可能会对生物体的整体代谢产生不可预见的影响;此外,还需要妥善处理人们对转基因生物可能带来的环境和健康风险的担忧。尽管存在这些困难,但合成生物学在芳香族化合物研究中所能带来的好处使其成为未来值得关注的研究领域。通过开发新的工具和技术,研究人员可以充分发挥芳香族化合物的潜能,创造出新的应用和产品。
3.2 精确操纵和修改植物中与芳香化合物生产相关的基因
利用新的高效基因编辑工具,如CRISPR-Cas9系统,精确操纵和修改植物中与芳香族化合物生产相关的基因,可以使基因修饰更加高效和具有针对性,减少对传统育种方法的依赖。此外,人工智能的使用,使得高通量筛选和数据分析更加容易,促使研究人员能够从大量的候选基因中筛选出对植物代谢(包括芳香族化合物的生产)有潜在影响的基因,并预测其作用方式,有助于更有效地确定目标基因和最佳的修改策略。合成生物学还可以用于生产新型化合物和现有芳香族化合物的衍生物,这将促进具有改良特性和商业价值的新产品开发。Stepanyuk等[107]利用合成生物学技术生产出具有独特和理想香味的新型香味化合物,而这些香味在自然界中是不存在的。
3.3 增加育种资源,提高育种效率
合成生物学在生物育种方面展现出巨大的潜力,尤其是在利用莽草酸途径方面,可以提高育种效率,增加育种资源,并适应市场需求。然而,也存在一些需要克服的挑战和限制,包括安全评估、社会认可和法律监管等,这些问题的解决对于成功推广基于合成生物学的生物育种并实现其潜在效益至关重要。合成生物学可能对植物中芳香化合物的研究和操纵产生革命性的影响,促进植物育种、作物生产和新型产品的开发取得重大突破;随着基因编辑工具和技术的不断发展和改进,合成生物学将在未来植物科学和农业发展中发挥重要作用。
-
![]()
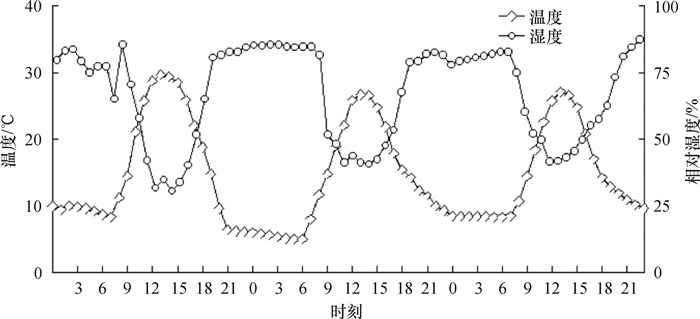
图 1 1月10—12日测定期内自然环境的温、湿度变化
Figure 1. Changes of temperature and moisture in the natural environment from 10 to 12 January
![]()
图 2 不同磷水平处理对闽楠苗木叶片光合生理特性的影响
各图中,柱子上方凡具有一个相同小写英文字母者,表示差异不显著(P>0.05,Duncan’s法)。
Figure 2. Photosynthetic physiological characteristics of Phoebe bournei seedlings with different phosphorus treatments
表 1 不同磷素水平处理对苗木脯氨酸、丙二醛含量和SOD活性的影响1)
Table 1 Proline and MDA contents and SOD activity of Phoebe bournei seedlings with different phosphorus treatments

 下载: 导出CSV
下载: 导出CSV
表 2 不同磷素水平下闽楠苗木抗低温得分
Table 2 Cold resistance scores of Phoebe bournei seedlings with different phosphorus treatments
 下载: 导出CSV
下载: 导出CSV
-
[1] 郭淑红, 薛立, 张柔, 等. 4种幼苗对低温胁迫的光合生理响应[J].华南农业大学学报, 2012, 33(3): 373-377. doi: 10.3969/j.issn.1001-411X.2012.03.020 [2] 王建林, 于贵瑞, 房全孝, 等.不同植物叶片水分利用效率对光和CO2的响应与模拟[J].生态学报, 2008, 28(2):525-533. doi: 10.3321/j.issn:1000-0933.2008.02.010 [3] VARONE L, RIBAS-CARBOS M, CARDONA C, et al.Stomatal and non-stomatal limitations to photosynthesis in seedlings and saplings of Mediterranean species pre-conditioned and aged in nurseries: Different response to water stress[J]. Environ Exp Bot, 2012, 75(1):235-247.
[4] WANG D, HECKATHORN S A, WANG X Z, et al.A meta-analysis of plant physiological and growth responses to temperature and elevated CO2[J].Oecologia, 2012, 169(1):1-13. doi: 10.1007/s00442-011-2172-0
[5] PALLARDY S G.Physiology of woody plants[M].Missouri: Academic Press, 2010:225-226.
[6] 侯立刚, 陈温福, 马巍, 等.低温胁迫下不同磷营养对水稻叶片质膜透性及抗氧化酶活性的影响[J].华北农学报, 2012, 27(1):118-123. doi: 10.3969/j.issn.1000-7091.2012.01.022 [7] 侯伟, 孙爱花, 杨福孙, 等.低温胁迫对西瓜幼苗光合作用与叶绿素荧光特性的影响[J].广东农业科学, 2014, 41(13):35-39. doi: 10.3969/j.issn.1004-874X.2014.13.008 [8] TRAN H T, HURLEY B A, PLAXTON W C. Feeding hungry plants: The role of purple acid phosphata-ses in phosphate nutrition[J].Plant Sci, 2010, 179(1):14-27. https://www.sciencedirect.com/science/article/pii/S0168945210001007?_rdoc=3&_fmt=full&_origin=browse&_ct=14&md5=bbbea140ea15724c44af575f477a38e2
[9] GARRISH V, CERNUSAK L A, WINTER K, et al. Nitrogen to phosphorus ratio of plant biomass versus soil solution in a tropical pioneer tree, Ficus insipida[J].J Exp Bot, 2010, 61(13):3735-3748. doi: 10.1093/jxb/erq183
[10] 赵琼, 曾德慧.林木生长氮磷限制的诊断方法研究进展[J].生态学杂志, 2009, 28(1):122-128. http://d.old.wanfangdata.com.cn/Periodical/stxzz200901020 [11] 吴国欣, 王凌晖, 梁惠萍, 等.氮磷钾配比施肥对降香黄檀苗木生长及生理的影响[J].浙江农林大学学报, 2012, 29(2):296-300. doi: 10.3969/j.issn.2095-0756.2012.02.021 [12] 丁彦芬, 高俊飞, 张利.配方施肥对榉树幼苗生长的影响[J].南京林业大学学报(自然科学版), 2014, 38(S1):35-38. http://d.old.wanfangdata.com.cn/Periodical/njlydxxb2014z1008 [13] 王樱琳, 韦小丽, 段如雁, 等.闽楠幼苗对大量元素缺乏的响应[J].西北林学院学报, 2014, 29(2):61-65. doi: 10.3969/j.issn.1001-7461.2014.02.11 [14] 王艺, 王秀花, 吴小林, 等.缓释肥加载对浙江楠和闽楠容器苗生长和养分库构建的影响[J].林业科学, 2013, 49(12):57-63. http://d.old.wanfangdata.com.cn/Periodical/lykx201312009 [15] ZHOU S X, DUURSMA R A, MEDLYN B E, et al.How should we model plant responses to drought? An analysis of stomatal and non-stomatal responses to water stress[J].Agr Forest Meteorol, 2013, 183(22):204-214.
[16] HUANG Z, ZOU Z R, HE C X, et al.Physiological and photosynthetic responses of melon (Cucumis melo L.) seedlings to three Glomus species under water deficit[J].Plant Soil, 2011, 339(1):391-399. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=JJ0221679337
[17] 陈建勋, 王晓峰.植物生理学实验指导[M].广州:华南理工大学出版社, 2002:115-118. [18] 何跃君, 薛立, 任向荣, 等.低温胁迫对六种苗木生理特性的影响'[J].生态学杂志, 2008, 27(4):524-531. http://d.old.wanfangdata.com.cn/Periodical/stxzz200804006 [19] TAIZ L, ZEIGER E.Plant physiology[M]. Sinauer Associates:Sunderland, 2006:227-228.
[20] 陈琳, 曾杰, 贾宏炎, 等.林木苗期营养诊断与施肥研究进展[J].世界林业研究, 2012, 25(3):26-31. http://d.old.wanfangdata.com.cn/Periodical/sjlyyj201203005 [21] OGAYA R, PEUELAS J, ASENSIO D, et al.Chlorophyll fluorescence responses to temperature and water availability in two co-dominant Mediterranean shrub and tree species in a long-term field experiment simulating climate change[J]. Environ Exp Bot, 2011, 73(2):89-93.
[22] GULIAS J, CIFRE J, JONASSON S, et al.Seasonal and inter-annual variations of gas exchange in thirteen woody species along a climatic gradient in the Mediterranean island of Mallorca[J].Flora-Morphol, Distribut Funct Ecol Plant, 2009, 204(3):169-181. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=JJ0220297874
[23] 郑春芳, 刘伟成, 陈少波, 等.短期夜间低温胁迫对秋茄幼苗碳氮代谢及其相关酶活性的影响[J].生态学报, 2013, 33(21):6853-6862. http://d.old.wanfangdata.com.cn/Periodical/stxb201321011 [24] 张毅龙, 张卫强, 甘先华.低温胁迫对6种珍贵树种苗木光合荧光特性的影响[J].生态环境学报, 2014, 23(5):777-784. doi: 10.3969/j.issn.1674-5906.2014.05.008 [25] FLEXAS J, DIAZESPEJO A, GAGO J, et al.Photosynthetic limitations in Mediterranean plants:A review[J].Environ Exp Bot, 2014, 103(3):12-23. https://www.researchgate.net/publication/261565547_Photosynthetic_limitations_in_Mediterranean_plants_A_review
[26] 于金平, 俞珊, 梁有旺, 等.NaCl胁迫对美国白蜡幼苗部分生理指标的影响[J].植物资源与环境学报, 2014, 23(1):110-112. doi: 10.3969/j.issn.1674-7895.2014.01.18 [27] ANDIVIA E, MÁRQUEZ-GARCÍA B, VÁZQU-EZ-PIQUÉ J, et al.Autumn fertilization with nitrogen improves nutritional status, cold hardiness and the oxidative stress response of Holm oak (Quercus ilex ssp. ballota[Desf.] Samp) nursery seedlings[J]. Trees, 2012, 26(2):311-320. doi: 10.1007/s00468-011-0593-3
[28] 徐艳丽, 鲁剑巍, 周世力, 等.氮磷钾肥对高羊茅生长及抗寒性的影响[J].植物营养与肥料学报, 2007, 13(6):1173-1177. doi: 10.3321/j.issn:1008-505x.2007.06.030 [29] FAVARETTO V F, MARTINEZ C A, SORIANI H H, et al.Differential responses of antioxidant enzymes in pioneer and late-successional tropical tree species grown under sun and shade conditions[J].Environ Exp Bot, 2011, 70(1):20-28. doi: 10.1016/j.envexpbot.2010.06.003
[30] 王东光, 尹光天, 杨锦昌, 等.磷肥对闽楠苗木生长及叶片氮磷钾浓度的影响[J].南京林业大学学报(自然科学版), 2014, 38(3):40-44. http://d.old.wanfangdata.com.cn/Periodical/njlydxxb201403008 -
期刊类型引用(3)
1. 陈传卫. 大豆玉米间作对作物生长特性及产量的影响. 基层农技推广. 2025(04): 9-12 .  百度学术
百度学术
2. 吴玉雯,张应榕,吴晓芊,张俊,马悦,周乐儒,王娟红,车庆轩,陈波浪. 施磷方式对棉花根系形态和磷吸收效率的影响. 中国土壤与肥料. 2025(04): 151-161 . 百度学术
3. 胡闻君,陈颖,周园园,邬品中,龚奕杰,王珊,冯均科. 玉米大豆间作试验初步研究. 农业与技术. 2024(24): 33-36 . 百度学术
其他类型引用(1)