A nonlinear rheological model of castor pressing based on stress-relaxation tests
-
摘要:目的
揭示蓖麻籽压榨过程中的流变特征.
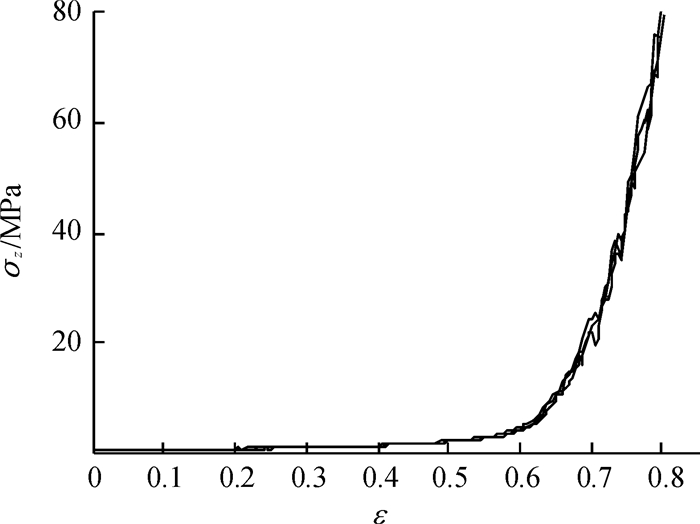
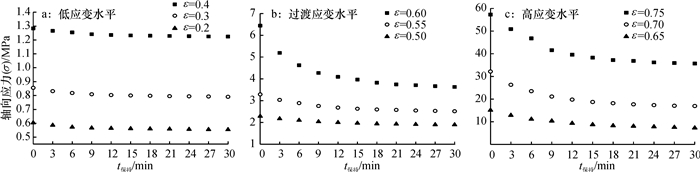
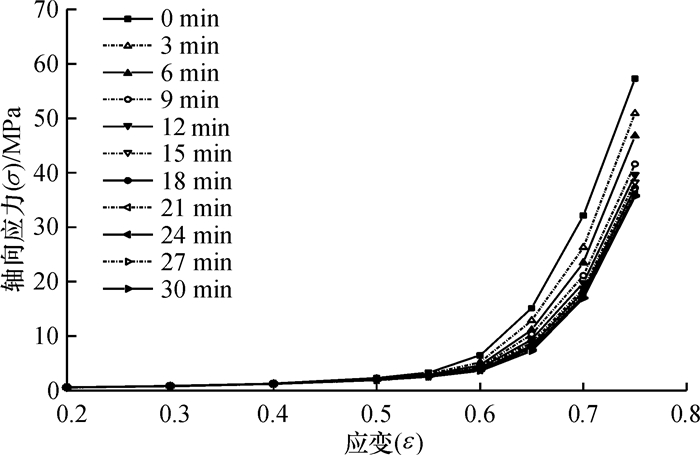
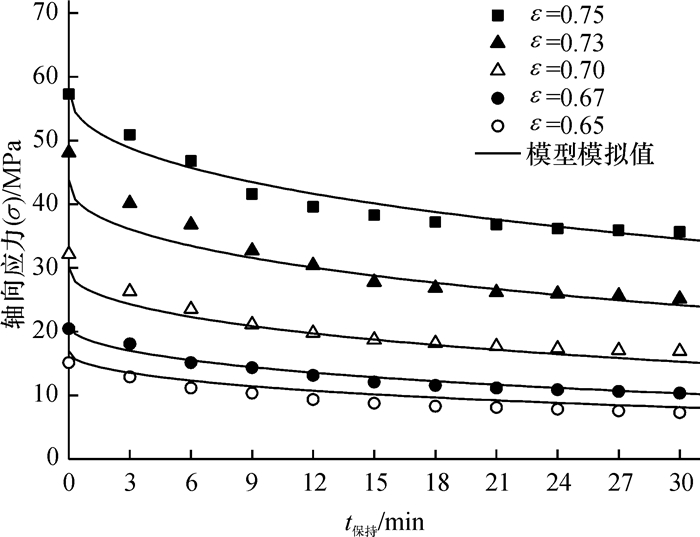
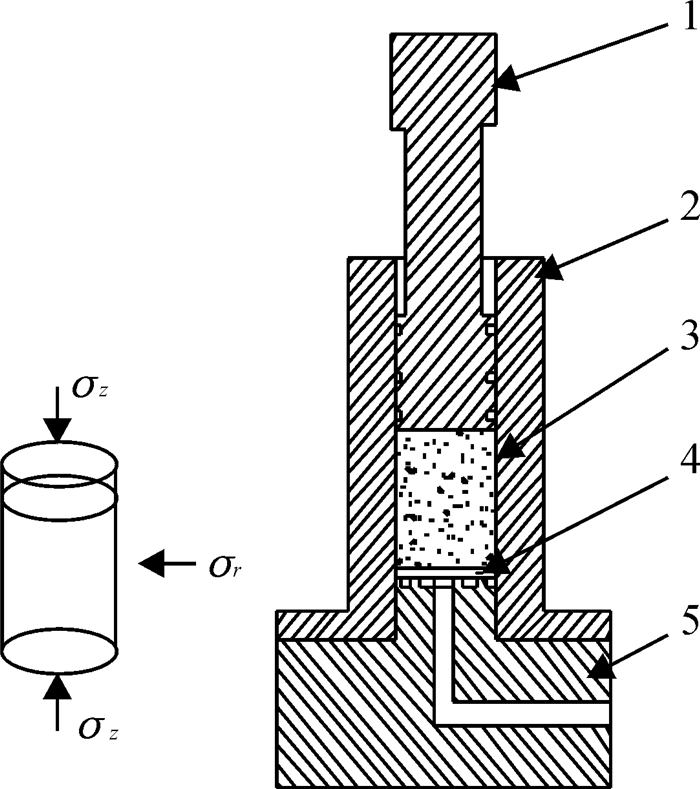
方法利用单轴压榨试验装置进行应力松弛试验,获取了松弛过程中应力与时间关系.基于试验结果和半理论半经验法,利用改进的广义Maxwell模型建立了蓖麻籽压榨非线性流变模型,使用非线性回归法分段对模型参数进行求解,并将模型计算值与试验结果对比.
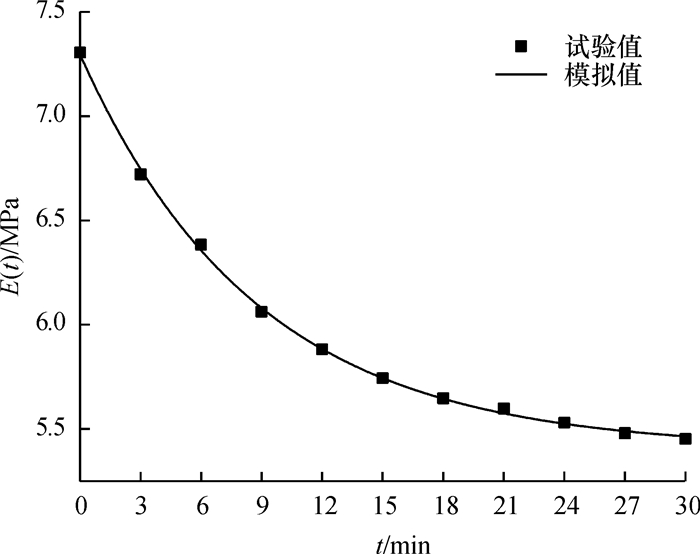
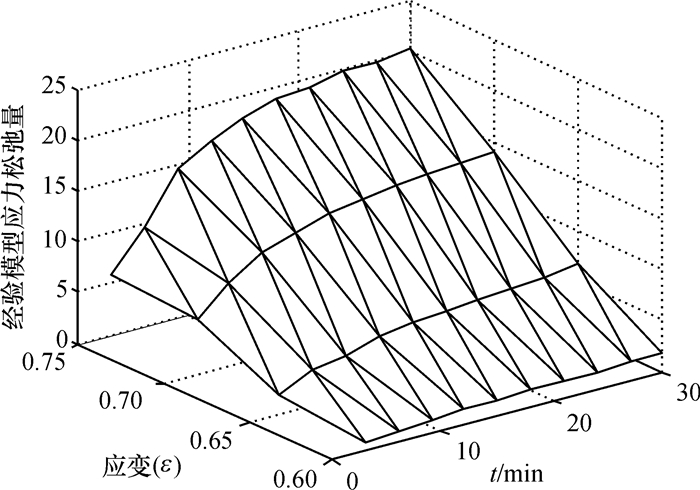
结果和结论综合对比结果可知:模型模拟值与应力松弛试验结果吻合较好,该模型能较好的模拟压榨应变大于0.65时蓖麻籽的非线性流变特性.
-
关键词:
- 蓖麻籽 /
- 应力松弛 /
- 改进的广义Maxwell模型 /
- 非线性流变模型
Abstract:ObjectiveTo reveal rheological characteristics in the process of castor seed press.
MethodThe stress-time relationship of stress-relaxation process was obtained by stress-relaxation tests with the uniaxial compression equipment. Based on the results and the semi-theoretical and semi-empirical method, a nonlinear rheological model was established using the improved generalized Maxwell model. The parameters of the model were achieved by nonlinear regression method.The calculated values of model were compared with the experimental results.
Result and conclusionThe results show that the model calculated values matching well with the experimental results, which means the model can simulate nonlinear rheological properties of castor seeds when the strain is over 0.65.
-
![]()
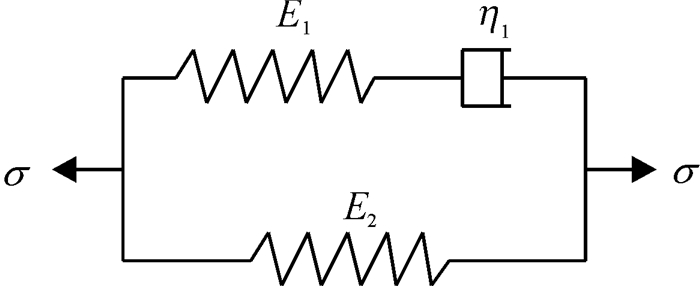
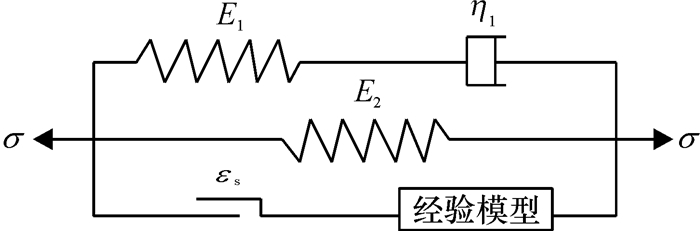
图 5 三元件广义Maxwell模型
Ei:弹性模量;η1:粘性系数.
Figure 5. Generalized Maxwell model of three elements
![]()
图 10 松弛试验与非线性理论模型对比
Figure 10. The comparison of stress-relaxation value and nonlinear model
-
[1] 郑晓. 油料压榨理论与试验研究[D]. 武汉: 武汉理工大学, 2005. [2] 张亚新. 植物油料压榨的塑性本构模型及有限元模拟分析[D]. 武汉: 武汉工业学院, 2009. [3] BARGALE P C. Mechanical oil expression from selected oilseeds under uniaxial compression[D]. Saskatchewan: University of Saskatchewan, 1997.
[4] 郑晓, 林国祥, 李智, 等.菜籽与菜籽仁的非线性粘弹塑性本构模型[J].农业机械学报, 2005, 36(11): 87-91. doi: 10.3969/j.issn.1000-1298.2005.11.022 [5] 汪训流, 陆新征, 叶列平.往复荷载下钢筋混凝土柱受力性能的数值模拟[J].工程力学, 2007, 24(12): 76-81. doi: 10.3969/j.issn.1000-4750.2007.12.014 [6] 罗迎社. 金属流变成形的理论, 实验与应用研究[D]. 长沙: 国防科学技术大学, 2000. [7] 于怀昌, 周敏, 刘汉东, 等.粉砂质泥岩三轴压缩应力松弛特性试验研究[J].岩石力学与工程学报, 2011, 30(4): 803-811. http://d.old.wanfangdata.com.cn/Periodical/yslxygcxb201104018 [8] 高洪梅, 刘汉龙, 刘金元. EPS颗粒轻质混合土蠕变特性试验研究[J].河海大学学报:自然科学版, 2010, 38(4): 402-406. http://d.old.wanfangdata.com.cn/Periodical/hhdxxb201004009 [9] 蔡美峰.岩石力学与工程[M].北京:科学出版社, 2002:200-203. [10] 冯上环, 赵有科.木材应力松弛特性及其影响因素概述[J].木材加工机械, 2010 (3): 37-40. doi: 10.3969/j.issn.1001-036X.2010.03.009 [11] 刘志平, 石林英.最小二乘法原理及其MATLAB实现[J].中国西部科技, 2008, 7(17): 33-34. doi: 10.3969/j.issn.1671-6396.2008.17.015 [12] 陈淑铭, 乔田田.一个求解非线性最小二乘问题的新方法[J].烟台大学学报:自然科学与工程版, 2004, 17(1): 14-22. http://d.old.wanfangdata.com.cn/Periodical/ytdxxb200401003 [13] 郑晓, 李智, 林国祥, 等.菜籽与脱皮菜籽冷榨的应力应变关系研究[J].农机化研究, 2004(6): 187-189. doi: 10.3969/j.issn.1003-188X.2004.06.070 [14] 葛荣德.川北公夫粉末压制方程应用中的几个问题及方程的修正[J].粉末冶金技术, 1993, 11(2): 90-94. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=QK000000471837
 下载:
下载: